- 120 -
Regional Differences in Proliferative Activity of Nasal Epithelium in Rat
Sang Hag Lee, M.D., Jae Yong Lee, M.D. and Heung Man Lee, M.D.
ABSTRACT
We investigated the active proliferation sites of epithelial cells in normal nasal mucosa by immunohistochemical staining of proliferating cell nuclear antigen (PCNA), the marker of S phase of cell cycle and active cell proliferation. The whole nasal mucosa of the ten normal Sprague-Dawley rats were processed for PCNA immunolabeling. In respiratory portion, distinctly positive reaction was seen mainly in the anterior aspect, that is, the nuclei of squamous and non-ciliated cuboidal/transitional epithelium. These types of epithelial cells are transformed to pseudostratified ciliated epithelium in the posterior direction where positive reaction became scanty. In olfactory epithelium, the nuclei immunoreactive for PCNA were distinct in some area, but absent in other adjacent areas, lacking of region-specific immunolabeling that was observed in respiratory mucosa. These results suggest that anterior portion of nasal cavity is the main proliferation zone of normal nasal respiratory epithelium as well as the main site of protective function. In contrast, the neurogenesis of the olfactory nerve cells is not site-specific, indicating that any region covered by olfactory mucosa may be the main proliferation zone.
KEY WORDS:Proliferating cell nuclear antigen·Respiratory epithelium·Olfactory epithelium.
INTRODUCTION
Nasal mucosa is the initial site of contact with inhaled irri- tants such as altered airflow, atmospheric pollutants, allergens and virus infections, and functions to protect itself and lower respiratory tract from these environmental toxicants. It is ge- nerally believed that the nasal epithelium plays an important role in these protective functions by absorbing water-soluble gases, trapping and clearing inhaled particles.
A number of histopathological studies revealed that in lab- oratory rodents, inhalation of harmful toxicants results in sig- nificant morphologic alterations to surface epithelia lining the nasal respiratory tract, increasing the rate of surface epithelial cell proliferation.1-3) This proliferative response is believed necessary to replace the damaged epithelium and to protect the nasal mucosa from further insult by repairing and adapting the epithelial barrier. However, little attention has been paid to the nasal epithelium in terms of its proliferative response in the steady state without inhalation of toxicants.
The olfactory epithelium presents unique cytological char-
acteristics in the nervous system of vertebrates in that there is continuous turnover and replacement of the sensory neurons in the olfactory system. Mature olfactory neurons die and are replaced from undifferentiated neuroblasts over the entire life span of the individuals. Furthermore, a number of experim- ental techniques have been applied to induce degeneration of the olfactory neurons in order to study subsequent neuroge- nesis and replacement of this cell population.4-6) These studies clearly demonstrated replacement of receptor cells, indicating that the olfactory epithelium is significantly recovered follo- wing injury. However, the pattern, density or location of olfa- ctory cell proliferation in normal nasal mucosa has not been determined precisely.
The present study was undertaken to determine the distr- ibution of epithelial cells undergoing DNA replication in the various regions of the rat nasal cavity. DNA replication was detected with immunohistochemical method using antibody against proliferating cell nuclear antigen (PCNA). PCNA is auxiliary protein for DNA polymerase delta, which is expre- ssed in late G1, during S and early G2 phases of the prolife- rating cell.7)8)
MATERIALS AND METHODS
Tissue preparationsTen healthy adult Sprague-Dawley rats (200-300 g) were used in this experiment. They were anesthetized with intrap- Department of Otorhinolaryngology-Head and Neck Surgery,
College of Medicine, Korea University, Seoul, Korea
Address correspondence and reprint requests to Sang Hag Lee, M.D., Department of Otorhinolaryngology-Head and Neck Surgery, College of Medicine, Korea University, 126-1, 5 Ka, Anam-Dong, Sungbuk-Ku, Seoul 136-705, Korea
Tel:82-2-920-5486, 5761, Fax:82-2-925-5233 Accepted for publication on September 14, 1997
eritoneal injection of sodium pentobarbital (30 mg/Kg) and perfused through the aorta with normal saline, followed by 150 ml of 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). After perfusion fixation, the nasal mucosa was tot- ally separated from nasal septum and lateral wall of nasal ca- vity, and immersed in 20% sucrose solution at 4℃ for 6 hours.
