- 118 -
Effects of βββ-toxin of Staphylococcus aureus and Neuraminidase β of Streptococcus pneumoniae on Ciliary Activity of Nasal
Ciliated Epithelial Cells*
Yang-Gi Min, M.D.1, Chung Seop Kim, M.D.2, Chae-Seo Rhee, M.D.1, In Ho Jung, M.D.3, Seok-Won Park, M.D.1 and Tae-Young Kwon, M.D.1 ABSTRACT
Background and Objectives:The in vitro effects of pneumococcal neuraminidase and staphylococcal β-toxin on ciliary activity were investigated at different concentrations and lengths of exposure. Materials and Methods:The ciliated epithelial cells were taken from the maxillary sinus mucosa of rabbits. Ciliary beat frequency (CBF) was measured at concentrations of 0.01, 0.1 and 1.0 U/mL of neuraminidase and 0.1, 1.0, 2.0, 5.0 and 10 U/mL of β-toxin using a video-computerized analysis technique. The CBF was measured 2, 4, 6, 12, 24 and 48 hours after administration of the neuraminidase and β-toxin. In the control group, normal saline was percutaneously applied to the right maxillary sinus. In the experimental group, 2 U/mL of β- toxin was applied to the left maxillary sinus using the same technique. At 7 days, all of the mucosae were taken from the infer- omedial wall of the maxillary sinus for light microscopy. Results:There was no change in CBF during a 48-hour incubation at 0.01, 0.1 and 1.0 U/mL of neuraminidase. However, the CBF dropped significantly after an 8-hour incubation at 2.0 U/mL of β-toxin (p<0.05, repeated measures ANOVA). No ciliary activity was observed after a 12-hour incubation at 10 U/mL of β- toxin. The mucoid, purulent discharge was observed in the maxillary sinuses of the experimental group. Prominent epithelial disruption and infiltration of inflammatory cells into the epithelium and lamina propria were observed in the β-toxin-applied group. Conclusion:The results of this study suggest that staphylococcal β-toxin may reduce ciliary activity and induce sinusitis without occlusion of the natural ostium of the maxillary sinus in rabbits. This study provides another animal model of sinusitis for understanding the pathogenesis of sinusitis caused by bacterial exotoxins.
KEY WORDS:Neuraminidase·β-toxin·Ciliary beat frequency·Sinusitis.
INTRODUCTION
S. pneumoniae forms colonization in the upper airway in 70% of healthy adults and children and develops air- way diseases such as otitis media, paranasal sinusitis and bronchitis. Neuraminidase is a single chain polypeptide
of 65,000 molecular weight and has the function of separating N-acetylneuramic acid of terminal from ad- jacent glucose by affecting the glycoprotein and glycoli- pid of the cell membrane. There has been no report on the enzyme that influences the invasion of S. pneumon- iae1) and its effects on the ciliated epithelial cells of the air-way.
S. aureus produces toxins, as α-, β-, γ-, and δ-to- xin and leucocidin, that injure the cell membrane and also produces materials such as protein A, clumping factor, staphylocoagulase, staphylokinase, and gelatinolytic ac- tivity, which affect cells indirectly. β-toxin (beta-he- molysin, sphingomyelinase C) of S. aureus is a toxin that causes hot-cold lysis and has a molecular weight ranging from 29,000 to 30,000. It shows substrate sp- ecificity to sphingomyelin and lysophosphatides. It is activated by Mg2+ and degrades sphingomyelin into ceramide (N-acylsphingosine) and phosphorylchline.
*This work was supported by grant No. 981-0710-083-1 from the Basic Research program of the KOSEF.
1Department of Otorhinolaryngology-Head and Neck Surgery, Seoul National University, College of Medicine, seoul, 2Otor- hinolaryngology, Seoul Red Cross Hospital, Seoul, 3Otorhinol- aryngology, Dong-A University, College of Medicine, Pusan, Korea
Address correspondence and reprint requests to Yang-Gi Min, MD, Department of Otorhinolaryngology-Head and Neck Sur- gery, Seoul National University, College of Medicine, 28 Yon- gon-Dong, Chongro-Gu, Seoul 110-744, Korea
Tel:82-2-760-2446, Fax:82-2-745-2387 E-mail:[email protected],kr
Accepted for publication on June 22, 1999
Sphingomyelin exists mostly in the outer leaflet of the lipid bilayer of the cell membrane and occupies 5-15%
of the total phospholipid, although this differs greatly de- pending on the species and type of cell.2) There is a co- rrelation between toxin sensitivity and the content of the sphingomyelin. Studies on β-toxin have been mainly li- mited to red blood cells and few studies on its influence on other cells have been made.
