- 112 -
Cooperative effect of Alternaria and rhinovirus on the activation of nasal polyp epithelial cells
Seung-Heon Shin, MD
1, Mi-Kyung Ye, MD
1, Byeong-Gyu Jeon, MD
1and Yong Ju Jang, MD
21
Department of Otolaryngology, Catholic University of Daegu School of Medicine, Daegu, South Korea
2
Department of Otolaryngology, Asan Medical Center, University of Ulsan College of Medicine, Seoul, South Korea
ABSTRACT
Background and Objectives : The nasal epithelium is the first barrier encountered by airborne allergens and is an active participant in airway inflammation. The aim of this study was to determine the activation mechanism of nasal epithelial cells with Alternaria and the effect of rhinovirus on the Alternaria induced activation of nasal epithelial cells. Materials and methods : Cultured epithelial cells were stimulated by Alternaria with or without rhinovirus-16 (RV-16) infection. Release of interleukin (IL)-6, IL-8, and granulocyte macrophage colony-stim- ulating factor (GM-CSF) into culture supernatants were measured to determine the activation of epithelial cells.
Nuclear factor-κB (NF-κB) and activator protein-1 (AP-1) of the epithelial cells were analyzed using western blot analysis. Intracellular NF-kB and AP-1 activity were evaluated by enzyme-linked immunosorbent assay. To deter- mine the epithelial cell activation mechanism, cytokine production was inhibited with NF-kB, AP-1, and mitogen activated protein kinase (MAPK) inhibitors. Results : Exposure of epithelial cells to Alternaria enhanced the production of cytokines. Intracellular NF-kB expression and activity were significantly increased by Alternaria, but not by RV-16. AP-1 expression and activity were not influenced by Alternaria. Increased IL-6 production was significantly inhibited by transcription factor inhibitors. However, IL-8 and GM-CSF production were not inhibited by these transcription factor inhibitors. Conclusions : Our in-vitro results demonstrate that Alternaria activates nasal polyp epithelial cells via NF-kB pathway and that NF-kB, AP-1, and MAPK are involved in the production of IL-6.
KEY WORDS : Epithelial cell · Alternaria · Rhinovirus · Nuclear factor-κB · Activator protein-1.
INTRODUCTION
Fungi are environmentally ubiquitous and coexist with- out affecting the normal cellular immunity of the host. Air- borne fungal spores enter the upper and lower respiratory tract by inhalation, but are rarely pathogenic in healthy individuals. Airborne fungi and fungal elements induce immune and inflammatory responses. Proteases of fungal extracts are potent inducers of epithelial cell desquama- tion and production of proinflammatory cytokines.
1) 2)Al- ternaria, Aspergillus, Penicillium, and Cladosporium are commonly found in the upper airway secretion of patients with chronic rhinosinusitis (CRS) and normal healthy
controls.
3) 4)Alternaria extracts have a high protease de- pendent activation and are highly potent in inducing cy- tokine production. Allergic sensitivity to and the airborne presence of Alternaria are associated with development and persistence of CRS and asthma.
5) 6)Rhinovirus (RV) is the major cause of the common cold and the most common acute infection-related illnesses in humans, and are associated with acute exacerbations of bronchial asthma.
7)RV infect nasal epithelial cells and cause a local inflammatory response that is associated with the production of kinins and cytokines.
8) 9)Intercel- lular adhesion molecule (ICAM)-1 is the airway epithelial receptor for major subgroup RVs (RV-14, RV-16, and RV- 39).
10)Cytokine production to RV-16 infection in airway epithelial cells is inhibited by antibodies to ICAM-1, im- plicating the integrin-mediated pathway. NF-κB activa- tion is required for a number of RV responses including airway epithelial cell expression of IL-6, IL-8, ICAM-1, and vascular cell adhesion molecule-1.
10)Activated respiratory epithelial cells are a major source Address correspondence and reprint requests to Seung-Heon Shin, MD,
PhD, Department of Otorhinolaryngology, Daegu Catholic University Medical Center, 3056-6 Daemyung 4 Dong, Nam-Gu, Daegu 705-718, South Korea
Tel: +82-53-650-4530 · Fax: +82-53-650-4533 E-mail: hsseung @cu.ac.kr
Received for publication on July 3, 2012
Accepted for publicatoin on August 14, 2012
of inflammatory chemical mediators, which modulate the local inflammatory reaction and the recruitment of inflam- matory cells. Viral respiratory tract infection cuases im- paired mucociliary clearance and edematous ostiomeatal obstructin leading to the development of rhinosinusitis.
