- 24 -
Expression of GABA Transporters in the Rat’s Olfactory Bulb during Development*
Yong-Jin Park, M.D.
1, Mun-Yong Lee, M.D.
2and He-Ro Yoon, M.D.
1 ABSTRACTDevelopmental changes in the expression of two GABA (gamma-aminobutyric acid) transporter proteins, GAT-1 and GAT-3, in the olfactory bulb of embryonic and postnatal rats were examined with immunocytochemistry using antisera against GAT-1 and GAT-3. The expression and localization of GAT-1 and GAT-3 showed distinct temporal patterns during olfactory bulb dev- elopment. GAT-1 immunoreactivity appeared weakly in most likely growing axons of the presumptive glomerular layer from embryonic day 18 and increased during the first postnatal week. In contrast, GAT-3 immunoreactivity, first detected at E16, was found in radial glial cell fascicles and was replaced by what were likely astroglial cells postnatally. At P7, GAT-1 and GAT-3 immunoreactivities reached the adult pattern i.e., GAT-1 immunoreactivity was observed in the labeled punctate struct- ures in all layers of the olfactory bulb except the nerve fiber layer, while GAT-3 immunoreactivity was observed in the astroglial processes of all layers of the olfactory bulb. Our results suggest that GABA transporters, especially GAT-3, play important roles in regulating the GABA levels of developing olfactory bulbs.
KEY WORDS:GABA transporter (GAT-1, GAT-3)·Immunocytochemistry·Olfactory bulb·Development·Rat.
INTRODUCTION
Gamma-aminobutyric acid (GABA), widely known as an inhibitory neurotransmitter in the mammalian ce- ntral nervous system, is regulated by GABA transporters (GATs) located in the plasma membrane of neurons and glial cells.1)2) These transporters terminate GABAergic transmission through rapid Na+ and Cl- dependent GA- BA uptakes into neurons and surrounding glial cells.3) The transporters may also release GABA into the syna- ptic cleft and extracellular space in a Ca2+-independent, nonvesicular manner.4) At present, at least three distinct GATs (GAT-1, GAT-2, GAT-3) have been cloned. Ph-
armacological and immunocytochemical studies suggest GAT-1 to be predominantly a neuronal transporter, wh- ereas GAT-3 is believed to be a glial transporter in the nervous system.5) In the olfactory bulb, GAT-1 immun- oreactivity is mostly localized to some glial cells and to most periglomerular and granule cells, while GAT-3 im- munoreactivity is expressed in glial cells exclusively.6)
The presence of GABA in both the neurons and the fiber tracts of the early embryonic brain before the on- set of synaptogenesis has been suggested.7)8) The role of GABA in developing tissue is probably different from that in the adult:in developing tissue, GABA likely plays a neurotrophic role or acts as an excitatory neur- otransmitter rather than as an inhibitory transmitter.9)10) Furthermore, recent studies indicate that GABA transp- orter (GAT-1 and GAT-3) mRNAs are transiently exp- ressed in the ventricular and subventricular zones of the developing brain of a mouse.11) Therefore, developmental regulation of the GABA uptake system may be of sig- nificant importance in the developing brain.
The expression of GABA transporters has been obse- rved in the olfactory bulb in the early embryonic stages,12) but the precise localization of each GABA transporter in the olfactory bulb during these stages has not been studied. The objective of this study was to investigate
*This research was supported by the 1998 Graduates’ Associ- ation Research Grant of the Otolaryngology Department, Col- lege of Medicine, The Catholic University of Korea.
1Department of Otolaryngology-Head and Neck Surgery, 2An- atomy, College of Medicine, The Catholic University of Korea, Seoul, Korea
Address correspondence and reprint requests to Yong-Jin Park, M.D., Department of Otolaryngology-Head and Neck Surgery, St. Vincent Hospital, College of Medicine, The Catholic Uni- versity of Korea, 93 Chi-Dong, Pal Dal-Gu, Suwon 442-060, Korea
Tel:82-331-249-7450, Fax:82-331-257-3752 E-mail:[email protected]
Accepted for publication on February 27, 1999
the localization of GAT-1 and GAT-3 in the developing olfactory bulb of a rat by immunocytochemistry.
