- 18 -
Expression of Placenta Growth Factor mRNA Transcripts and Protein in Nasal Mucosa and Nasal Polyps
Sang-Hag Lee, M.D.1, Byung-Hoon Park, M.D.1, Hyun-Soo Jun, M.D.1, Woo-Jin Cho, M.D.1, Joon Choi, M.D.1, Hyun Woo Lim, M.D.1, Heung-Man Lee, M.D.1,
Joon-Hwan Oh, M.D.1 and Jae-Yong Lee, M.D.2
ABSTRACT
Placenta growth factor (PIGF) was originally described as a placenta produced homodimeric protein that shares substantial structural similarity with vascular endothelial growth factor (VEGF). It is becoming increasingly evident that PIGF may di- rectly or indirectly modulate several key vascular events in various tissues. These include angiogenesis or vasculogenesis, vas- cular maturation and stabilization, vascular permeability, and endothelial cell survival. Inflammatory reaction in the nasal mucosa increases mucosal vascular permeability, resulting in edematous nasal mucosa with polypoid change. In this respect, PIGF may play a role in the formation of nasal polyp. In the present study we evaluated the expression of PIGF mRNA and protein in human inferior turbinate mucosa and nasal polyp. The expression and localization of PlGF mRNA and protein were inves- tigated in the inferior turbinate mucosa and nasal polyps using reverse transcription-polymerase chain reaction (RT-PCR), Western blotting and immunohistochemistry. These results showed that the mRNA transcripts and protein for PlGF are expres- sed in human turbinate mucosa and nasal polyps. Semiquantitative RT-PCR revealed that PIGF mRNA in nasal polyps increased in its expression level than that in nasal turbinate mucosa. Likewise, immunoblot analysis demonstrated a higher expression of PIGF protein in nasal polyp tissues, compared with that of the nasal turbinate mucosa. However, immunohistochemical findings revealed that PlGF is localized in the endothelial lining of blood vessels in the inferior turbinate mucosa, whereas it is expressed in the epithelial cells of nasal polyps. These results indicate that PlGF mRNA and protein are expressed in normal turbinate mucosa and nasal polyp. Further, based on the fact showing that the expression site of PIGF is different in both tissues, the action mechanism of PIGF may be different in human nasal mucosa and nasal polyp. That is, the PIGF may play a role in the physiological function of normal nasal mucosa, possibly the maintenance of blood vessel and in the pathogenesis of nasal polyp formation.
KEY WORDS:Inferior turbinate mucosa・Nasal polyp・PlGF.
INTRODUCTION
Nasal polyposis is a common chronic disease of the nasal or paranasal sinus mucosa, with inflammatory cell infiltration and structural modifications of the epithe- lium and lamina propria. They are rounded projections
of edematous mucous membrane that arise from the paranasal sinus cavities, causing extremely disabling symptoms and require long-term treatment with cortico- steroids or even surgery. Their histopathology typically consists of an overlying intact respiratory epithelium with the underlying edematous or myxomatous stroma, showing various degrees of tissue remodeling in the epithelium, glands, connective tissue and vessels. Alt- hough numerous papers have recently dealt with the pathogenesis of this nasal disease, the mechanism lea- ding to the growth of nasal polyps still remain un- clear.1-4)
Numerous growth factors such as platelet-derived growth factors, keratinocyte growth factors, and insulin- like growth factor I have been postulated to be causative factors.1)5)6) Furthermore, several fibroangiogenic gro-
1Department of Otorhinolaryngology-Head & Neck Surgery, Col- lege of Medicine, Korea University, Seoul, 2Department of Otor- hinolaryngology-Head & Neck Surgery, College of Medicine, Soonchunhyang University, Bucheon Hospital, Bucheon, Korea Address correspondences and reprint requests to Sang-Hag Lee, M.D., Department of Otorhinolaryngology-Head & Neck Surgery College of Medicine, Korea University, 126-1, 5Ga, Anam-dong, Sungbuk-gu, Seoul 136-705, Korea
Tel:82-2-920-5486, Fax:82-2-925-5233 E-mail:[email protected]
Accepted for publication on August 19, 2004
wth factors such as basic fibroblast growth factor and vascular endothelial growth factor (VEGF) have been shown to be increased in nasal polyps, suggesting that these angiogenic factors, including edema and angio- genesis, could be involved in the pathogenesis of nasal polyps.7)8)
Placental growth factor (PIGF) is a member of the VEGF family of growth factors, encoding for a protein with an approximately 50% identity to VEGF in the platelet-derived growth factor-like domain. PIGF has been originally identified in the placenta, where it has been proposed to control trophoblast growth and diffe- rentiation. PIGF is also expressed during vascular deve- lopment and stimulates endothelial cell proliferation, migration and angiogenesis in vivo assays.9-10) In this respect, PIGF may play a role in the pathogenesis of nasal polyp formation. However, little attention has been paid to the expression of PIGF in nasal polyp tis- sue. In the present study, we examined the expression of PIGF mRNA transcripts and proteins in the human nasal mucosa and nasal polyp.