11)RV was detected in 21% of epithelial cell samples from patients with CRS and fungi have been found in 6-100%
of CRS.
4)Preexisting RV infection enhance fungal inva- sion with the development of fungal sinusitis. Data has suggested the presence of synergism between RV infec- tion and airborne fungal exposure that enhances the pro- duction of chemical mediators in nasal epithelial cells.
However, to date, there have been no studies reporting on the interaction of fungi and RV in the activation of epithe- lial cells. The aim of this study was to evaluate the effect of Alternaria stimulation of RV-infected nasal polyp epi- thelial cells (NPECs) and to characterize the inflammatory signals that regulate cytokine production.
MATERIALS AND METHODS
NPEC culture
Nasal polyps were obtained from sixteen patients un- dergoing endoscopic sinus surgery. Patients were ex- cluded if they had an allergy or had received systemic or topical steroids or antibiotic and antihistamine medi- cations during the 4 weeks preceding the study. Allergy status were defined with skin prick test or multiple al- lergen simultaneous test-chemiluminescent assay. This study was approved by the Institutional Review Board of Daegu Catholic University Medical Center and each pa- tient signed a consent form that outlined the objectives of the research and experiments. Specimens were placed in Ham’s F12 medium supplemented with 100 international units (IU) of penicillin, 100 ug/ml streptomycin, and 2 ug/
ml amphotericin B, and were transported to the laboratory.
NPECs were isolated by the protease digestion method.
Cell suspensions (106 cells/ml) were plated in wells of six-well culture plates and placed in a 5% CO
2humidified incubator at 37oC. The culture medium was changed after 24 hours and every 2 days thereafter.
RV-16 infection of NPECs and activation with Al- ternaria
RV serotype 16 (RV-16) was purchased from the Amer- ican Type Culture Collection (Manassas, Virginia). Vi- ral stocks were prepared by infection of a sensitive cell monolayer (Ohio HeLa) as described previously (Jang et al., 2009). Second passage human NPECs were seeded into well of 24-well plates at approximately 1X105 cells/
ml and incubated for 24 hours at 37oC. In the RV-16 infec- tion group, the cells were washed with phosphate buffered saline (PBS) and infected with RV-16 at a multiplicity of infection of one for 8 hours in FBS free culture medium.
One multiplicity of infection of RV-16 was adopted for the experiments based on the concentrations used in a previ- ous study, and demonstrated to yield the greatest cytokine release.
12)In the control group, the cells were cultured without RV-16 infection.
After 8 hours, the supernatants were harvested and the cells were washed with PBS then the RV-16-infected epi- thelial cells were activated with Alternata (25 or 50 ug/
ml) (Greer Lab, Lenoir, North Carolina). Endotoxin was removed with EndoClean
TM(BioVintage, San Diego, California). After 48 hours of incubation the supernatants and epithelial cells were harvested and stored at –70oC until assayed. The use of the Alternaria concentration was based on information from a previous study as well as preliminary activation experiments as part of the present study, which showed these concentrations to be non-toxic to the epithelial cells.
2)For inhibition experiments, nuclear factor- κB (NF-κB) inhibitor (BAY 11-7082), activator protein-1 (AP-1) in- hibitor (curcumin), and mitogen-activated protein kinase (MAPK) inhibitor (SB 203580) were purchased from Calbiochem (San Diego, California). Bay 11-7082 and curcumin were dissolved in dimethyl sulfoxide and SB 203580 was dissolved in water. NPECs were pre-treated with transcription factor inhibitors for 1 hour and the su- pernatants and epithelial cells were harvested. The pro- duction of cytokines including interleukine (IL)-6, IL-8, and granulocyte-macrophage colony stimulating factor (GM-CSF) was quantified using commercially available ELISA kits (R&D System, Minneapolis, Minnesota).
After 48 hours incubation of RV-16-infected epitheli- al cells with Alternaria (50 ug/ml), the cell pellets were placed in cryo-tubes and 1 ml Trizol reagent was added.
RNA was extracted according to the manufacturer’s in- structions (Roche Diagnostics, Mannheim, Germany).
