- 92 -
사람 정상 코점막 상피세포에서 15-Lipoxygenase-1에 의한 점액섬모 분화
연세대학교 의과대학 이비인후과학교실,1 기도 점액연구소,2 두뇌한국 21 의과사업단3
장정현
1·김정홍
1·윤주헌
1,2,3·이정권
1,2·서형석
1·김경수
1,2,315-Lipoxygenase-1 Mediates Mucociliary Differentiation in Normal Human Nasal Epithelial Cells
Jung Hyun Chang, MD1, Jeong Hong Kim, MD1, Joo-Heon Yoon, MD1,2,3, Jeung Gweon Lee, MD1,2, Hyung Seok Seo, MD1 and Kyung-Su Kim, MD1,2,3
1Department of Otorhinolaryngology and 2The Airway Mucus Institute; and 3Brain Korea 21 Project for Medical Science, Yonsei University College of Medicine, Seoul, Korea
ABSTRACT
Background and Objectives:15-lipoxygenase-1 (15-LO-1) is involved in the differentiation of human tracheobronchial epithelial cells. Here we investigated the relation between 15-LO-1 expression and the differentiation of normal human nasal epithelial (NHNE) cells. Materials and Methods:NHNE cells, RT-PCR, Western blot analysis, and scanning electron mi- croscopy (SEM) were used. Results:In retinoic acid (RA)-sufficient culture media, 15-LO-1 expression in NHNE cells in- creased time-dependently, but its expression was undetectable in RA-deficient culture media. Moreover, in RA-deficient culture media, IL-4 time-dependently induced 15-LO-1 expression at a concentration of 1 ng/mL. In addition, MUC8 gene expression, a marker of mucociliary differentiation, was up-regulated by 15-LO-1, which was itself induced by IL-4. In SEM, the ciliated epithelium was observed with the treatment of IL-4. Conclusion:Our findings suggest that 15-LO-1 may be re- lated to the differentiation of human nasal epithelium, and that it may mediate the mucociliary differentiation of NHNE cells.
KEY WORDS:Arachidonate, 15-Lipoxygenase·Nasal mucosa·Cell differentiation·Cilia.
서 론
상기도에서 15-lipoxygenase-1(15-LO-1)의 존재 가능성 에 관한 연구로 사람 코 및 기관지 점막 조직에 대해 고성능 액체 크로마토그라피(high performance liquid chromato- graphy)를 시행하여 이 효소의 대사산물인 15(S)-hydroxyei- cosatetraenoic acid를 검출함으로서 15-LO-1의 존재가 증 명되었고, 사람 기관조직의 상피세포에서의 발현은 면역조 직화학염색을 이용하여 이의 국소화(localization)가 보고 되었으며, 사람 코점막에서의 국소화는 최근 면역조직화학
염색을 통해 존재부위가 밝혀졌다.1-4)
15-LO-1은 적혈구, Caco-2 세포, 사람 기관-기관지 상피 세포 등의 분화에 관여한다.5-7) 사람 기관-기관지 상피세포 배양에서 15-LO-1은 세포 분화에 직접 관여하며, 일부 점 막세포에서는 효소를 활성시킨다.7)8) 한편 사람 기관 조직과 코는 같은 상기도에 속하는 섬모 상피세포이므로, 15-LO- 1이 사람 기관-기관지 상피세포의 성장과 분화에 관여한다 는 것으로 미루어 볼 때 사람 코점막 상피세포의 분화에 관여할 가능성을 추측할 수 있다.