The bacteria that cause paranasal sinusitis also injure epithelial cells and impair ciliary motility, in some cases even causing ciliostasis by producing material that dec- reases ciliary function.3)4) The impaired ciliary function leads to increased inflammation. Depending on type, toxins may decrease ciliary function without causing hi- stological damage and at the same time injure the cell membrane or decrease ciliary motility by causing dam- age to the dynein arm in cilia.5) Studies on human nasal mucosal cells in vitro show that Hemophilus influenzae and Pseudomonas aeruginosa decrease ciliary motility.6) At times, however, ciliary function that is impaired by bacterial endotoxins recovers normally after an injection of exogeneous ATP.7) Pneumolysin of S. pneumoniae, the most common pathogen in acute paranasal sinusitis, also decreases ciliary motility without damaging the ep- ithelial cells.8) When pneumolysin-sufficient variants and pneumolysin-deficient variants are placed into a cu- lture medium containing human ciliated epithelial cells, however, ciliary activity in both variants decrease, which suggests that a material other than pneumolysin is resp- onsible for the decrease in ciliary motility.9)
Ciliary beat frequency (CBF) measured in chronic paranasal sinusitis patients vary across studies, with some researchers reporting a decrease in CBF10) and others indicating normal findings.11) Normal CBF may be due to the fact that CBF was measured after removal of the mucus containing bacterial toxins.
We attempted to determine whether pneumococcal ne- uraminidase and staphylococcal β-toxin cause a decr- ease in CBF or ciliostasis in the ciliated epithelial cells of the paranasal sinus. We also attempted to determine at which concentration the change in ciliary motility occurs.
By inducing paranasal sinusitis by an exotoxin without artificial natural ostium occlusion or bacterial inoculat- ion, we attempted to define the role of exotoxins in the pathogenesis of paranasal sinusitis and the pathophysi- ology of impaired ciliary activity.
MATERIALS AND METHODS
Preparation and culture of ciliated epithelial cells
Healthy rabbits (New Zealand white rabbits) with bo- dy weights ranging between 1.6 kg and 3.0 kg were used as experimental animals. As a first step, an intramuscular injection was made with a mixture consisting of ketam- ine (30 mg/kg) and 2% xylazine hydrochloride (30 mg/
kg), and under a status of maintained self-respiration, general anesthesia and fixation were conducted. The hairs of the nasal dorsum were removed to sterilize with 1% povidone iodine. Following this, the anterior wall of the maxillary sinus was exposed through a 1-inch incision from the skin to the periosteum. Using an elec- tric drill and a rounger under the operating microscope, the anterior wall of the maxillary sinus was removed to expose the maxillary sinus. After confirming that there was no pus, the mucosa on the medial side of the max- illary sinus was carefully dissected and obtained. The mucosa was washed with normal saline, cut into 2×2 mm2 segments and stabilized for 3 hours in a 37℃, 5%
CO2 incubator containing DMEM-F12 culture medium (Dulbecco’s Modified Eagel Madium-Han’s nutrient F12;Gibco BRL, Grand Island, NY) consisting of an- tibiotics and antifungal agents (10 IU/mL penicillin, 10 ug/mL streptomycin, 0.025 ug/mL amphotericin B) to eradicate normal flora.
Administration of neuraminidase to cultural medium containing maxillary sinus mucosa
S. pneumoniae’s neuraminidase (Sigma, St. Louis, MO) was used as the toxin after dilution with DM-EM-F12 culture medium to dilution degrees of 0.02, 0.2 and 2.0 U/mL and final concentrations of 0.01, 0.1 and 1.0 U/
mL at the time of placement into the culture medium co- ntaining the epithelial cells. The culture medium was set in a 37℃, 5% CO2 incubator and the actual pH of the culture medium was measured with a pH measurer. The pH of the diluted culture medium with toxin read 7.5.
One mL DMEM/F12 culture medium was placed in the prepared culture dish and the stabilized mucosa was ad- hered to the bottom of the culture dish with platinum line. Ciliary beat was viewed using inverted microscope
at 1,000× magnification. Only the culture dishes sho- wing active ciliary activity, as observed under the inv- erted microscope, were used for the experiment. Five culture dishes were used for the experimental group and one was used for the control group at each β-toxin co- ncentration. In the experimental group, 1 mL diluted β- toxin was mixed to make final concentrations of 0.01, 0.1 and 1.0 U/mL, while in the control group, 1 mL DM- EM/F12 culture medium containing no toxin was mixed to produce a total volume of 2 mL of culture medium in each culture dish.