MATERIALS AND METHODS Subjects
Rat embryos and postnatal pups were obtained from timed pregnant Sprague-Dawley rats. The date of conc- eption [embryonic day (E) 1] was determined through the presence of a vaginal plug. The date of birth was designated as postnatal day 0 (P0). Examinations were conducted at embryonic day 16 (E16), E18, E20, P0, P3, P5, P7, P14, and adulthood. At each time, three an- imals were examined.
Tissue preparation
Embryos [embryonic day 16 (E16), E18 and E20]
were removed from anesthetized (4% chloral hydrate:
10 mg/kg) gravid rats by cesarean section. Postnatal pups at P1 and P3 were anesthetized by inducing hypother- mia, while rats at P5, P7, P14, and adulthood were ane- sthetized with 4% chloral hydrate (10 mg/kg). The heads were removed from E16 embryos, immersed in 4% pa- raformaldehyde (PFA) and 0.2 % picric acid in a 0.1M phosphate buffer (PB, pH 7.4), and cryoprotected in 20% sucrose at 4℃. All other rats were transcardially perfused with 0.01 M phosphate buffered saline (PBS, pH 7.4) followed by 4 % PFA and 0.2% picric acid in PB. The olfactory bulbs were then removed and post- fixed in the same fixative for three hours at 4℃. After several washes in PB, the olfactory bulbs were cryopr- otected in 20% sucrose in PB for one day at 4℃, cut into 12 μm thick coronal sections on a cryostat, colle- cted onto gelatine-coated slides and air-dried.
Immunocytochemistry
For immunocytochemistry, the sections were prein- cubated with 10% normal donkey serum for one hour before being incubated overnight at 4℃ with rabbit polyclonal antibodies to GAT-1 or GAT-3 (Chemicon Inc. Temecula, CA, USA) diluted at 1:50 or 1:75. Sec- tions at E18 were also incubated with mouse monoclonal antibody to vimentin (Boehringer Mannheim, Germany) diluted at 1:50. Bound antibodies were visualized using peroxidase-labeled donkey anti-rabbit antibodies (1:200,
Jackson ImmunoResearch lab, PA, USA) for GAT-1 and GAT-3 labeling or peroxidase-labeled donkey anti-mouse antibodies (1:100, Jackson ImmunoResearch lab, PA, USA) for vimentin labeling with 0.05% 3, 3’-diamino- benzidine tetrahydrochloride-0.01% H2O2 as a detection system. Controls consisted of omitting the primary ant- ibody or incubating with normal rabbit serum in place of the primary antibody.
RESULTS
During development, each GABA transporter in the rat’s olfactory bulb exhibited a distinct temporal pattern.
Omission of primary antibodies or incubation with no- rmal rabbit serum in place of the primary antibody did not result in the specific labeling.
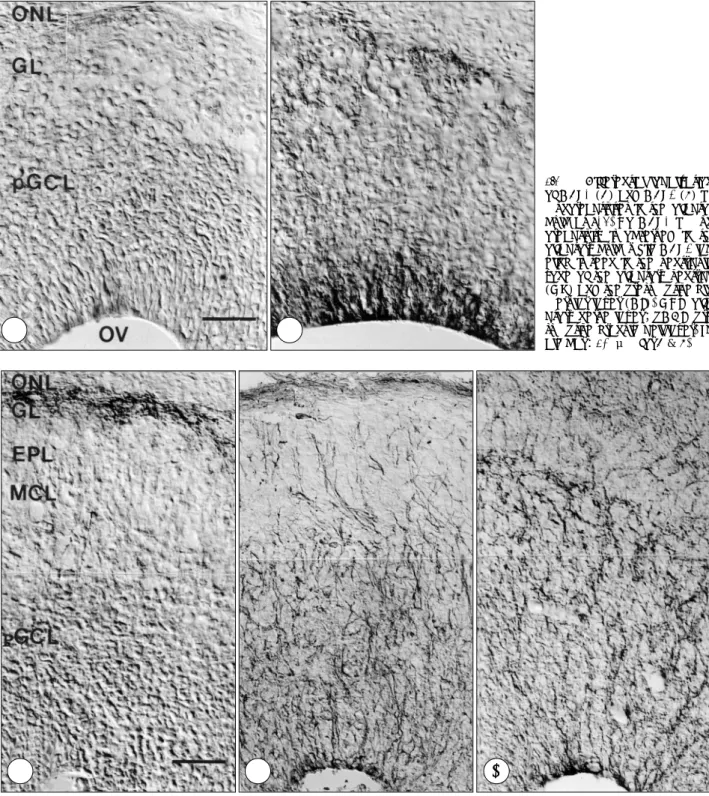
Of the two GABA transporters, GAT-3 immunoreac- tivity was detected earlier. At E16, strong GAT-3 imm- unoreactivity was observed in the ventricular zone of the olfactory ventricle and presumptive glomerular layer, while no GAT-1 immunolabeling was observed (Fig.