사람 기관-기관지 상피세포에서 15-LO-1의 발현과 점액 분비는 IL-4에 의해 조절된다.9) 그리고 여러 점액 유전자 중 MUC 8은 사람 코점막 상피에서 섬모 상피세포의 표식 자로 간주된다.10)11) 그러므로, 15-LO-1이 코점막 상피 세포 에서 IL-4에 의해 조절되고 IL-4에 의한 변화 특히 MUC8 의 변화가 일어난다면 사람 코점막 상피세포의 분화에 15- 심사완료일:2005년 8월 24일
교신저자:김경수, 135-720 서울 강남구 도곡동 146-92 연세대학교 의과대학 이비인후과학교실, 기도 점액연구소, 두뇌한국 21 의과사업단
전화:(02) 3497-3463・전송:(02) 3463-4750 E-mail:[email protected]
LO-1이 관여한다는 단서를 제공할 수 있다 하겠다. 이에 본 연구를 통해 사람 정상 코점막 상피세포의 분화에 따라 15- LO-1 이 어떻게 발현하는 가를 알고자 하였으며, 사람 정상 코점막 상피세포에서 IL-4에 의해 15-LO-1이 유도되는 가를 알고자 하였다. 만일 15-LO-1이 IL-4에 의해 조절된 다면 IL-4에 의한 MUC8유전자의 발현을 조사하여 IL-4 에 의해 유도되는 MUC8과 15-LO-1의 관계를 보고자 하 였다. 또한 주사전자현미경으로 관찰을 하여 15-LO-1을 매 개로 한 형태학적 변화를 알고자 하였다.
재료 및 방법
세포 배양
0.45 mm 크기의 pore를 가진 Transwell-clear(Costa Co., Cambridge, MA, USA) culture insert(diameter 24.5 mm) 에 passage-2 사람 정상 코점막 상피세포를 배양하였다 (105 cells/culture). 사용된 배양액은 BEGM과 DMEM을 1:
1로 섞고, 여기에 fetal bovine serum을 첨가하였으며, re- tinoic acid(RA)(Sigma Co: St. Louis, MO, USA)는 10-7 M 농도로 사용하였다.11) 막의 위-아래에 모두 배양액을 넣어 9일간 배양하였으며, 9일째 막 상부의 배양액을 제거하여 air-liquid interface(ALI)법을 시행하여 아래로부터 영양 분을 받도록 배양하였다.
Western blot analysis
배양된 세포를 1 ml의 radioimmunoprecipitation assay buffer(1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS) 을 이용하여 세포 용해액을 얻었다. 이 세포 용해액을 4℃
에서 20분간 10,000×g로 원심분리하여, 침전된 총단백으 로 Western blot 분석을 시행하였다. 단백농도는 소 혈청 알 부민을 이용하는 BCA protein assay로 농도를 결정하였다.
단백(30 μg)을 8% SDS-polyacrylamide gel electropho- resis(PAGE)로 분리한 후 니트로셀룰로즈막(Amersham Phar- macia Biotech:Piscataway, NJ, USA)에 전이하였다. 막을 10% 탈지분유가 함유된 TBST(Tris-buffered saline/0.05%
Tween-20) 용액에서 4℃로 하룻밤 동안 담가두어 비특이 반응을 억제하였고, 2% 탈지분유가 포함된 TBST를 이용하 여 1:400의 농도로 항15-LO-1 항체를 사용하거나 1:500 의 농도로 항actin 항체를 사용하여 실온에서 4시간 동안 반응시켰다. 세척을 시행한 후 막은 TBST에 1:5,000으로 희석한 horse reddish peroxidase 항체(Amersham Phar- macia Biotech)로 한 시간 동안 처리하고 수 차례의 세척 후 enhanced chemiluminescence(Amersham Pharmacia Bio-
tech)와 autoradiography를 이용하여 밴드를 확인하였다.
Autoradiography에서 얻어진 밴드의 세기를 Scion Image (Scion Co., Frederick, MD, USA)를 이용하여 수치화하였다.
