- 19 -
Distribution of IL-4, IL-8 and GM-CSF in Nasal Turbinate Mucosa Following Sulfur Dioxide Exposure In Vitro
Chul Hee Lee, M.D.
1, Kang Soo Lee, M.D.
2, Chae-Seo Rhee, M.D.
1and Byeong Ho Song, M.D.
1 ABSTRACTThere is increasing evidence that airway epithelial cells, when exposed to various gas-derived air pollutants, play an important role in airway inflammation by releasing inflammatory cytokines. However, there is little information on air pollutant-induced cytokine expression at the tissue level and on the role of sulfur dioxide (SO2), one of the major ambient air pollutants, in cy- tokine production. We studied whether or not a low concentration of sulfur dioxide induces an increase in tissue expression of interleukin-4 (IL-4), interleukin-8 (IL-8), and granulocyte/macrophage colony stimulating factor (GM-CSF). After exposing surgically obtained normal human nasal turbinates to 0.05 ppm SO2 for one hour, we conducted specific immunohistochemical staining to assess the tissue expression of each cytokine. We found that the percent expression of IL-8 and GM-CSF in the su- rface epithelium was significantly higher in each SO2-exposed tissue than in the matched control tissue. However, there was no significant difference in the number of submucosal IL-4-positive cells between exposed and control specimens. These results suggest that exposure to a low concentration of SO2 increases airway inflammation, apparently by inducing an increase in the expression of GM-CSF and IL-8.
KEY WORDS:Sulfur dioxide·IL-4·IL-8·GM-CSF·Nasal epithelium.
INTRODUCTION
As more is becoming known about its associations with increased respiratory morbidity and mortality, air pollution is becoming an increasingly significant public health concern worldwide. Both epidemiological and la- boratory studies have generated evidence suggesting that increased exposure to various gas-derived air pollutants, such as nitrogen dioxide (NO2), ozone (O3) and res- pirable particulate matter, plays a role in the clinical manifestation of both allergic and non-allergic airway inflammation.1) Though the mechanisms and cell types involved in air pollutant-induced airway inflammation have not been fully established, it is well known that airway epithelial cells play an active role in the allergic
airway inflammatory response by secreting pro-inflam- matory cytokines.2-6) Little is known, however, about air pollutant-induced cytokine expression at the tissue level.
Our study was conducted to determine whether or not a low concentration of sulfur dioxide (SO2), one of the major ambient air pollutants, induces a significant incr- ease in the tissue expression of granulocyte/macrophage colony stimulating factor (GM-CSF), interleukin-8 (IL- 8) and interleukin-4 (IL-4) in cultured human nasal turbinate.
MATERIALS AND METHODS Subjects
Six healthy nonatopic nonrhinitic subjects with nasal septal deviation were recruited for the study. None of the subjects had experienced symptoms of upper resp- iratory tract infection within the preceding one month.
The absence of allergy was determined by the absence of the main symptoms of allergic rhinitis and by nega- tive results on a skin prick test and/or radioallergosor- bent test. This study was approved by the Seoul National University Ethics Committee. All subjects were given written informed consent forms to complete prior to the
1Department of Otorhinolaryngology, College of Medicine, Seoul National University, Seoul,
2Department of Otorhinolaryngology, College of Medicine, Hallym University, Hallym University Sacred Heart Hospital, Anyang, Korea
Address correspondence and reprint requests to Chul Hee Lee, M.D., Department of Otorhinolaryngology, Seoul National Un- iversity Hospital, 28, Yongon-dong Chongno-ku, Seoul 110-744, Korea
Tel:82-2-760-3646, Fax:82-2-745-2387 E-mail:[email protected]
Accepted for publication on March 24, 1999
study.