The RNA purity and concentration were measured using
a spectrophotometer (Beckman, Mountain View, Cali-
fornia). One microliter of RNA was used for the reverse
transcription polymerase chain reaction amplifications
and performed in a thermal cycle (PerkinElmer, Norwalk,
Connecticut). The primers used in this study were as fol-
lows: IL-6 (sense, CCA GTA CCC CCA GGA GAA GA
and antisense, TTG TTT TCT GCC AGT GCC TC, 191
bp), IL-8 (sense, ACC GGA AGG AAC CAT CTA AC
and antisense, ATT TGG GGT GGA AAG GTT TG, 179
bp), GM-CSF (sense, ATG TTT GAC CTC CAG GAG
CC, antisense, TTG TAG TGG CTG GCC ATC AT, 125
bp), and β-actin (sense, CTG GCC GGG ACC TGA CT,
antisense, GCA GCC GTG GCC ATC TC, 268 bp). The annealing temperature was 57oC for IL-6 and IL-8, and 54 oC for GM-CSF. Samples were visualized using agarose gel electrophoresis and the band densities were measured using the multi Gauge v2.02 software (Fujifilm, Tokyo, Japan). The band intensities were expressed as a percent- age of treated versus untreated cells.
Expression and activation of NF-κB and AP-1 To perform Western blot analysis for NF-κB and AP-1, NE-PER nuclear and cytoplasmic extraction reagents (Pierce Chemical, Rockford, Illinois) was used to iso- late proteins. The protein was quantified using BCATM protein assay reagent kit (Pierce Chemical). The result- ing protein was separated on a NuPAGE 4-12% Bis-Tris
gel (Invitrogen, Carlsbad, California) and transferred onto a nitrocellulose membrane (Bio-Rad Laboratories, Hercules, California). The membrane was blocked with membrane blocking solution (Invitrogen) and probed with anti-human NF-κB p65 subunit, c-Jun, and histone H1 antibodies (Santa Cruz Biotechnology, Santa Cruz, Cali- fornia) at 4oC overnight. After washing, the membranes were incubated with secondary goat anti-rabbit antibody coupled with horseradish peroxidase (HRP) for 1 hour at room temperature, and developed with SuperSignal West Pico Chemiluminescent substrate (Pierce Chemical). The band intensities were expressed as a percentage of treated versus untreated cells.
The nuclear NF-κB and AP-1 activities were quantified using an ELISA-based TransAM kit (Active Motif, Carls- bad, California) according to manufacturer’s protocol. In brief, the nuclear protein extracts were added to a well of 96-well plate that was immobilized with oligonucleotide containing NF-κB and AP-1. NF-κB and AP-1 contained in nuclear extracts bound specifically to the oligonucle- otide during incubation for 1 hour at room temperature.
NF-κB antibody (1:1000 dilution) and AP-1 antibody (1:500 dilution) were then added to each well for 1 hour followed by HRP-conjugated secondary antibody for 1 hour. After adding 100 ul of detection reagent for up to 15 minutes, the colorimetric reaction was stopped. NF-κB and AP-1 activities were determined by reading the absor- bance on a spectrophotometer at 450 nm with a reference wavelength of 655 nm.
STATISTICAL ANALYSIS
Results are expressed as the arithmetic mean ± standard deviation of the mean from five independent experiments.
Inflammatory signal differences between groups was as- sessed by the Kruskal-Wallis test and the cytokine produc- tions and mRNA expression were compared with one-way ANOVA (SPSS ver. 14.0, SPSS, Chicago, Illinois). P val- ues < 0.05 were considered statistically significant.
RESULTS
Effect of Alternaria on cytokine release by RV-16- infected NPECs
Alternaria activated NPECs, causing a significantly re- lease IL-6, IL-8, and GM-CSF (p < 0.05). RV-16 infection alone also induced the production of cytokines. When the RV-16-infected epithelial cells were stimulated with Alter- naria for 48 hours, cytokine production was significantly increased when compared with Alternaria stimulation or RV-16 infection alone (p < 0.05)(Fig. 1).
Fig. 1. Effect of Alternaria and rhinAovirus-16 (RV-16) on IL-6, IL-8, and GM-CSF production in nasal polyp epithelial cells (NPECs).
Alternaria and RV-16 induced significantly larger cytokine pro- duction from the NPECs. Alternaria enhanced the production of cytokines from RV-16-infected NPECs. Results are shown as mean±SD from 5 independent experiments. Alt 50, 10: Alternaria 50 & 10 ug/ml, Alt+RV: activated after RV-16 infection, *: signifi- cant when compared with negative control {(-)}, +: significant when compared with RV-16 infected epithelial cells {(-)+RV}.