MATERIALS AND METHODS Sample preparations
Tissue from normal inferior turbinates was obtained from 20 subjects (13 men and 7 women, 20 to 35 years of age) undergoing surgery for augmentation rhinopla- sty. Nasal polyps were taken from 20 patients (17men and 3 women, 35 to 50 years of age) undergoing endo- scopic sinus surgery for chronic polypoid sinusitis.
These patients were selected on the basis of a history of chronic polypoid sinusitis of more than 3 years’ du- ration.
Tissues were cut into two portions ;one portion was dissected, frozen in liquid nitrogen, and stored at -70℃ for subsequent RNA/or protein isolation. For immunohistochemistry, the other portion was immer- sed overnight in a freshly prepared fixative containing 4% paraformaldehyde in phosphate-buffered saline (PBS), pH 7.4. All samples were then dehydrated in a graded series of ethanol to xylene and embedded in paraffin wax. The paraffin-embedded tissues were sec- tioned at 4 μm, collected on gelatin-coated glass slides and allowed to dry at room temperature.
Total RNA isolation and RT-PCR
The frozen tissues (50-100 mg) were homogenized in 1 ml of TRIzol® Reagent (GIBCOBRL, Grand Is- land, NY, USA), and total RNA was isolated as descri- bed previously.11) Total RNA from each sample was respectively reverse transcribed in 20 μl of a reaction mixture containing 2.5 U of M-MLV reverse transcrip- tase (GIBCO BRL, Grand Island, NY, USA) and 50 pm of random hexanucleotides at 42℃ for 60 mins.
Then 1 μl of each reaction product was amplified in 25 μl of PCR mixture containing 0.125 U of Taq DNA polymerase and 12.5 pmol of each primer. Primer sequences of PIGF and GAPDH gene used in this study are as follows:sense:5’- CTCCTAAAGATCCGTT- CTGG-3’, antisense:5’-GGTATAAATACACGAGC- CG-3’, sense:5’-ATCTTCCAGGAGCGAGATCC-3’, antisense : 5’-ACCACTGACACGTTGGCAGT-3’.
Amplified PCR products were resolved in 2% agarose gel, stained with ethidium bromide and photographed under ultraviolet light. Specificity of each PCR products was verified by its predicted size and DNA sequencing.
Immunohistochemical and western blot analysis
Sections were deparaffinized, rehydrated for 20 min in PBS and incubated in methanol containing 0.3% pe- roxide for 30 min. The sections were incubated in PBS containing normal sheep serum. Subsequently the sec- tions were incubated with the anti-PIGF antibody (1 μg/ml, Santa Cruz Biotechnology, USA) in PBS over- night at 4℃, and were incubated with a biotin-conjuga- ted anti-goat IgG antibody (Vector Lab) in PBS for 1 hour. The sections were then incubated with peroxidase- conjugated streptavidin for 1 hour. The color was deve- loped using DAB as a substrate for peroxidase.