They were embedded in OCT compounds (Tissue Tek® Miles Scientific, Naper-ville, USA) and quickly frozen in liquid ni- trogen. Whole tissue samples were serially sectioned from up- per to lower portion in nasal septum and from frontal to rostral portion in lateral wall of nasal cavity including turbinates.
After being sectioned at 15 μm in a cryostat (Leica CM 1800, Germany) and selected one every 5-section intervals, they were mounted on poly-L-lysine-coated glass slides and processed for PCNA immunolabeling.
Immunohistochemistry for PCNA
Cryostat sections were air-dried and immersed in methanol mixed with 0.6% H2O2 for 30 minutes for blocking endogen- ous peroxidase. After washing in phosphate-buffered saline (PBS, pH 7.4) for 5 min, the sections were boiled in citrate- buffer (pH 6.0, 3 microwave exposure cycles at 650 W, 5 mi- nutes) and incubated in 0.05% saponin (Sigma Chemical Co- mpany, USA) for 30 minutes followed by rinsing in PBS again.
They were incubated in normal horse serum for 30 minutes, and after washing in PBS, incubated with a 1:100 dilution of anti-PCNA monoclonal mouse antibody (DAKO Co., USA) in moist chamber at 4 ℃ overnight. The sections were was- hed in PBS and incubated with biotinylated anti-mouse IgG secondary antibody for 1 hour, followed by streptavidin-con- jugated peroxidase for 1 hour (Vectastain® ABC Kit PK 4002, Vector Lab., Burlingame, CA, USA). The immunoreactivity was visualized by diaminobenzidine (D-AB)-H2O2 solution (5 mg DAB/10 ml PBS+5 μl 30% H2O2). The immunost- ained sections were dehydrated in a graded ethanol series and mounted with Permount® (Fisher Scientific, New Jersey, USA).
They were observed under light microscopy and the epithelial cell nuclei with intensely brown red, granular or diffuse stai- ning pattern corresponding to S phase PCNA were regarded as PCNA-positive.
RESULTS
Nasal septumIn respiratory portion, distinctly positive reaction was obs- erved in the basal and suprabasal layers of squamous and non- ciliated cuboidal/transitional epithelium, which were located in anterior portion of nasal cavity. In these epithelia immuno- labeled cells were observed in the basal cell layers, and existed
continuously along the basement membrane of epithelium (Fig. 1). These sqaumous and non-ciliated cuboidal/transiti- onal type of epithelial cells were transformed to pseudostratified ciliated respiratory epithelium in the posterior direction where positive reaction became scanty and was limited to the basal cells. In this anterior portion of ciliated respiratory epithelium, PCNA-positive cells were scattered with some distance along the basement membrane, and multiple layers of positive cells were rarely seen (Fig. 2). PCNA-positive reaction became more sparse in the posterior portion of ciliated respiratory ep- ithelium, and the positive nuclei existed with wider distance.
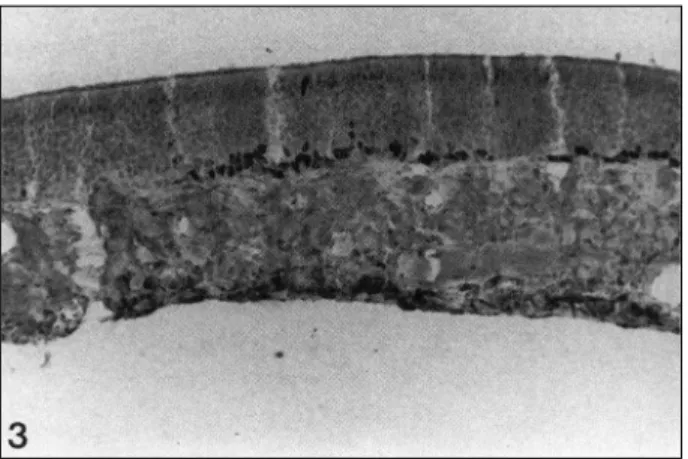
This distribution pattern of cell nuclei positive for PCNA were observed in all sections from upper to lower portion of nasal septum. In olfactory epithelium, the nuclei immunoreactive for PCNA were observed in the basal cell layers, but not in the intermediate or upper olfactory neuronal cells. The labeling was distinct in some area, but absent in adjacent other portions (Fig. 3). However, compared with the respiratory mucosa, the olfactory epithelium did not show the region-specific immu-
Fig. 1. The anterior portion of nasal septal mucosa (squamous and non-ciliated cuboidal/transitional epithelium). These epit- helia show distinctly PCNA-positive reaction in the basal layers (×100).