1A, 1B). GAT-1 immunoreactivity was first detected at E18 in the presumptive glomerular layer (Fig. 2A). In contrast, GAT-3 immunoreactivity was observed in the radially oriented fibers extending from the olfactory ve- ntricle toward the pial surface. The fibers were identified as radial glial cells by vimentin, the radial glial marker (compare Fig. 2B, 2C). At P0, intense GAT-1 immun- oreactivity was observed in the strongly labeled punctate structures of the granule cell layer as well as the glom- erular layer (Fig. 3A). In the granule cell layer, the la- beling was more prominent in the outer part adjacent to the mitral cell layer. In contrast, GAT-3 immunoreacti- vity was observed in what is believed to be astrocytes and radial glial cells on the basis of their morphology and topography as shown in the olfactory bulb of the neonatal rat13) (Fig. 3B). At this stage, GAT-3 immu- noreactivity was present in the glomerular, external pl- exiform and granule cell layers, while no labeling was observed in the nerve fiber layer. At P7, GAT-1 and GAT-3 immunoreactivities reached the adult pattern.
GAT-1 immunoreactivity was observed in the labeled punctate structures in the glomerular, external plexiform and granule cell layer, but not in the nerve fiber layer (Fig. 4A). GAT-3 immunoreactivity was seen in most likely astroglial processes of all layers of the olfactory bulb, including the nerve fiber, glomerular, external pl-
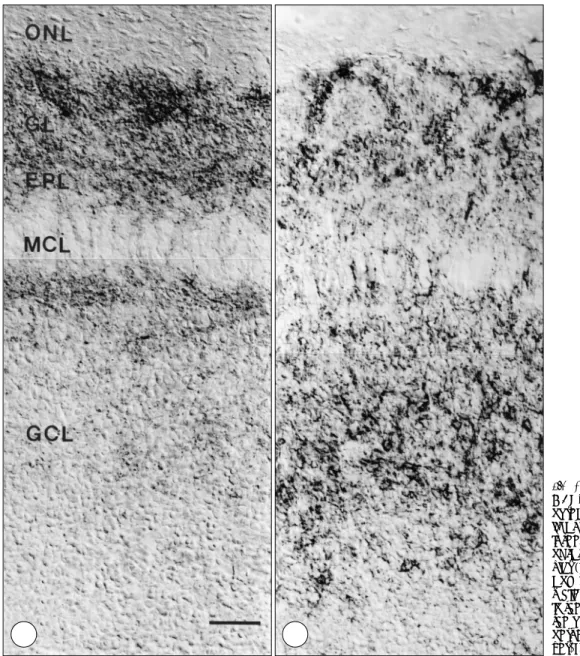
exiform and granule cell layers (Fig. 4B). In the adult, GAT-1 immunoreactivity was observed in the fine gr- anular structures in the glomerular, external plexiform and granule cell layers and was most prominent in the glomerular layer (Fig. 5A). But no immunolabeling was
observed in the nerve fiber layer. Unlike with GAT-1 immunoreactivity, strong GAT-3 immunoreactivity was observed in the likely glial cell processes of all layers of the olfactory bulb, i.e. the olfactory nerve, glomerular, external plexifrom and granule cell layers (Fig. 5B).
Fig. 2. Comparison of GAT-1 (A), vimentin (B) and GAT-3 (C) immunoreactivities in the olfactory bulb at E18. GAT-1 immunoreactivity is observed only in the presumptive glomerular layer (GL), while GAT-3 labeling is seen in the radially oriented fibers extending from the olfactory ventricle toward the pial surface. Note that GAT-3 immunoreactivity is similar to vimentin labeling. ONL, olfactory nerve layer;
EPL, external plexiform layer;MCL, mitral cell layer;pGCL, presumptive granule cell layer. Scale bar:40 μm for A, B and C.