For western blot analysis, tissues frozen in liquid nitrogen were crushed into pieces and vigorously vorte- xed in a buffer solution containing 50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1% Triton X-100, and 1 mM phenylmethylsulfonyl fluoride. 50 μg of the extracted protein was resuspended in a SDS sample buffer and boiled for 5 minutes. Equal amounts of total protein was separated on 10% SDS-polyacrylamide gels and trans- ferred to immobilon (Millipore, Bedford, Massachuse- tts). The blots were reacted with the anti-PIGF antibody (1 μg/ml, Santa Cruz Biotechnology, USA) in TBS
(20 mM Tris-HCl pH 8.0, 150 mM NaCl, 0.05% Triton X-100, and 5 % skim milk) for 2 hours at 4℃, and then reacted with a biotin-conjugated anti-goat IgG an- tibody (Vector Lab) in PBS for 1 hour. Antibody reac- tions were detected by using the ECL detection kit (Amersham), followed by detection of chemilumines- cence on x-ray film. After stripping, the blots were incu- bated firstly with an anti-β-actin antibody (Sigma, St.
Louis, Missouri) and secondarily with peroxidase-con- jugated antimouse IgG antibody (Vector), and then were reacted with chemiluminescence reagents before exposure. Intensity of detected bands was quantified by scion image (Scion Image Corp.). Relative intensities of PIGF signals were obtained by dividing intensities of PIGF signals by those of β-actin signals. Statistical significance of differences was tested by a one-way
PIGF
GAPDH
M T T T T P P P P
502 bp 248 bp 184 bp
0.5 0.4 0.3 0.2 0.1
T P
Ratio of relative PIGF protein expression
Fig. 1. Representative lanes of RT-PCR amplification with primers detecting PIGF, allowing the discrimination of sp- lice variants in the inferior turbinate mucosa and nasal polyp. T:inferior turbinate mucosa, P:nasal polyp, M:
100 bp DNA marker.
0.5 0.4 0.3 0.2 0.1
T P
Ratio of relative PIGF protein expression
19 kD PIGF
β-actin
T T T T P P P P
Fig. 2. Representative Western Blots for PIGF and β-actin (A). A strong band representing the molecular weight of PIGF can be seen in all inferior turbinate mucosa and nasal polyp studied. Gel electrophoresis was performed under reduced conditions of protein extracted from inferior turbinate mucosa and nasal polyp. KD shown in thousands. T:inferior turbinate mucosa, P:nasal polyp (lower panel). The density was calculated using an image analyzer and the bar represents the ratio of PIGF to β-actin in inferior turbinate mucosa and nasal polyp (mean±SD) (upper panel). p>0.05.
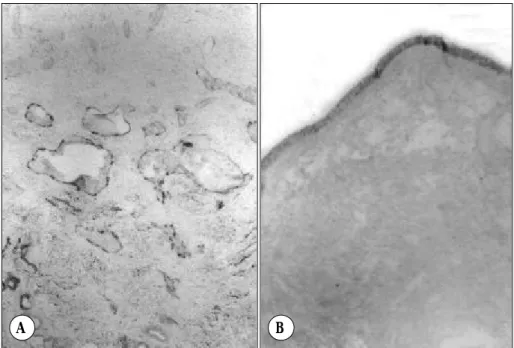
Fig. 3. Immunohistochemistry localizes the expression of PlGF protein in the endothelial cells of inferior turbinate mucosa (A) (orginal magnification:
×100) and epithelial cells of nasal po- lyp (B). (orginal magnification:×40).
A B
analysis of variance followed by Student’s t test. A p value of <0.05 was considered to be significant.