Fig. 2. The posterior portion of nasal septal mucosa (anterior po- rtion of pseudostratified ciliated respiratory epithelium). In this portion, PCNA-positive reaction is not prominent and sparse even if it exists (×100).
nolabeling, that is, anterior-posterior gradient of PCNA-positive nuclei was not observed in olfactory epithelium of nasal septal mucosa. Schematic illustration of the distribution pattern of PCNA-positive cells in the nasal septal mucosa is shown in Fig. 4.
Lateral wall of nasal cavity including turbinates
In nasal mucosa of turbinates and lateral wall of nasal cavity, the distribution pattern of epithelial cell nuclei positive for PCNA was similar compared with septal mucosa. Positive reaction for PCNA was observed in both respiratory and olf- actory mucosa. In respiratory portion, densely clustered positive nuclei were noted mainly in the squamous and non-ciliated
cuboidal/transitional epithelium which were located anteriorly in nasal cavity (Figs. 5 and 6). These positive cells existed in the basal cell layers. In the anterior end of nasoturbinate and maxilloturbinate, there was some portion covered with squ- amous epithelium in which abundant positive reaction was observed. These dense PCNA-positive cells in the squamous and non-ciliated cuboidal/transitional epithelium became sca- nty and scattered as directed posteriorly where the epithelium was substituted for pseudostratified ciliated respiratory epith- elium. In this anterior portion of ciliated respiratory epithelium, PCNA immunolabeling was limited to the basal cells and in- continuously distributed (Fig. 7). The posterior portion of pseudostratified ciliated respiratory epithelium, PCNA-positive cells became more sparse, and existed with wider distance. As observed in the septal mucosa, respiratory portion of turbina- tes and lateral wall of nasal cavity showed anterior-posterior gradient of PCNA positivity. In olfactory epithelium, as in nasal septal mucosa, the nuclei immunoreactive for PCNA were observed in the basal cell layers, and the labeling was
Fig. 4. Schematic illustration of distribution pattern of PCNA- positive epithelial cells in normal nasal septal mucosa. Black dots represent PCNA-positive epithelial cells. In respiratory por- tion, positive reaction is mainly seen in the squamous and non- ciliated cuboidal/transitional epithelium which are located in the anterior portion of nasal cavity. These types of epithelial cells are transformed to pseudostratified ciliated epithelium in the posterior direction where positive reaction become scanty.
In the olfactory epithelium, the nuclei immunoreactive for PCNA are distinct in some area, but absent in other adjacent portions:
I-squamous epithelium, II-non-ciliated cuboidal/transitional ep- ithelium, III-pseudostratified ciliated respiratory epithelium, IV- olfactory epithelium.
Fig. 3. The olfactory portion of nasal septal mucosa. This olfac- tory epithelium shows PCNA-positive cells in basal cell layers and this positive reaction is distinct in some area while adjacent other portions are absent of PCNA immunolabeling (×100).
Fig. 5. The coronal section of the turbinates at the level of squ- amous and non-ciliated cuboidal/transitional epithelium. These epithelia shows distinctly PCNA-positive reaction in the basal layers (×100).