Reverse transcription-polymerase chain reaction (RT-PCR)
15-LO-1과 MUC8에 대한 oligonucleotide primer를 이 미 알려진 염기 서열을 토대로 만들었다.12)13) β2-micro- globulin(β2M)에 대한 oligonucleotide amplimers는 RT- PCR의 대조군으로서 Clontech Laboratories(Palo Alto, CA, USA;a 335-bp PCR fragment)에서 구입하였다. RT-PCR 은 Perkin-Elmer Cetus DNA Thermal Cycler(Perkin- Elmer, Norwalk, CT, USA)를 이용하여 시행하였다. 방법 을 약술하면 총RNA(20 μL의 반응용량 당 1 μg)를 ran- dom hexanucleotide primers와 Moloney murine leukemia virus reverse transcriptase(RT)를 이용하여 complemen- tary DNA(cDNA)로 역전사하였다. 95℃에서 1분간 변성을 시행하였고 15-LO-1에 대해 58℃에서 1.5분, MUC8에 대 해 55℃에서 1분간 결합하였다. 연장반응은 15-LO-1에 대해 72℃에서 1.5분, MUC8에 대해 1분간 시행되었다. 15- LO-1과 MUC8 mRNA 발현을 비교하기 위해 비교역학분석 (comparative kinetic analysis)을 시행하였다. PCR 산물은 50 ng/mL ethidium bromide를 함유하는 2% Seakem aga- rose gel(FMC, Rockland, ME, USA)에서 전기 영동으로 분 리하여 폴라로이드 타입 55 필름으로 사진을 찍었다. 이후 PCR 산물의 강도는 Molecular Dynamics Densitometer (Sunnyvale, CA, USA)를 이용하여 확인하였고, Image- Quant software를 이용하여 결과를 분석하였다. 최종 산물 (amplified products)이 mRNA에서 생기고 genomic DNA 의 오염이 없었다는 것을 증명하기 위해 역전사 반응에서 역 전사 효소(reverse transcriptase)를 생략하여 PCR산물이 관찰되지 않음을 확인함으로써 음성 대조군(negative control) 으로 삼았다. 표적 유전자의 증폭은 sequencing(dsDNA Cycle Sequencing System, Gibco-BRL, Carlsbad, CA, USA)으 로 확인하였다. 결과는 각 실험의 강도를 해당하는β2M의 강도와 비교한 후 이 수치를 대조군과 비교하여 비율로 나 타내었다. 실험은 동일한 방법으로 3회 이상 실시하였으며 통계 검증은 Mann-Whitney test를 이용하여 유의수준은 p<0.05를 유의한 것으로 하였다.
주사전자현미경 관찰
주사 전자현미경 관찰을 위해서 insert의 막을 칼로 절제 한 후, 막위에 배양된 상피세포를 2.5% glutaraldehyde에
4℃에 4~6시간 고정한 후 0.1M phosphate buffer로 세척 하였다. 1% osmium tetroxide에 1시간 동안 다시 고정한 다음 금방 만들어 여과한 2% tannic acid에 4℃에 밤새 처 리한 후 다시 1% OsO4에 30분간 고정하였다. 탈수과정을 거쳐 critical point drying 시킨 후 gold coating하여 Mis- 2400 주사전자현미경(Hitachi CO., Japan)으로 관찰하였다.
결 과
Retinoic acid에 의한 15-LO-1의 발현
분화 정도에 따른 15-LO-1의 발현을 알아보기 위해 RA를 첨가한 경우와 RA를 첨가하지 않은 경우에 각각 배 양된 사람 정상 코점막 상피세포를 Western blot 분석하였 다. ALI 배양 7일 후 RA를 첨가한 사람 정상 코점막 상피 세포의 경우 ALI 배양 0일과 비교 시 15-LO-1의 발현이 약 3배 정도 높았으며, 발현은 배양시간에 따라 증가하였 다. RA를 첨가하지 않은 경우는 배양시간과 무관하게 15- LO-1이 발현되지 않았다(Fig. 1A). 이처럼 15-LO-1의 발 현이 RA를 첨가한 경우 배양 시간에 따라 증가하므로 15- LO-1유도에 있어 RA의 영향을 배제하기 위하여 RA를 첨 가하지 않은 실험조건으로 아래의 실험을 진행하였다.