RESULTS
RNA was respectively isolated from inferior turbinate mucosa and nasal polyp tissues of 20 patients in which cDNA was amplified by the use of specific primer pairs, tested for PIGF. As a control for RNA integrity and RT-PCR procedure, the same RNA samples were also analyzed for GAPDH gene expression. This housekee- ping gene is commonly employed as an internal RNA control under different experimental conditions where transcription effects are being tested. All samples sho- wed an amplification product of the expected size of 502 bp at apparently similar densities. PIGF mRNA was detected in the human turbinate tissues and nasal polyp tissues examined. PIGF isoforms were visualized as the 184 bp (PIGF-1) and 248 bp (PIGF-2) amplification products (Fig. 1), and highly expressed in nasal polyps compared with normal nasal mucosa.
Incubation of immunoblots with the PIGF antiserum was able to detect PIGF protein in human turbinate and nasal polyp tissues. The expression levels of PIGF pro- tein in the nasal polyp is also upregulated, compared with inferior turbinate mucosa (Fig. 2). In nasal mucosa from healthy inferior trubinates, PIGF was only spora- dically detectable in the vascular endothelium (Fig. 3A).
PIGF-positive blood vessels were scattered over the whole area of the turbinate-submucosal layer, while nonimmune controls were negative. In contrast, PIGF immunoreactivity in nasal polyps was strongly positive in the epithelial layer, but not in the vascular endothe- lium (Fig. 3B).
DISCUSSION
Our present study examined the expression of PIGF mRNA transcript and protein in the human nasal mu- cosa and nasal polyp. This study has demonstrated for the first time that mRNAs coding for PIGF are present in human nasal mucosa and nasal polyp. The expression of PIGF protein was detected by immunohistochemistry and confirmed by Western blotting. In this study, results from immunohistochemistry revealed that the expres- sion of PIGF was localized mainly in the vascular endothelium of the inferior turbinate mucosa, but not in
nasal epithelium or inflammatory cells. On the contrary, in nasal polyp, the immunoreactivity for PIGF was lo- calized in the epithelial layer, but not in the vascular endothelium.
Angiogenesis, the process by which new vascular networks are formed from preexisting capillaries, is physiologically essential for embryogenesis, develop- ment and wound repair.12) It is also related to the prog- ression of various pathological conditions such as cancer growth and metastasis, diabetic retinopathy, rheumatoid arthritis, and collateral path formation in occlusive vas- cular disease.13) On the other hand, although very few studies have dealt with vascular remodeling in nasal polyp, the modifications of vessels observed in nasal polyps are suggestive of angiogenic processes, inclu- ding the endothelial folds or projection to the interstitium.
The endothelial projections complicatedly agglutinated to other parts of the endothelium and projections, indi- cating that capillary permeability of nasal polyps is higher than that of nasal mucosa.14) In animal models of nasal polyp formation, numerous vessels are present in the stroma of the growing polyps.4) Further, a plethora of candidate angiogenic growth factors have been iden- tified that may play a role in the formation of nasal polyp, including fibroblast growth factor, transforming growth factor and platelet-derived growth factor as well as insulin-like growth factor.2)5-7) More recently, atten- tion has focused on VEGF, which is a relatively specific mitogen for endothelial cells. The biological effects of VEGF include increased vascular dilatation and vascu- lar permeability.16) In normal nasal mucosa and nasal polyp, VEGF was mainly expressed in inflammatory and epithelial cells.8) Recent experiments using PIGF deficient mice revealed a novel biologic function of PIGF in the control of vascular leakage and edema for- mation.17) Therefore, the abundant expression of PIGF in nasal polyp demonstrated in the present study stron- gly indicates the involvement of pathological angiogene- sis and increased vascular permeability in the formation of nasal polyp.