Administration of β-toxin to culture medium containing maxillary sinus mucosa
The β-toxin of S. aureus (Sigma, St. Louis, MO) was used as the toxin after dilution with DMEM-F12 culture medium to degrees of 0.2, 2.0, 4.0, 10 and 20 U/mL and final concentrations of 0.1, 1.0, 2.0, 5 and 10 U/mL at the time of placement into the culture medium containing the epithelial cells. Five culture dishes were used for the experimental group and one was used for the control group at each β-toxin concentration. In the ex- perimental group, the above mentioned 1 mL diluted β-toxin was mixed to form final concentrations of 0.1, 1.0, 2.0, 5 and 10 U/mL, while in the control group, 1 mL DMEM/F12 culture medium containing no toxin was mixed to produce a total volume of 2 mL of culture me- dium in each culture dish.
Measurement of ciliary beat frequency
In the experimental and control groups that were ad- ministered with neuraminidase and β-toxin, respecti- vely, CBF was measured at time intervals of 0, 2, 4, 6, 12, 24 and 48 hours. The measurements were conducted at five areas showing the most active ciliary movements in each culture dish and the average CBF was obtained.
With an inverted microscope at 1,000× magnification, a charged coupled device (CCD) camera (Digistar, Xo- med, Jacksonville, FL) was used to record the ciliary movement through a monitor in a clear view. After rec- ording for 10 seconds, the CBF was obtained with a vi- deo-computerized analysis system. The image that was used was not the entire image viewed in the monitor but only the area where the cilia moved actively. This was possible because the multiangled windows of the anal-
ysis system allows selection of only the desired images.
A video-computerized analysis system converts the light coming through an inverted microscope to analog sign- als of the National Television System Committee (NTSC) type with a CCD camera. The analog signals are then converted to digital data with the image graver system of a computer, and the CBF can be obtained through an an- alysis program based on Fast-Fourier Transform (FFT) technology.8) Because ciliary motility is easily influenced by temperature, a heating chamber and a temperature re- gulator were used to maintain the temperature of the culture medium at 37℃.
Induction of Experimental Sinusitis by β-to- xin of S. aureus
The β-toxin obtained from S. aureus was used since it was diluted with DMEM/F12 to arrive at the minimum concentration of 2 U/mL at which the CBF decreased significantly compared to the control group. The nasal dorsum skin of the 10 rabbits was obtained to conduct puncture and aspiration on the anterior walls of the ma- xillary sinus. After confirming that there was no pus, 1 mL normal saline was percutaneously applied to the left maxillary sinuses, which were then classified as the co- ntrol group. The β-toxin diluted at a concentration of 2 U/mL was applied to the right maxillary sinuses, wh- ich were defined as the experimental group. Following this, the nasal dorsum of the skin was sutured.
After one week of percutaneous application of β- toxin to the maxillary sinus, an intramuscular injection of mixture containing ketamine (30 mg/kg) and 2% xy- lazine hydrochloride (30 mg/kg) was performed on the rabbits. The hairs of the nasal dorsum were removed and sterilized with 1% povidone iodine, and a 1-inch incision was made from the skin to the periosteum, thus exposing the anterior wall of the maxillary sinus. Using an electric drill and a rounger under the operating microscope, the anterior wall of the maxillary sinus was removed to ex- pose the maxillary sinus. Following this, the mucosa on medial side of the maxillary sinus was carefully dissec- ted and obtained. The obtained tissue was placed for 24 hours in a 10% formalin solution and was paraffinem- bedded for thin-section at a thickness of 4 μm. Dehy- dration was performed with alcohol at gradually lower concentrations and hematoxylin-eosin stain was condu- cted. The maxillary sinus mucosa was then observed for
ciliary loss, infiltration of the inflammatory cells and epithelial ulceration. According to degree as based on these three findings, the lesions were classified into one of three categories-mild, moderate or severe. The lesion was defined as ‘mild’ if ciliary loss and epithelial ulcer was observed in 5% or less of the specimen. When found in 5-20% of the specimen, the lesion was classified as
‘moderate,’ and when in more than 20% of the speci- men, ‘severe.’ Infiltration was defined as ‘severe’ if most of the epithelial layer and the subepithelial layer were infiltrated with inflammatory cells, ‘mild’ if the infiltration was limited to the subepithelial layer, ‘mod- erate’ if the findings were in mid-range.