600 500 400 300 200 100 0
IL-6 (pg/ml)
(-) Alt 50 Alt 10 (-)+Rv Alt 50+Rv Alt 10+Rv
800 700 600 500 400 300 200 100 0
GM-CSF (pg/ml)
(-) Alt 50 Alt 10 (-)+Rv Alt 50+Rv Alt 10+Rv 900
800 700 600 500 400 300 200 100 0
IL-8 (pg/ml)
(-) Alt 50 Alt 10 (-)+Rv Alt 50+Rv Alt 10+Rv
Effect of Alternaria on the expression and activa- tion of NF-κB and by RV-16 infected NPECs
When NPECs were stimulated with Alternaria for 48 hours, nuclear NF-κB expression was significantly in- creased. RV-16 infection of NPECs did not influence the expression of NF-κB. AP-1 expression was significantly increased with RV-16 infection alone. However Alter- naria did not influence the expression of AP-1 (Fig. 2).
Activation of NF-κB and AP-1 was evaluated using an ELISA-based assay kit incorporating positive controls (jukat nuclear extract for NF-κB, K-562 nuclear extract for AP-1). Nuclear NF-κB activity was significantly in- creased by Alternaria with or without RV-16 infection.
However AP-1 activity was not increased by Alternaria and RV-16 (Fig. 3).
Effect of transcription factor inhibitors on the re- lease of cytokines from RV-16 NPECs
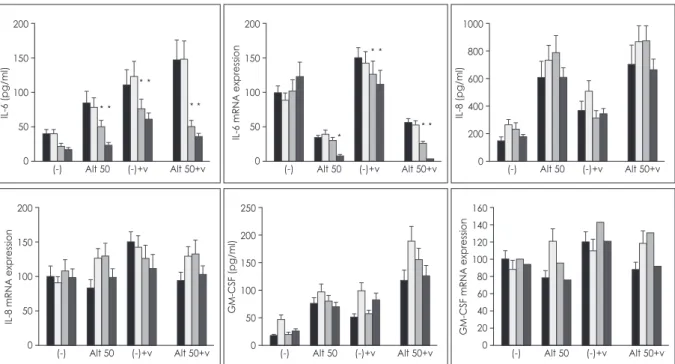
To determine the intracellular signaling mechanism, the optimal concentration of BAY-11-7082 (2.5 uM), SB- 203580 (7.5 uM), and curcumin (10 uM) previously deter- mined to produce the maximum inhibitory effect without cell toxicity.13) The increased IL-6 protein production and mRNA expression, activation of NPEC by Alternaria, RV-16 infection of NPECs, and Alternaria stimulation of RV-16-infected NPECs were significantly inhibited with
NF-κB and AP-1 inhibitors. However, IL-8 and GM-CSF protein production and mRNA expression were not inhib- ited by pretreatment with the transcription factor inhibi- tors (Fig. 4).
DISCUSSION
Fungi and RV cause upper and lower respiratory tract infections and trigger the majority of asthma and chronic obstructive pulmonary disease exacerbations.
1)14)Of the many identified species of fungi, most are not patho- genic. However some of them are associated with airway inflammatory disease. In this study, Alternaria was used to activate NPECs, because Alternaria alternate is one of the most frequently encountered species that is a normal contaminant of nasal secretion, might have strong immu- nologic activity in nasal epithelial cells.
15)Our preliminary experiments with normal nasal epithelial cells showed that the maximal cytokine production was found at 50 ug/ml Alternaria. RV is the most common viral pathogen associ- ated with upper respiratory infection, which can lead to secondary bacterial rhinosinusitis and acute otitis media.
Rhinosinusitis occurs in 0.5% - 2.5% of patients with viral upper respiratory infection due to impaired mucociliary clearance and edematous ostiomeatal obstruction.
16)Case- control studies have shown that RV infection synergizes with allergens in the induction of asthma exacerbations and that airborne fungi modulate the immunologic reac-
Fig. 2. Expression of NF-κB and AP-1 in nasal polyp epithelial cells stimu- lated by Alternaria (50 ug/ml) with or without rhinovirus-16 infection.
NF-κB expression was significantly increased by Alternaria. Results are shown as mean±SD from 5 inde- pendent experiments and repre- sentative electrophoresis results are shown. (-): negative control, Alt: Al- ternaria, V: rhinovirus-16,
*: p < 0.05.
Fig. 3. Intensity of NF-κB and AP-1 activity in nasal polyp epithelial cells stimulated by Alternaria (50 ug/ml) with or without rhinovirus-16 infec- tion. Nuclear NF-κB activity was sig- nificantly increased by Alternaria.
Results are shown as mean±SD from 5 independent experiments. (-):
negative control, Alt: Alternaria, V:
rhinovirus-16, *: p < 0.05.