IL-4에 의한 15-LO-1의 발현
IL-4에 의한 15-LO-1의 발현 여부를 알아보기 위해 RA 를 첨가하지 않은 조건에서 ALI 배양 직후부터 IL-4를 첨 가한 경우와 그렇지 않은 경우로 각각 배양 하여 Western blot 분석을 하였다. 1 ng/ml의 IL-4가 첨가된 경우 배양 3일째부터 15-LO-1이 발현되기 시작하여 발현이 배양 시 간에 따라 증가하여 배양 14일째 가장 강한 발현을 보였다.
그러나 IL-4가 첨가되지 않은 배양의 경우 15-LO-1이 배 양 시간과 무관하게 발현되지 않았다(Fig. 1B).
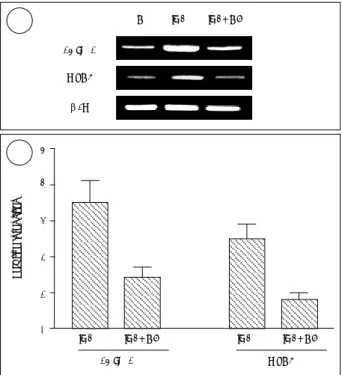
IL-4에 의한 MUC8의 발현과 15-LO-1 발현과의 연관성 IL-4에 의한 MUC8의 발현과 15-LO-1 발현과의 연관 성을 알아보기 위하여 RA를 첨가하지 않은 조건에서 ALI 배양 직후부터 14일간 각 조건 하에 세포를 배양하였다. 배 양 조건은 IL-4와 caffeic acid 모두를 처치하지 않은 대조 군, 1 ng/ml IL-4만을 첨가한 군, 1 ng/ml IL-4와 15- LO-1 억제제인 2.2 μM caffeic acid 모두를 첨가 배양한 군 등의 세 군으로 구분하였으며, caffeic acid는 IL-4 처
Fig. 1. Expression of 15-LO-1 versus culture time. Expressions were determined by Western blotting on normal human nasal epi- thelial cells grown in RA-sufficient (RA+) or RA-deficient (RA-) media. 15-LO-1 expression values are shown as ratios versus actin using the densitometer intensities of hybridization bands.
A. In RA-sufficient culture, 15-LO-1 expression is three-fold higher on culture day 7 than on culture day zero, and this expression further increases with time. In RA-deficient culture, 15-LO-1 expression is undetectable regardless of culture duration B. In RA-deficient media, cells were treated (+) or not treated (-) with 1 ng/ml IL-4 for 14 days at the air-liquid interface, and 15- LO-1 expression was followed by Western blotting. In the pre- sence of IL-4, 15-LO-1 expression is observed from culture day 3 and then increases with time.
0.8 1.2 2.0 15-LO-1
Actin Ratio
0 day
1 day 3 day 7 day 14 day
- + - + - + - + IL-4 RA
0 day 7 day 14 day
- + - +
- + 15-LO-1
Actin
Ratio 0.1 0.3 0.9
A AA A
BBB B
15-LO-1 MUC8
β2M
C IL-4 IL-4+CA
Fig. 2. The up-regulation of 15-LO-1 and MUC8 at the gene level by IL-4. NHNE cells were cultured in RA-deficient media containing 1 ng/ml IL-4 or 1 ng/ml IL-4+2.2 μM caffeic acid for 14 days at the air-liquid interface. Total RNA was obtained and RT-PCR performed. Expression values of the mRNA are calculated as ratios versus β2M using the densitometer in- tensities of bands. After then, the relative intensities of the ex- pression values were measured compared to those of each control. 15-LO-1 and MUC8 mRNA expressions are found to be up-regulated by IL-4 but these are held at control levels co- treating with IL-4+caffeic acid. A is a representative of three separate experiments and the relative intensities in B are the means±standard deviations of three experiments.
IL-4 IL-4+CA IL-4 IL-4+CA
15-LO-1 MUC8
Relative intensities
5 4
3 2
1 0
A A A A
B B B B
치 1시간 전에 전처치하였다. 각 조건 하에서 14일간 처치 후 총 RNA를 추출하였고, 이를 이용하여 RT-PCR을 시행 하였다.