Statistical analysis
The changes in CBF caused by neuraminidase and β-toxin at different concentrations were statistically analyzed using the SAS system for windows (version 6.11). To check for differences between the control gr- oup and the experimental group over time, and to dete- rmine whether each measuring value in the experimental group changed, p-values were obtained using repeated measures analysis of variance (ANOVA). To check for differences between the control group and the experi- mental group at each experimental time point, p-values were obtained using the Wilcoxon rank sum test with a statistical significance of 0.05.
RESULTS
Influence of neuraminidase and β-toxin on CBF of the ciliated epithelial cells in rabbits
The CBF measured at 0.01 U/mL, 0.1 U/mL and 1.0 U/mL neuraminidase showed no statistically significant difference in CBF between the control groups (repeated measure ANOVA and Wilcoxon rank sum test, p>0.05).
The CBF also showed no significant time-based differ- ence (Fig. 1).
The CBF measured under 0.1 U/mL and 1.0 U/mL β-toxin showed no statistically significant difference between the control and experimental groups (repeated measure ANOVA and Wilcoxon rank sum test, p>0.05).
The CBF also showed no significant time-based differ- ence. The CBF measured at 2.0 U/mL β-toxin showed no statistically significant difference (repeated measure
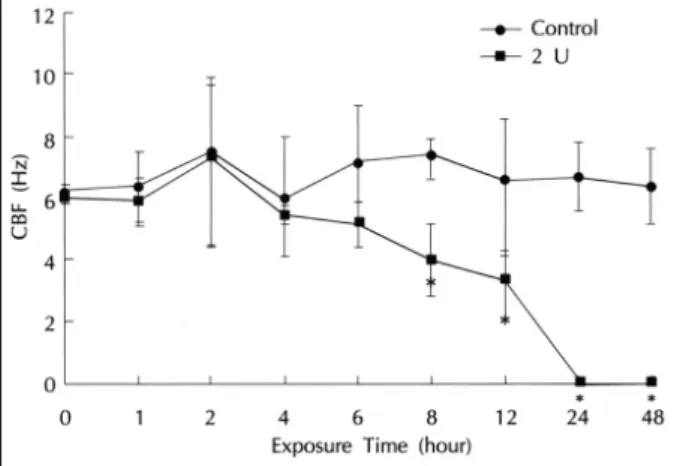
ANOVA and Wilcoxon rank sum test, p>0.05). There was no significant change in CBF until six hours after the measurement but the CBF showed a statistically si- gnificant decrease in the experimental group compared to the control group from the 8th hour. At the 24th hour, the ciliary motility of the experimental group ceased (Fig. 2).
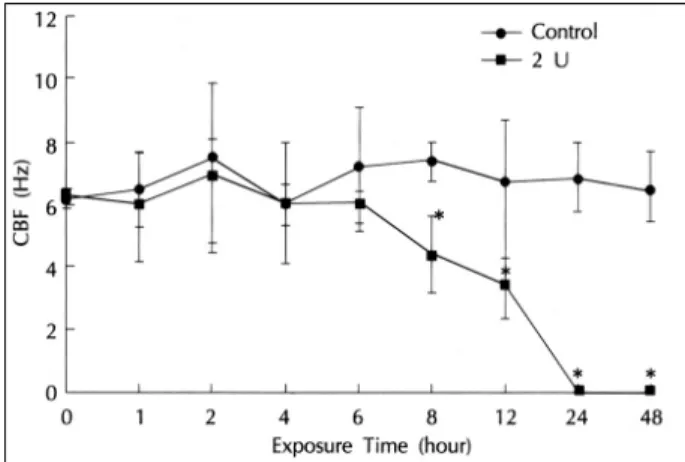
The CBF measured at 5.0 U/mL β-toxin showed no statistically significant difference between the control and experimental groups (repeated measure ANOVA and Wilcoxon rank sum test, p>0.05). There was no signif- icant change of CBF until 6 hours after the measurement but the CBF showed a statistically significant decrease in the experimental group compared to the control group from the 8th hour. At the 24th hour, the ciliary motility of
Fig. 2. Effects of 2.0 U/mL staphylococcal β-toxin on CBF of the ciliated epithelial cells of the maxillary sinus as a function of exposure time in rabbits. CBF is significantly different between the two groups. CBF significantly changes as a function of the time in the toxin-exposed group. *Significantly different from co- ntrol.
Fig. 1. Effects of 0.01, 0.1 and 1 U/mL pneumococcal neurami- nidase on DBF of the ciliated epithelial cells of the maxillary sinus as a function of exposure time in rabbits. CBF is not signifi- cantly different between all groups. CBF does not significantly change as a function of the time in the toxin-exposed groups.
the experimental group ceased (Fig. 3).