Fig. 1. Differential localizations of GAT-1 (A) and GAT-3 (B) im- munoreactivities in the olfactory bulb at E16. No GAT-1 immun- oreactivity is observed in the olfactory bulb, while GAT-3 lab- eling is seen in the ventricular zone of the olfactory ventricle (OV) and the presumptive glo- merular layer (GL). ONL, olfa- ctory nerve layer;pGCL, pre- sumptive granule cell layer. Sc- ale bar:40 μm for A & B.
AA
AA BBBB
A AA
A BBBB CCCC
DISCUSSION
The present study was undertaken to gain an under- standing of the expression patterns and localizations of GAT-1 and GAT-3 during olfactory bulb development in the rat. In the adult olfactory bulb, GAT-1 immunor- eactivity occurs in fine granular structures, likely axon terminals, in all layers of the olfactory bulb except the nerve fiber layer. In contrast, GAT-3 is observed in the likely astroglial cell processes in all layers of the olfac- tory bulb, including the nerve fiber layer. The expression of the two GABA transporters observed in this study at adulthood is in agreement with all previous studies6)14) except one by Nishimura et al.,6) that reports an expre-
ssion of GAT-1 mRNA in the nerve fiber layer. Durkin et al.,14) however, has found no expression of GAT-1 mRNA in the nerve fiber layer. The reason for the disc- repancy is still unknown, and further studies are needed to determine whether GAT-1 is present in the nerve fiber layer.
The expression and localization of GAT-1 and GAT- 3 show very distinct temporal patterns during olfactory bulb development. Prenatally, GAT-3 immunoreactivity, first detected at E16, is found in radial glial cell fascicles, whereas GAT-1 immunoreactivity is found in most li- kely growing axons from E18. This result is generally consistent with a study by Jursky and Nelson,12) which reports on the expression of GAT-1 and GAT-3 in the developing rat brain, except in the fact that GAT-1 im-
Fig. 3. Differential localizations of GAT-1 (A) and GAT-3 (B) immu- noreactivities in the olfactory bu- lb at P0. GAT-1 immunoreactivity is seen in the strongly labeled pu- nctate structures of the glomer- ular (GL), external plexiform (EPL) and granule cell layers (GCL), while GAT-3 labeling is observed in the likely astrocytes throughout the olfactory bulb. ONL, olfactory nerve layer;MCL, mitral cell la- yer. Scale bar:40 μm for A & B.
A A A
A BBBB
munoreactivity was detected earlier (at E16) in the de- veloping olfactory bulb in that study. Jursky and Nelson’s study does not describe the exact areas in the olfactory bulb where GAT-1 immunoreactivity occurs, so the re-
ason for the discrepancy cannot be easily be determined.
A difference in the type of antibodies, however, is a possible reason.
GAT-1 immunoreactivity is very weak and localized
Fig. 5. Differential localizations of GAT-1 (A) and GAT-3 (B) immun- oreactivities in the olfactory bulb of the adult rat. GAT-1 immunore- activity is observed in the fine gr- anular structures in all layers of the olfactory bulb except in the nerve fiber layer (ONL). In contrast, GAT- 3 labeling is observed in the likely glial cell processes throughout all layers of the olfactory bulb. GL, glomerular layer;EPL, external pl- exiform layer;MCL, mitral cell la- yer;GCL, granule cell layer. Scale bar:40 μm for A & B.
Fig. 4. Differential localizations of GAT-1 (A) and GAT-3 (B) immun- oreactivities in the olfactory bulb at P7. GAT-1 immunoreactivity is observed in the punctate structu- res in the glomerular (GL), external plexiform (EPL) and granule cell layers (GCL), but not in the nerve fiber layer (ONL), while GAT-3 im- munoreactivity is seen in all layers of the olfactory bulb. MCL, mitral cell layer. Scale bar:40 μm for A & B.
AA
AA BBBB
A A A
A BBBB
exclusively to the glomerular layer prenatally and incr- eases during the first postnatal week, while strong GAT- 3 immunoreactivity is transiently observed in the radial glial cells prenatally and replaced by likely astroglial cells postnatally.
In the adult olfactory bulb, periglomerular and granule cells make dendrodendritic GABAergic synapses onto mitral and tufted cells and participate in tuning the pr- operties of odor response through the inhibitory action of GABA on the mitral and tufted cells.15) In the dev- eloping stages, however, GABA depolarizes immature neurons16) and may act as a neurotrophic and chemoki- netic agent17)18) in the developing nervous system. It is likely that GABA is present in a distinct cellular pool, and the presence of uptake processes by GABA trans- porters may be involved in the regulation of GABA co- ncentration. Therefore, the early appearance of GABA transporters, especially GAT-3, in the olfactory bulb may play important roles in regulating GABA levels in the developing olfactory bulb.