대조군과 비교 시 IL-4만을 첨가한 경우 15-LO-1의 발 현 비율은 3.5±0.6이었고, IL-4와 caffeic acid 모두를 첨 가한 경우 15-LO-1의 발현 비율은 1.4±0.3으로 나와 caf- feic acid에 의해 유의하게 15-LO-1이 억제되는 것을 알 수 있었다(p<0.05). MUC8의 발현을 보면 IL-4만을 첨가 한 경우 대조군과 비교한 MUC8의 발현 비율은 2.5±0.4 이었고, IL-4와 caffeic acid 모두를 첨가한 경우 MUC8의 발현 비율은 0.8±0.2로 나와 caffeic acid에 의해 유의하 게 MUC8이 억제되었다(p<0.05)(Fig. 2).
IL-4에 의한 형태학적 변화
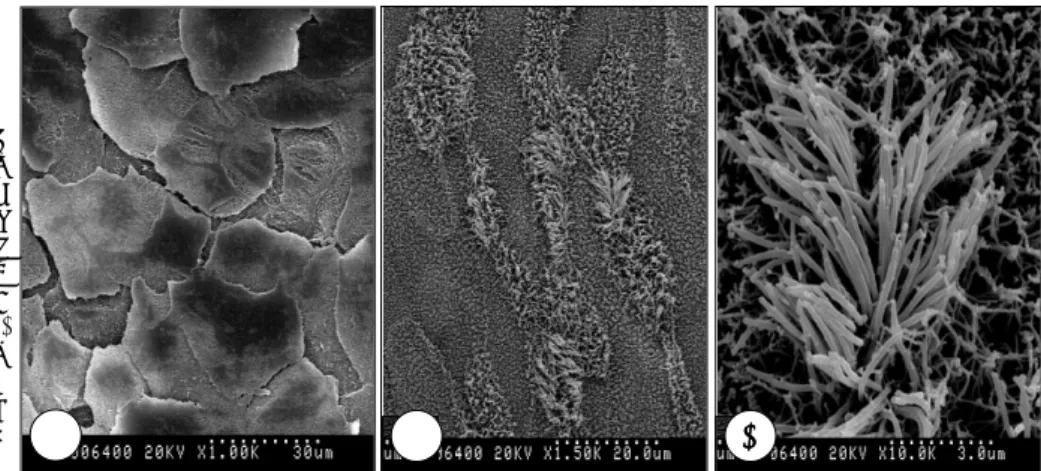
ALI 배양 직후부터 14일간 1 ng/ml IL-4를 첨가한 군과 이를 첨가하지 않은 대조군으로 나누어 배양한 후 각각을 주 사전자현미경으로 관찰하였다. 대조군의 경우 섬모가 없는 편평 상피세포가 관찰되었으나, IL-4를 첨가 배양한 경우 일부에서 섬모가 있는 섬모 상피세포가 관찰되었다(Fig. 3).
고 찰
NHNE세포의 경우 RA는 점액섬모 상피세포로의 분화에 필수적인 물질로, RA를 첨가하지 않고 배양한 경우 사람 정 상 코점막 상피세포는 편평 상피세포로 분화하게 되고 RA 를 첨가한 경우에만 점액섬모 상피세포로 분화화게 된다.14) 본 연구결과 상 RA를 첨가하고 배양한 경우 시간 경과에 따 라 15-LO-1이 발현되고, RA를 첨가하지 않은 경우 15- LO-1이 발현되지 않았다. 이런 결과로서 RA가 첨가되어 사 람 정상 코점막 상피세포가 점액섬모 세포로 분화하는 과정 에 15-LO-1이 증가하여 15-LO-1이 점액섬모 분화에 관
여함을 알 수 있었다. 그러나 이 결과는 15-LO-1이 NHNE 세포의 점액섬모 분화와 연관됨을 알려주는 것이고 15-LO- 1이 점액섬모 분화를 매개한다는 직접적 증거는 아니므로, 이에 RA의 영향을 배제하기 위해 RA를 첨가하지 않은 조 건에서 15-LO-1이 NHNE세포 분화에 어떤 직접적 영향 을 주는 가에 대해 연구를 시작하였다.