(-) Alt 50 (-)+v Alt 50+v 200
150 100 50 0
AP-1 Band Density
(-) Alt 50 (-)+v Alt 50+v 3
2.5 2 1.5 1 0.5 AP-1 Optical Density 0 (-) Alt 50 (-)+v Alt 50+v
500 400 300 200 100 0
NF-kB Band Density
(-) Alt 50 (-)+v Alt 50+v 2.5
2 1.5 1 0.5 NF-kB Optical Density 0
tions of RV-infected respiratory epithelial cells.
17)In this study, RV-16 infection and Alternaria enhanced IL-6, IL-8, and GM-CSF production. The production of cy- tokines in NPECs induced by RV-16 infection was sig- nificantly enhanced when the cells were exposed to Al- ternaria. Airborne fungi modulated the RV-16-infected respiratory epithelial cell inflammatory reaction. There was some synergism between RV infection and airborne fungal exposure enhancing the inflammatory response of airway epithelial cells.
NF-κB and AP-1 are key transcription factors that or- chestrate the expression of many genes involved in in- flammation, embryonic development, lymphoid differ- entiation, oncogenesis, and apoptosis.
18)NF-κB regulates the expression of cytokines, growth factors, and effector enzymes in response to ligation of many receptors in- volved in immunity including T-cell receptors, CD40, B- cell receptors, and Toll-like receptors (TLRs). The innate immune response mainly represented by TLRs and Nod- like receptors that recognize their specific ligands activate the transcription factors NF-κB and AP-1, inducing pro- duction of inflammatory cytokines. Even though NF-κB and AP-1 transcription factors are regulated by different mechanisms, they appear to be activated simultaneously by the same multitude of stimuli and many genes require concomitant activation of NF-κB and AP-1, suggesting that these transcription factors work cooperatively.
18)RV infection of bronchial epithelial cells increases vascular
cell adhesion molecule-1 surface expression via NF- κB transcriptional upregulation.
19)Presently, Alternaria strongly induced the expression and activation of NF-κB and RV infection of NPECs did not influence the expres- sion and activation of NF-κB. And the expression of AP-1 was decreased. Intranuclear NF-κB and AP-1 have differ- ent expression patterns. In bronchial epithelial cells, NF- κB expression was increased in a time dependent manner for 48 hours, however AP-1 expression peaked after 6 hours stimulation with fungi then decreased as time went on. (data not shown) Although we did not perform a ki- netic study, AP-1 might increase at early stages after being stimulated with Alternaria.
Respiratory epithelial cells play role in the inflamma- tory response through the release of mediators that may either exert a direct effect on the airway or influence the activation of inflammatory cells.
20)According to the previ- ous studies, nasal polyp may represent more active inflam- matory tissue than healthy normal mucosa with almost same characteristics. So we used nasal polyp epithelial cells instead of normal nasal mucosa.
21) 22)Experiments involving in vitro infection of airway epithelial cells or their stimulation with RV and fungi have demonstrated the induced secretion of several cytokines and chemokines, including IL-1, IL-6, IL-8, GM-CSF, and RANTES.
1) 2) 9)11) 20)
Activation of transcription factors such as NF-κB and AP-1 induces the production of inflammatory cytokines.
RV-16 infection of respiratory epithelial cells leads to a
Fig. 4. Effect of BAY 11-7082, SB 203580 and curcumin on the release of IL-6, IL-8, and GM-CSF and their mRNA expression by Alternaria (50 ug/ml) with or without rhinovirus infection. IL-6 production was significantly inhibited by trascription factor inhibitors but not IL-8 and GM-CSF. Results are shown as mean±SD from 4 independent experiments. (-): negative control, Alt: Alternaria, V: rhinovirus-16, black:
non-treated, gradation: curcumin, dot: SB203580, gray: BAY11-7082, *: p < 0.05.
(-) Alt 50 (-)+v Alt 50+v 200
150 100 50 0
IL-6 (pg/ml)
(-) Alt 50 (-)+v Alt 50+v 200
150 100 50 0
IL-6 mRNA expression
(-) Alt 50 (-)+v Alt 50+v 1000
800 600 400 200 0
IL-8 (pg/ml)
(-) Alt 50 (-)+v Alt 50+v 200
150 100 50 0
IL-8 mRNA expression
(-) Alt 50 (-)+v Alt 50+v 250
200 150 100 50 0
GM-CSF (pg/ml)
(-) Alt 50 (-)+v Alt 50+v 160
140 120 100 80 60 40 20 0
GM-CSF mRNA expression