PIGF is highly expressed in the placenta throughout all stages of human gestation. Immunohistochemsitry revealed the presence of PIGF in the vasculosyncytial membrane and in the media of large blood vessels of the placenta villi. Low levels of PIGF mRNA are also detected by means of Northern blot analysis in the hu- man thyroid, heart, brain, lung and skeletal muscle.10)
The analysis of full-thickness healing wounds in human skin revealed appreciable levels of PIGF mRNA and protein in the migrating keratinocytes starting from day 3 after injury. At day 13, when the wound had reepithe- lized, PIGF immunostaining was completely negative.18) In human and experimental choroidal neovasculariza- tion, PIGF mRNA was consistently expressed in human neovascular membranes, and its expression was pre- sent in the intact choroids and significantly upregulated during the course of experimental choroidal neovascula- rization.19) Thus, it has been suggested that PIGF is expressed at a basal rate in various tissues in healthy human adults and upregulates in the pathological an- giogenesis as occurs in cancer and ischemic and infl- ammatory diseases.10) In the present study, PIGF was expressed in normal human inferior turbinate mucosa where its expression was localized to the vascular endo- thelium. Further, the expression level of PIGF evaluated by western blotting was upregulated in nasal polyp more than nasal turbinate. As in other organs, the exact role of PIGF in the normal human nasal mucosa is unclear. A recent in vivo study suggested that PIGF may induce anti-apoptotic genes in endothelial cells, main- taining the integrity of endothelial cells.20) Therefore, PIGF may play a role in maintaining the integrity of endothelial cells of normal nasal mucosa. On the other hand, nasal mucosa, because of its direct contact with outside environment, t is difficult to obtain in its normal state because of extensive changes from external stimuli.
Therefore, even healthy humans without symptoms shows marked inflammatory cell infiltrates in normal inferior turbinate mucosa.21) which may induce the up- regulated PIGF expression. A possible explanation of our results could be supported by the results showing that cytokine secretion was found in nasal mucus of normal subjects22) and that PIGF is upregulated follo- wing cytokine treatment.18) The high expression level of PIGF in normal human nasal mucosa is also compa- tible with the well-known expression of VEGF in vari- ous tissues in healthy human adults that do not show angiogenesis.16)
Receptors for PIGF include the fms -like tyrosine kinase (flt-1) receptor and neurophilin-1 (NP-1) recep- tor. PIGF does not bind to the kinase insert domain- containing receptor (KDR), which is a receptor for VEGF.9)16) This receptor is expressed in a number of cells, including vascular smooth muscle cells, and mo-
nocytes.23) An immunohistochemical study revealed that flt-1 receptor was expressed in the vascular endothelium of the healthy turbinates and nasal polyp tissues.24)
CONCLUSION
In the present study, PIGF was expressed in the vas- cular endothelium of healthy nasal turbinate and in the epithelium of nasal polyp, respectively. Considering the function of PIGF, the coexpression of flt-1 in PIGF- producing vascular endothelial cells of healthy nasal mucosa raises the possibility that PIGF might affect the vascular endothelial cell biology directly through an autocrine mechanism. On the contrary it could be spe- culated that PIGF may act through a paracrine mecha- nism to trigger the growth of vascular endothelium in nasal polyp.
REFERENCES
1) Bateman ND, Fahy C, Woolford TJ. Nasal polyps: Still more ques- tions than answers. J laryngol Otol 2003;117:1-9.
2) Coste A, Wang QP, Roudot-Thoraval F, Chapelin C, Bedbeder P, Poron F, et al. Epithelial proliferation in nasal polyps could be up- regulated by platelet-derived growth factor. Laryngoscope 1996;106:
578-83.
3) Larsen PL, Tos M, Kujipers W, van der Beek JMH. The early sta- ges of polyp formation. Laryngoscope 1992;102:670-7.
4) Norlander T, Westrin KM, Fukami M, Stierna P, Calsoo B. Experi- mentally induced polyps in the sinus mucosa: a structural analysis of the initial stages. Laryngoscope 1996;106:196-203.
5) Ishibashi T, Tanaka T, Nibu K, Ishimoto S, Kaga K. Keratinocyte growth factor and its receptor messenger RNA expression in nasal mucosa and nasal polyps. Ann Otol Rhinol Laryngol 1998;107:
885-90.