Fig. 6. The coronal section of the lateral wall of transitional epi- thelium. PCNA-positive reaction is prominently seen in the basal layers (×100).
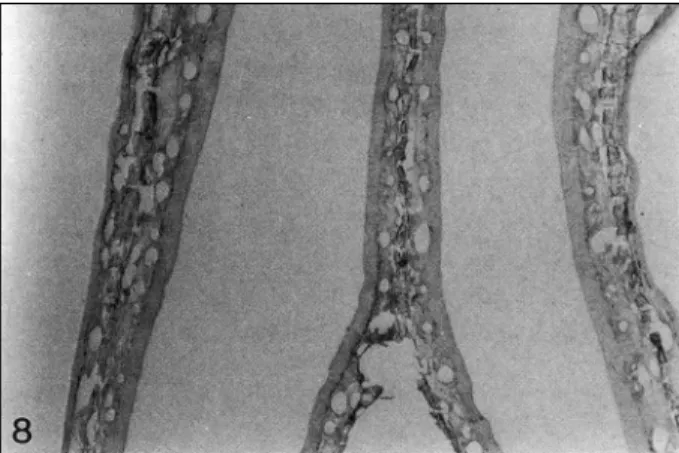
distinct and clustered in some portion, while adjacent other portions were absent of positive reaction (Fig. 8). The olfac- tory epithelium did not show the region-specific distribution of PCNA-positive nuclei that was observed in the respiratory mucosa. Schematic illustration of the distribution pattern of PCNA-positive cells in the turbinates and lateral wall of the nasal cavity is shown in Fig. 9.
DISCUSSION
This is the first study in which immunohistochemistry with antibodies to PCNA has been used to reveal proliferative act- ivity of epithelial cells lining the nasal mucosa of normal rat.
This immunohistochemical method is relatively simple com- pared to the use of tritiated thymidine or bromdeoxyuridine and demonstrates the usefulness of this technique in localizing the epithelium with proliferative ability in nasal tissue.2)9) This study also shows that cell turnover occurs without overt signs of cell damage or death under normal conditions. Cell prolif- eration in the non-olfactory epithelium was most prominent in the anterior region of the nose, showing region-specific dist- ribution while in the olfactory region, the epithelial cells with actively proliferative ability were more widely distributed.
The non-olfactory epithelium of the rat nasal cavity is co- mposed of three morphologically distinct epithelial populati- ons. These populations have specific locations within the nasal cavities and are composed of different epithelial cell types.
These regions include:stratified squamous epithelium in the nasal vestibule;ciliated pseudostratified respiratory epithel- ium in the main cavity of the nasal airway;a narrow region of non-ciliated, cuboidal, transitional epithelium lying between the squamous and respiratory epithelium in the anterior aspect of the main nasal cavity. These specifically defined epithelial regions are similarly observed in all commonly used labora- tory animals.1-3)10) It seems that the distribution pattern of these epithelial population is a normal findings and not a patholo- gical change in nasal respiratory epithelium.
Air stream alone is thought to be the physiological irritant which includes various kinds of toxicants;virus, bacteria, ch- emical pollutants, physical factors and other substances. In the present study, increased numbers of PCNA stained nuclei was predominantly observed in the squamous and transitional epithelium. These findings indicate that exposure of normal nasal surface epithelium to inhaled air is accompanied by ep- ithelial cell proliferation in even normal nasal breathing. Pa- rticularly, squamous and transitional epithelia of the nasal cavity are assumed to be more intensely stimulated by the continuous physiological air stream and may be particularly vulnerable to inhaled air because of their proximal location in the nasal airway. These assumptions are supported by previous
Fig. 7. The coronal section of lateral wall of nasal cavity at the level of anterior portion of pseudostratified ciliated respiratory epithelium. This portion shows scanty and scattered PCNA-po- sitive nuclei limited to basal cell layers (×100).
Fig. 8. The coronal section of nasal mucosa of the turbinates and lateral wall of nasal cavity at the level of olfactory epithelium.
This epithelium shows PCNA-positive cells in basal cell layers and this positive reaction is distinct in some area while adjacent other portions are absent of PCNA immunolabeling (×40).