Nasal biopsy, gas exposure, and tissue proces- sing
Specimens of nasal mucosa were obtained from the inferior turbinates during septoplasty operations. The ti- ssues were then washed in normal sterile saline solutions;
immediately placed in a prewarmed culture medium, Du- lbecco’s modified Eagle Medium-Ham’s nutrient F12 (DMEM-F12;Gibco BRL, Grand Island, NY);tran- sferred to the laboratory;divided into several sections as needed;and placed in a 37℃, 5% CO2 incubator for 24 hours. We used an in vitro system with the explant cell culture technique. The explant-containing culture dish was placed in a specially designed acryl gas chamber (30×20×50 cm). The chamber was supplied with a mixture of SO2 and filtered synthetic air through a gas- blending chamber. A constant air flow of 3 L/min and a temperature of 37℃ were maintained during the expo- sure stage. Each explant tissue was exposed to 0.05 ppm SO2 for one hour. The concentration of the SO2 was checked with gas detector tubes (GV-100S, Gastec, Ja- pan). Exposed tissues were frozen at six hours after the exposure to SO2, and serially sectioned into 4 μm thi- cknesses. The control specimen was exposed only to the culture media and frozen at the same time as the mat- ched exposed specimen.
Immunohistochemistry and quantitation of im- munoreactivity
The sections of tissue were stained using the strepta-
vidin-biotin peroxidase system. The primary monoclonal antibodies used in this study were mouse anti-human IL- 4 (Genzyme, Cambridge, MA), IL-8 (Genzyme, Cam- bridge, MA), and GM-CSF (Genzyme, Cambridge, MA).
After preincubation with a blocking antibody in a DA- KO LSAB kit (DAKO Ltd., Glostrup, Denmark) for 30 minutes at room temperature, the sections were incuba- ted with one of the following:mouse anti-human IL-4, diluted 1:2,000, mouse anti-human IL-8, diluted 1:20 or mouse anti-human GM-CSF, diluted 1:2,000. After washing in PBS, the sections were then incubated with biotin-labelled goat anti-mouse IgG (1:200) for 30 minutes at room temperature, washed again in PBS, and subsequently conjugated by avidin horseradish perox-I- dase for 45 minutes at room temperature. They were then washed and stained for peroxidase activity using 3,3-diamino-benzidine-tetrahydrochloride (DAB) (Si- gma, St. Louis, MO).
The length of the positively stained epithelium was expressed as a percentage of the total length of the ep- ithelium to quantitate the immunoreactivity of IL-8 and GM-CSF. Using an eye reticule, we counted IL-4-pos- itive submucosal cells in five high power fields of the lamina propria under 400X magnification. The mean value of the immunoreactivity findings in the five fields was recorded as the result.
Statistical analysis
The differences in cytokine expression between the control and exposure groups were analyzed using the Wilcoxon signed rank test. The acceptable level of sig-
Fig. 1. IL-8 expression at six hours after one-hour exposure to 5% CO2 in air (A) and to SO2 (B). Following exposure to SO2, immunore- activity to anti-IL-8 antibody is observed on the surface epithelial cells (×200).
AAA
A BBBB
nificance was set at p<0.05.
RESULTS
IL-8 (Fig. 1) and GM-CSF (Fig. 2) expression was higher predominantly in the surface epithelium following exposure to SO2, whereas IL-4-positive cells were pre- dominantly localized in the lamina propria irrespective of exposure to SO2 (Fig. 3). The percent expression of IL-8 in the surface epithelium was significantly higher in the SO2-exposed tissues than in the controls (40.5±
14.2% vs 78.2±8.8%, p=0.001) (Fig. 4). Likewise, GM-CSF expression in the surface epithelium was sig- nificantly higher after the exposure to SO2 (25.8±7.2%
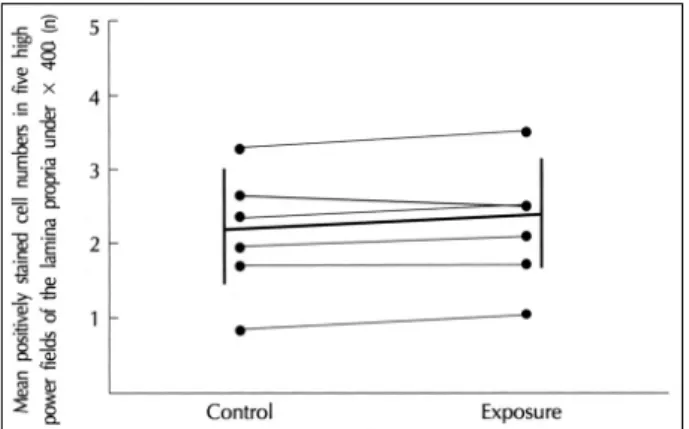
vs 82.5±7.7%, p<0.001) (Fig. 5). However, there was no significant difference in the number of submucosal IL-4-positive cells between the exposed and control sp-
ecimens (2.1±0.8 vs 2.2±0.8, p=0.104) (Fig. 6).