본 연구결과에서 RA를 첨가하지 않은 조건에서 배양 시 시간 경과에 따라 사람 정상 코점막 상피세포에서 IL-4에 의해 15-LO-1가 발현되었다. 이러한 결과는 사람 정상 기 관-기관지 세포에서 시간과 용량에 따라 IL-4가 15-LO- 1을 유도한다는 보고와 일치하였다.9) 이 결과에 기초하여 본 연구에서 IL-4를 이용하여 15-LO-1을 유도하였고, 또 한 점액섬모 분화의 표식자인 MUC8의 유도 여부 및 15- LO-1 억제제인 2.2 μM의 caffeic acid를 이용하여 15- LO-1과 MUC8의 연관성을 보고자 하였다.10)11)15)
1 ng/ml IL-4으로 NHNE세포를 처치한 후 15-LO-1의 발현이 시간 경과에 따라 증가하였으며, 반면1 ng/ml IL- 4과 2.2 μM caffeic acid로 함께 처치하였을 경우 억제되 었다. 이와 유사하게 MUC8의 발현은 IL-4에 의해 증가하 였으며, IL-4와 caffeic acid를 함께 처치한 경우 MUC8의 발현이 억제되었다. 이러한 결과는 MUC8의 발현이 15-LO- 1에 의해 조절되며, MUC8이 점액섬모 분화의 표식자이므 로 15-LO-1이 사람 정상 코점막 상피세포에서 점액섬모 분 화를 매개한다는 것을 의미한다. 또한 15-LO-1에 대한 이 러한 결과는 백서 코점막의 발육에 백혈구형 12-LO가 관 여한다는 사실과 맥을 같이 하는 결과로, 백서에 대한 연구 에서 15-LO-1이 사람 정상 코점막 상피세포의 분화에 관 여할 것이라는 추측에 대한 직접적 증거라 하겠다.16)
주사전자현미경으로 형태를 관찰한 결과 RA가 없는 조 건에서 IL-4를 첨가하지 않은 경우 편평상피가 관찰되었 으나, IL-4를 첨가배양한 경우 RA가 없는 상태에서도 섬
Fig. 3. Scanning electron microscopic findings. NHNE cells were cultured in RA-deficient media for 14 days at the air-liquid interface. A:Squamous epi- thelium is observed in NHNE cell cul- ture omitting IL-4 (×1000). B:Ciliat- ed epithelial cells are noted in NHNE cell culture containing 1 ng/ml IL-4.
Ciliated cells are found in some por- tions and the most areas are squa- mous cells (×1000). C:The magnified photo of Fig. 3B, which shows cilia (×
2500). AAAA BBBB CCCC
모가 있는 점액섬모 분화가 관찰되었다. 본 연구에서 계량 적인 방법으로 연구를 하지 않아 정확한 수치는 알 수 없 지만 일부에서만 섬모 상피세포로 분화가 되어, RA를 준 경우 ALI 배양 14일째 섬모 상피세포가 전체 세포 중 7.4
±0.5%란 결과와 비교시 양적으로 분화가 적은 양상이었 다.10) 이런 결과는 IL-4가 RA가 없는 상태에서도 섬모 상 피세포로의 분화를 유도하지만 RA를 투여한 것만큼 강력 하지 않다는 것을 의미하며 향후 IL-4와 RA의 분화 유도 능에 대한 연구가 필요하리라 생각한다.
결 론
사람 정상 코점막 상피세포의 경우 RA가 없는 조건 하에 IL-4는 15-LO-1을 유도하였고, IL-4에 의한 15-LO-1의 증가에 의해 MUC8 유전자의 증가와 점액섬모상피로의 분 화가 관찰되었다. 이러한 결과로 15-LO-1이 사람 정상 코 점막 상피세포의 분화에 관여하며 특히 점액섬모상피로의 분화를 유도한다고 생각한다.
중심 단어:15-Lipoxygenase・코점막・세포 분화・섬모.