Fig. 9. Schematic illustration of distribution pattern of PCNA-po- sitive epithelial cells in normal nasal mucosa of the lateral wall of nasal cavity including turbinates. Black dots represent PCNA- positive epithelial cells. In both respiratory and olfactory mucosa, distribution pattern of the positive reaction for PCNA is identical to that of the septal mucosa:I-squamous epithelium, II-non- ciliated cuboidal/transitional epithelium, III-pseudostratified ci- liated respiratory epithelium, IV-olfactory epithelium.
studies showing that surgical closure of the nostril induces the regeneration of atrophic or metaplastic respiratory epithelium in human and dog.11)12)
The previous studies reported the effects of inhaled toxic- ants on surface epithelia lining the nasal cavity. These studies suggested that the distribution of toxicant-induced lesions in the nasal cavity is thought to arise from a combination of the effects of regional uptake patterns, local clearance processes, and differences in the metabolic capacity or inherent sensiti- vity of various cell types to a given toxicants.13)14) There may also be differences in the susceptibility of transitional and sq- uamous epithelium in anterior nasal cavity to inhaled toxicants.
Both epithelia lining the anterior nasal cavity normally contain few mucous cells, while numerous mucous cells are present in the ciliated respiratory epithelium. Therefore, increased nu- mbers of mucous cells may help to protect against toxicants- induced injury through increased local secretion of mucus.
Laboratory rodents exposed to highly water-soluble, gaseous irritants most often show lesions on the distal tips and lateral margins of the naso-and maxillo-turbinates and on the adjacent lateral wall in the anterior aspect of the nasal cavity, which are covered by transitional epithelium. Relatively high amounts of cytochrome P-450-dependent mono-oxygenase have been re- ported in the nasal mucosa of the rat, rabbit, and dog, covering the anterior nasal cavity. The anterior aspect of the dorsal co- ncha of the dog nasal cavity, a region covered by transitional epithelium, has recently been shown to have higher metaboli- zing activity for certain xenobiotic substrates than other regi- ons of canine nasal mucosa. It has also been demonstrated that similar non-ciliated cuboidal cells lining the maxilloturbinates in the anterior nasal cavity of the rabbit contain certain cyt- ochrome P-450 isozymes not evident in adjacent respiratory epithelium.15) It seems likely that these transitional epithelial cells in rabbit, dogs, rats, and other rodents may play signi- ficant roles in the metabolism of certain inhaled xenobiotics, covering the main proliferation zone of the normal nasal mu- cosa as well as the main site of protective function.
Olfactory neurons die and are continuously replaced in fully mature mammals because of the unique ability of basal olf- actory epithelial cells to generate new populations of sensory neurons throughout the life span of many animals. Several po- ints of interest in previously performed regeneration studies are the origin of new cells, the effect of absence of olfactory bulb, cell turnover rate, etc.4-6)16)17) Evidence from in vivo ex- periments suggested that the horizontal basal cells undergo asymmetric divisions to produce one horizontal basal cell and one neuronal precursor cell. The neuronal precursor divides symmetrically to generate two undifferentiated neurons. As the immature neurons differentiate, they migrate away from the basal layer of the epithelium.16)17) In addition to these preex-
isting results, this study has systemically analyzed the distri- bution pattern of actively proliferating zone in the olfactory mucosa and produced two major findings. First, basal cells only demonstrated PCNA immunopositivity in the olfactory epi- thelium of rat, indicating the origin of new cells. Second, the distribution of basal cells with proliferation activity did not show the region-specific pattern in the whole nasal cavity. That is, the neurogenesis of olfactory nerve cells is most likely to occur in some area, but not in adjacent or other areas. These findings were constantly noted in septal and ethmoturbinates mucosa which was covered by the olfactory mucosa. Taken to- gether, it seems that the neurogenesis of olfactory mucosa, at least, is independent of air stream inhaled into the nasal cavity.