DISCUSSION
In this study we set the concentration of and exposure time to SO2 at 0.05 ppm (Standard Korean air quality demonstrates an SO2 concentration of 0.25 ppm.) and one hour, respectively, because 0.05 ppm SO2 did not cause the epithelial desquamation of cultured turbinate explants. This may be attributable to the possibility that a low concentration of SO2 leads to morphologic chan- ges in the nasal respiratory epithelium by reducing the protective function of the in vitro condition.
Although we did not measure the solubility of SO2
in the culture medium, it is known that SO2 forms sul- furous acid (H2SO3) with water (the water solubility of SO2 is 6.4% at 35℃),7) thus producing an acidic envi-
Fig. 3. Anti-IL-4 staining of a control specimen (A) and its matched SO2-exposed turbinate tissue (B). IL-4 immunoreactivity is localized to several submucosal cells in both specimens (×200).
Fig. 2. Photomicrograph showing GM-CSF immunostaining before (A, ×200) and after (B, ×400) exposure to SO2. Immunoreactivity is present predominantly in the surface epithelial cells and also in several submucosal cells following the exposure.
AAA
A BBBB
A AA
A BBBB
ronment. However, it is not very likely that the SO2- induced acidity caused the cytokine expression since a concentration of SO2 of 0.05 ppm is not sufficient to disturb the pH, and the culture media used in this study have a buffering capacity.
The airway epithelial cells have been the focus of most studies on the mechanism of air pollution-induced airway diseases due to their spatial arrangement and pr- edominance in the airways. According to these studies, there is increasing evidence suggesting that airway ep- ithelial cells play a role in the pathogenesis of airway disease-for one, these cells have been found to express and synthesize a variety of inflammatory cytokines.2-6) Although exposure of nasal or bronchial epithelial cells to air pollutants such as NO2, O3, and diesel exhaust pa- rticles (DEP) can lead to a significant increase in the release of pro-inflammatory cytokines,2)4)5) the effects of SO2 on the expression of inflammatory cytokines in the human nasal airway have not been closely studied.
Furthermore, very little information is available on the role of SO2 in cytokine production. Knorst et al. have demonstrated that the functional impairment of alveolar macrophages following exposure to SO2 can lead to a decrease in the release of tumor necrosis factor-α and interleukin-1β.8) To our knowledge, the present study is the first to examine the effects of SO2 on the tissue expression of inflammatory cytokines in nasal turbinate mucosa. Our findings indicate that the expression of GM-CSF and IL-8, but not IL-4, increases at the tissue level six hours after an exposure of one hour to a low concentration of SO2. We did not demonstrate the time- dependent expression of these cytokines, but the fact that the increased expression of GM-CSF and IL-8 was observed at six hours after exposure corresponds closely to the results of in vitro studies reported by Devalia2) and Otoshi.6)
Although the in vitro model used in this study was limited in that naturally occurring neurohumoral effects on airway inflammation were excluded, the results of the study do demonstrate that SO2 induces airway inflam- mation by increasing the expression of proinflammatory cytokines. Our results are in agreement with several pr- evious studies demonstrating an increase in the release of IL-8 and GM-CSF after exposure to NO2, O3 and DEP.2-6) The role of IL-4 in the air pollutant-induced airway response, however, is still unclear. Several recent in vivo studies suggest that IL-4 is involved in DEP- induced IgE production,9)10) while another in vitro st- udy reports that the release of IL-4 is not stimulated by NO2.2)
It is known that nasal epithelial cells are a major so- urce of IL-8 and GM-CSF6)11) and that IL-4 is observed predominantly in the mast cells and eosinophils in the
Fig. 4. Surface epithelial expression of IL-8 for individual speci- mens (thin solid lines). The thick solid line refers to group means and standard deviations. The percent expression of IL-8 signifi- cantly increases following exposure to SO2.