CONCLUSION
The expression and localization of GAT-1 and GAT- 3 showed distinct temporal patterns during olfactory bulb development. GAT-1 immunoreactivity appeared weakly in most likely growing axons of the pre-sumptive glo- merular layer from embryonic day 18 and increased po- stnatally, while GAT-3 immunoreactivity, first detected at E16, was found in radial glial cell fascicles and was replaced by likely astroglial cells postnatally. Therefore, it is likely that GABA transporters, especially GAT-3, play important roles in regulating GABA levels in the developing olfactory bulb.
REFERENCES
1) Wood JD, Sidhu HS. A comparative study and partial characteriz-
ation of multi-uptake systems for aminobutyric acid. J Neurochem 1987;49:1202-8.
2) Kanner BI, Bendhan A. Two pharmacologically distinct sodium- and chloride-coupled high-affinity -aminobutyric acid transporters are present in plasma membrane vesicles and reconstituted prepa- rations from rat brain. Proc Natl Acad Sci USA 1990;87:2550-4.
3) Clark JA, Deutch AY, Gallipoli PZ, Amara SG. Functional expre- ssion and CNS distribution of a -alanine-sensitive neuronal GABA transporter. Neuron 1992;9:337-48.
4) Attwell D, Barbour B, Szarkowski M. Nonvesicular release of ne- urotransmitter. Neuron 1993;11:401-7.
5) Guastella J, Nelson N, Nelson H, Czyzyk L, Keynan S, Miedel MC, et al. Cloning and expression of a rat brain GABA transporter. Sc- ience 1990;249:1303-6.
6) Nishimura M, Sato K, Mizuno M, Yoshiya I, Shimada S, Saito N, et al. Differential expression patterns of GABA transporters (GAT1- 3) in the rat olfactory bulb. Mol Brain Res 1997;45:268-74.
7) Lauder JM, Han VKM, Henderson P, Verdoorn T, Towe AC. Pre- natal ontogeny of the GABAergic system in the rat brain: An im- munocytochemical study. Neuroscience 1986;19:465-93.
8) Cobas A, Fairen G, Alvarez-Bolado G, Sanchez MP. Prenatal de- velopment of the intrinsic neurons of the rat neocortex: A compa- rative study of the distribution of GABA-immunoreactive cells and GABA A receptor. Neurosci 1991;40:375-97.
9) Madtes P, Redburn DA. GABA as a trophic factor during develo- pment. Life Sci 1983;33:979-84.
10) Spoerri PE. Neurotrophic effects of GABA in cultures of embry- onic chick brain and retina. Synapse 1988;2:11-22.
11) Evans JE, Frostholm A, Rotter A. Embryonic and postnatal expre- ssion of four gamma- aminobutyric acid transporter mRNAs in the mouse brain and leptomeninges. J Comp Neurol 1996;376:431-46.
12) Jursky F, Nelson N. Developmental expression of GABA transp- orters GAT1 and GAT4 suggests involvement in brain maturation.
J Neurochem 1996;67:857-67.
13) Chiu K, Greer CA. Immunocytochemical analyses of astrocyte development in the olfactory bulb. Dev Brain Res 1996;95:28-37.
14) Durkin MM, Smith KE, Borden LA, Weinshank RL, Branchek TA, Gustafason EL. Localization of messenger RNAs encoding three GABA transporters in rat brain: An in situ hybridization study.
Mol Brain Res 1995;33:7-21.
15) Yokoi M, Mori K, Nakanishi S. Refinement of odor molecule tu- ning by dendrodendtitic synaptic inhibition in the olfactory bulb.
Proc Natl Acad Sci USA 1995;92:3371-5.
16) Cherubini E, Gaiarsa JL, Ben-Ari Y. GABA: An excitatory trans- mitter in early postnatal life. Trends Neurosci 1991;14:515-9.
17) Meier E, Hertz L, Schousboe A. Neurotransmitters as developme- ntal signals. Neurochem Int 1991;19:1-15.
18) Behar TN, Schaffer AE, Colton CA, Somogyi R, Olah Z, Lehel C, et al. GABA-induced chemokinesis and NGF-induced chemotaxis of embryonic spinal cord neurons. J Neurosci 1994;14:29-38.