In summary, the results of present study suggest that mitotic activity of epithelial cells may be related to the stimuli of en- vironmental factor even in normal physiologic status in the respiratory mucosa, but not in the olfactory mucosa, and that anterior portion of nasal cavity is the main proliferation zone of the normal nasal mucosa as well as the main site of prote- ctive function.
REFERENCES
1) Johnson NF, Hotchikiss JA, Harkema JR, Henderson RF. Prolife- rative responses of rat nasal epithelia to ozone. Toxicol Appl Pha- rmacol 1990;103:143-55.
2) Hotchikiss JA, Evans WA, Chen BT, Finch GL, Harkema JR. Re- gional differences in the effects of mainstream cigarette smoke on stored mucosubstances and DNA synthesis in F344 rat nasal res- piratory epithelium. Toxicol Appl Pharmacol 1995;131:316-24.
3) Monticello TM, Miller FJ, Morgan KT. Regional increases in rat nasal epithelial cell proliferation following acute and subchronic inhalation of formaldehyde. Toxicol Appl Pharmacol 1991;111:
409-21.
4) Inamitsu M, Nakashima T, Uemura T. Immunopathology of olfa- ctory mucosa following injury to the olfactory bulb. J Laryngol Otol 1990;104:959-64.
5) Costanzo RM. Comparison of neurogenesis and cell replacement in the hamster olfactory system with and without a target (olfac- tory bulb). Brain Res 1984;307:295-301.
6) Monti-Graziadei GA, Graziadei PPC. Neurogenesis and neuron regeneration in the olfactory system of mammals. II. Degeneration and reconstitution of the olfactory sensory neurons after axotomy.
J Neurocytol 1979;8:197-213.
7) Bielicki D, Markiewski M, Marlicz K, et al. PCNA/cyclin defined proliferative activity of epithelial cells in normal gastric mucosa and in chronic gastritis. Patol Pol 1993;44:75-8.
8) Diebold J, Lai MD, Lhrs U. Analysis of proliferative activity in colorectal mucosa by immunohistochemical detection of prolifer- ating cell nuclear antigen (PCNA): Methodological aspects and application to routine diagnostic material. Virchows Arch B Cell Pathol Incl Mol Pathol 1992;62:283-9.
9) Breuer R, Zajicek G, Christensen TG, Lucey EC, Snider GL. Cell kinetics of normal adult hamster bronchial epithelium in the ste- ady state. Am J Respir Cell Mol Biol 1990;2:51-8.
10) Harkema JR, Plopper CG, Hyde DM, Wilson DW, George JAST,
Wong VJ. Nonolfactory surface epithelium of the nasal cavity of the bonnet monkey: A morphologic and morphometric study of the transitional and respiratory epithelium. Am J Anat 1987;180:
266-79.
11) Batistic VCB, Sankovic F, Rijeka Z. Cytological and histological alterations in the nasal mucous membrane during experimental ob- struction of nasal openings. Rhinology 1987;25:5-12.
12) Elwany S. Ultrastructural observations on primary atrophic rhinitis:
Effect of partial closure of the nostril. ORL 1988;50:389-96.
13) Morgan KT, Monticello TM. Airflow, gas deposition and lesion distribution in the nasal passages. Environ Health Perspect 1983;
47:345-55.
14) Hotchkiss JA, Herrera LK, Harkema JR, Kimbell JS, Morgan KT,
Hatch GE. Regional differences in ozone-induced nasal epithelial proliferation in F344 rats: comparison with computational mass flux predictions of ozone dosimetry. Inhal Toxicol 1994;6:390-2.
15) Harkema JR. Comparative pathology of the nasal mucosa in lab- oratory animals exposed to inhaled irritants. Environ Health Pers- pect 1990;35:231-38.
16) Graziadei PPC, Monti-Graziadei GA. Neurogenesis and neuronal regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfac- tory sensory neurons. J Neurocytol 1979;8:1-18.
17) McKay-Sims A, Kittel P. Cell dynamics in the adult mouse olfact- ory epithelium: A quantitative autoradiographic study. J Neurosci 1991;11:979-84.