REFERENCES
1) Henke D, Danilowicz RM, Curtis JF, Boucher RC, Eling TE.
Metabolism of arachidonic acid by human nasal and bronchial epithelial cells. Arch Biochem Biophys 1988;267:426-36.
2) Nadel JA, Conrad DJ, Ueki IF, Schuster A, Sigal E. Immunocy- tochemical localization of arachidonate 15-lipoxygenase in erythro- cytes, leukocytes, and airway cells. J Clin Invest 1991;87:1139-45.
3) Shannon VR, Crouch EC, Takahashi Y, Ueda N, Yamamoto S, Holtzman MJ. Related expression of arachidonate 12- and 15-li- poxygenase in animal and human lung tissue. Am J Physiol 1991;
261:L399-L405.
4) Kim KS, Yoon JH, Kim CH, Kim SG, Chang JH, Kim JH. Ex- pression of 15-lipoxygenase and cyclooxygenase-2 in human nasal mucosa and cultured human nasal epithelial cells. Korean J Oto- laryngol 2003;46:649-53.
5) Kuhn H, Walther M, Kuban RJ. Mammalian arachidonate 15-li- poxygenase structure, function, and biological implications. Pros- taglandins Other Lipid Mediat 2002;68-69:263-90.
6) Grullich C, Duvoisin RM, Wiedmann M, van Leyen K. Inhibition of 15-lipoxygenase leads to delayed organelle degradation in the reticulocyte. FEBS Lett 2001;489:51-4.
7) Kumlin M, Hamberg M, Granstrom E, Bjorck T, Dahlen B, Matsu- da H, et al. 15 (s)-hydroxyeicosatetraenoic acid is the major ara- chidonic acid metabolite in human bronchi: Association with airway epithelium. Arch Biochem Biophys 1990;282:254-62.
8) Hill EM, Eling T, Nettesheim P. Differentiation dependency of eicosanoid enzyme expression in human tracheobronchial cells.
Toxicol Lett 1998;96-97:239-44.
9) Jayawickreme SP, Gray T, Nettesheim P, Eling T. Regulation of 15-lipoxygenase expression and mucus secretion by IL-4 in human bronchial epithelial cells. Am J Physiol 1996;276:L596-L603.
10) Yoon JH, Moon HJ, Seong JK, Kim CH, Lee JJ, Choi JY, et al.
Mucociliary differentiation according to time in human nasal epi- thelial cell culture. Differentiation 2002;70:77-83.
11) Kim CH, Kim HJ, Song KS, Seong JK, Kim KS, Lee JG, et al.
MUC8 as a ciliated cell marker in human nasal epithelium. Acta Otolaryngol 2005;125:76-81.
12) Nassar GM, Morrow JD, Roberts J, Lakkis FJ, Badr KF. Induction of 15-lipoxygenase by interleukin-13 in human blood monocytes. J Biol Chem 1994;269:27631-4.
13) Kim SS, Kim KS, Lee JG, Park IY, Koo JS, Yoon JH. Levels of intracellular protein and mRNA of mucin and lysozyme in normal human nasal and polyp epithelium. Laryngoscope 2000;110:276-80.
14) Yoon JH, Kim KS, Kim SS, Lee JG, Park IY. Secretory differen- tiation of serially passaged normal human nasal epithelial cells by retinoic acid: expression of mucin and lysozyme. Ann Otol Rhinol Laryngol 2000;109:594-601.
15) Shureiqi I, Chen D, Lotan R, Yang P, Newman RA, Fischer SM, et al. 15-lipoxygenase-1 mediates nonsteroidal anti-inflammatory drug- induced apoptosis independently of cyclooxygenase-2 in colon cancer cells. Cancer Res 2000;60:6846-50.
16) Chun HS, Yoon JH, Lee JH, Lee YJ, Park TJ, Kim KS. Expression of leukocyte-type 12-lipoxygenase in murine nasal mucosa accor- ding to the development. Korean J Otolaryngol 2005;48:40-5.