6) Petruson B, Hansson HA, Petruson K. Insulinlike growth factor I immunoreactivity in nasal polyps. Arch Otolaryngol Head Neck Surg 1988;114:1272-5.
7) Powers MR, LaGesse PC, Liebler JM, Wall MA, Rosenbaum JT.
Expression of basic fibroblast growth factor in nasal polyps. Ann Otol Rhinol Laryngol 1998;107:891-7.
8) Coste A, Brugel L, Maitre B, Boussat S, Papon JF, wingerstmann L, et al. Inflammatory cells as well as epithelial cells in nasal polyps express vascular endothelial growth factor. Eur Respir J 2000;15:
367-72.
9) Ziche M, Maglione D, Ribatti D, Morbidelli L, Lago CT, Battisti M, et al. Placenta growth factor-1 is chemotactic, mitogenic, and angio- genic. Lab Invest 1997;76:517-31.
10) Persico MG, Vinventi V, DiPalma T. Structure, expression and recep- tor-binding properties of placenta growth factor (PIGF). Curr Top Microbiol Immunol 1999;237:31-40.
11) Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidium thiocyanate-phenol-chloroform extraction. Anal Bio- chem 1987;162:156-9.
12) Risau W. Mechanisms of angiogenesis. Nature 1997;386:671-4.
13) Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat Med 1995;1:27-31.
14) Watanabe K, Komatsuzaki A. Ultrastructural findings of capillaries in nasal polyps. Rhinology 1992;30:49-56.
15) Wang QP, Escudier E, Roudot-Thoraval F, Abdi-Al Samad I, Peyne- gre R, Coste A. Myofibroblast accumulation induced by transfor- ming growth factor-beta is involved in the pathogenesis of nasal polyps. Laryngoscope 1997;107:926-31.
16) Witmer AN, Vrensen GFJM, Van Noorden CJF, Schlingemann RO.
Vascular endothelial growth factors and angiogenesis in eye disease.
Prog Retin Eye Res 2003;22:1-29.
17) Luttun A, Brusselmans K, Fukao H, Tjwa M, Ueshima S, Herbert JM, et al. Loss of placental growth factor protects mice against vas- cular permeability in pathological conditions. Biochem Biophys Res Commun 2002;295:428-34.
18) Failla CM, Odorisio T, Cianfarani F, Schitroma C, Puddu P, Zambu- runo G. Placenta growth factor is induced in human keratinocytes during wound healing. J Invest Dermatol 2000;115:388-95.
19) Rakic JM, Lambert V, Devy L, Luttun A, Carmeliet P, Claes C, et al.
Placental growth factor, a member of the VEGF family, contributes
to the development of choroidal neovascularization. Invest Ophth- almol Vis Sci 2003;44:3186-93.
20) Adini A, Kornaga T, Firoozbakht F, Benjamin LE. Placental growth factor is a survival factor for tumor endothelial cells and macropha- ges. Cancer Res 2002;62:2749-52.
21) Schrodter S, Biermann E, Halata Z. Histological evaluation of age- related changes in human respiratory mucosa of the middle turbinate.
Anat Embryol 2003;207:19-27.
22) Scavuzzo MC, Rocchi V, Fattori B, Ambrogi F, Carpi A, Ruffoli R, et al. Cytokine secretion in nasal mucus of normal subjects and pa- tients with allergic rhinitis. Biomed Pharmacother 2003;57:366-71.
23) Ahmed A, Ahmed DS, Khaliq A. Regulation of placental vascular endothelial growth factor (VEGF) and placenta growth factor (PIGF) and soluble Flt-1 by oxygen- a review. Placenta 2000;21:S16-24.
24) Wittekindt C, Hess A, Bloch W, Sultanie S, Michel O. Immunohis- tochemical expression of VEGF and VEGF receptors in nasal polyps as compared to normal turbinate mucosa. Eur Arch Otorhinolary- gol 2002;259:294-8.