Fig. 5. Percent of GM-CSF reactive surface epithelium. The ex- pression of GM-CSF is significantly higher in each exposed sp- ecimen than in its matched control specimen.
Fig. 6. Submucosal cell numbers immunoreactive with anti-IL-4 antibody. There is no difference in the number of IL-4-positive submucosal cells between exposed and control tissues.
nasal submucosa.12) In this study, we demonstrated that the expression of IL-8 (as well as GM-CSF) increases predominantly in the surface epithelium following exp- osure to SO2 and that IL-4 is confined to several subm- ucosal cells irrespective of SO2 exposure. In conclusion, these results suggest that exposure to a low concentration of SO2 increases airway inflammation, possibly by ind- ucing increased expression of GM-CSF and IL-8.
REFERENCES
1) Devalia JL, Bayram H, Rusznak C, Calderon M, Sapsford RJ, Addelaziz MA, et al. Mechanisms of pollution-induced airway di- sease: In vitro studies in the upper and lower airways. Allergy 1997;52 (38 suppl):45-51.
2) Devalia JL, Campbell AM, Sapsford RJ, Rusznak C, Quint D, Godadrd P, et al. Effect of nitrogen dioxide on synthesis of infl- ammatory cytokines expressed by human bronchial epithelial cells in vitro. Am J Respir Cell Mol Biol 1993;9:271-8.
3) Devalia JL, Rusznak C, Herdman MJ, Trigg CJ, Tarraf H, Davies RJ. Effect of nitrogen dioxide and sulfur dioxide on airway res- ponse of mild asthmatic patients to allergen inhalation. Lancet 1994;344:1668-71.
4) Rusznak C, Devalia JL, Sapsford RJ, Davies RJ. Ozone-induced mediator release from human bronchial epithelial cells in vitro and the influence of nedocromil sodium. Eur Respir J 1996;9:2298-305.
5) Bayram H, Devalia JL, Sapsford RJ, Ohtoshi T, Miyabara Y, Sa- gai M, et al. The effect of diesel exhaust particles on cell function and release of inflammatory mediators from human bronchial epi- thelial cells in vitro. Am J Respir Cell Mol Biol 1998;18:441-8.
6) Ohtoshi T, Takizawa H, Okazaki H, Kawasaki S, Takeuchi N, Ohta K, et al. Diesel exhaust particles stimulate human airway ep- ithelial cells to produce cytokines relevant to airway inflammation in vitro. J Allergy Clin Immunol 1998;101:778-85.
7) Budavari, ed. The Merck index: An encyclopedia of chemicals, drugs, and biologicals. 12th ed. Whitehouse Station, N.J.: Merck &
Co;1996.
8) Knorst MM, Kienast K, Muller-Quernheim J, Ferlinz R. Effect of sulfur dioxide on cytokine production of human alveolar macrop- hages in vitro. Arch Environ Health 1996;51:150-6.
9) Kennedy JS, Baker C, Welch MR, Altman LC. Synthesis of inter- leukin-1 alpha, interleukin-6, and interleukin-8 by cultured human nasal epithelial cells. J Allergy Clin Immunol 1994;93:1060-7.
10) Diaz-Sanchez D, Tsien A, Casillas A, Dotson AR, Saxon A. Enh- anced nasal cytokine production in human beings after in vivo ch- allenge with diesel exhaust particles. J Allergy Clin Immunol 1996;98:114-23.
11) Takano H, Yoshikawa T, Ichinose T, Miyabara Y, Imaoka K, Sa- gai M. Diesel exhaust particles enhance antigen-induced airway inflammation and local cytokine expression in mice. Am J Respir Crit Care Med 1997;156:36-42.
12) Bradding P, Feather IH, Wilson S, Bardin PG, Heusser CH, Ho- lgate ST, et al. Immunolocalization of cytokines in the nasal mu- cosa of normal and perennial rhinitic subjects. J Immunol 1993;
151:3853-65.