The CBF measured at 10 U/mL β-toxin showed no statistically significant difference compared with the co- ntrol group (repeated measure ANOVA and Wilcoxon rank sum test, p>0.05). There was no significant change of CBF until 6 hours after the measurement but the CBF showed a statistically significant decrease in the experimental group compared to the control group from the 8th hour. At the 12th hour, the ciliary motility of the experimental group ceased (Fig. 4).
Histopathologic findings of maxillary sinusitis induced by Staphylococcus aureus β-toxin Gross findings
One of the experimental animals died one week after the experiment. A gross observation of the secretion fr- om the maxillary sinus revealed a whitish purulent dis- charge in the maxillary sinuses of all 9 surviving rabbits in the experimental group. Meanwhile, in the maxillary sinuses of the control group formation of the whitish purulent discharge was not found.
Light microscopic findings
In the control group’s maxillary sinus mucosa speci- mens that were percutaneously administered with normal saline, epithelial cells with well-developed brush borders formed the epithelium in one layer. In the subcutaneous layer, there were a small number of serous glands and mucinous glands and there were an abundance of capi- llaries. A few lymphocytes and monocytes were found.
In the experimental group, there were in all cases signs of ciliary loss, infiltration of inflammatory cells and ep- ithelial ulcer findings of above ‘moderate’ degree (Fig.
5A, B).
DISCUSSION
There have been a number of studies on the working mechanism of S. pneumoniae on ciliated epithelial cells.
Cocchiara et al.12) reported that a substance P-induced histamine release is inhibited at 98% after a pretreatm- ent of 0.1 U/mL concentration neuraminidase on rats’
monocytes. Winter et al.13) reported that the neuramin-
idase does not appear to damage the cochlea in guinea pigs. Scanlon 14) reported that a neuraminidase concen- tration may reach 200 U/mL in a child’s middle ear eff- usion. In the present study, changes in CBF did not vary from those in the control group when the sinus ciliated epithelial cell was exposed to 0.01 U/mL, 0.1 U/mL and 1.0 U/mL neuraminidase.
The β-toxin is known to cause hemolysis by dam- aging the cell membrane, especially the red blood cell membrane, through dissolving sphingomyelin, a phos- pholipid of the cell membrane, into ceramide (N-acy- lsphingosine) and phosphocholine. Bramley et al.15) report finding that β-toxin had a higher correlation with intramammary infection than did mutations of S. aureus
Fig. 3. Effects of 5.0 U/mL staphylococcal β-toxin on CBF of the ciliated epithelial cells of the maxillary sinus as a function of exposure time in rabbits. CBF is significantly different between the two groups. CBF significantly changes as a function of the time in the toxin-exposed group. *Significantly different from corol.
Fig. 4. Effects of 10.0 U/mL staphylococcal β-toxin of CBF of the ciliated epithelial cells of the maxillary sinus as a function of exposure time in rabbits. CBF is significantly different between the two groups. CBF significantly changes as a function of the time in the toxin-exposed group. *Significantly different from co- ntrol.
that failed to express β-toxin mouse in the mammary gland. And according to O’Callaghan et al.,16) a scleral edema results from an application of β-toxin on a ra- bbit’s eye because the content of the sphingomyelin in the scleral epithelial cell membrane is high. There are, however, other reports arguing that the working mecha- nism of β-toxin cannot be related to the content of the sphingomyelin. Wilkinson17) reports finding that β-to- xin decreased the movement of human monocyte but did not have any influence on the granulocyte. In the cell membranes of human granulocyte and monocyte, the sphingomyelin content ratio of the total membrane lipid was found to be 20.8% and 10.9%, respectively. Alth- ough the sphingomyelin content in the monocyte was smaller, the β-toxin affected only the monocyte to kill it.18) Waley et al.19) cultured human monocyte in a cult- ure medium containing 0.1 U/mL β-toxin and observed the loss of ATP in the monocyte. According to Waley et al., the monocyte died because the β-toxin caused small pores on the cell membrane of the monocyte, re- sulting in irreversible depletion of the ATP.
Some recent studies report on the occurrence of mo- dulation of cell differentiation, cell cycle progression and tissue inflammation when ceramide forms as a result of dissolved sphingomyelin serving as a ‘second messen- ger.’20) In the present study, evaluating the changes that occurred after exposure to β-toxin showed no change in CBF when the concentration was below 1.0 U/mL.
In concentrations of above 2.0 U/mL, there was no dif- ference in the changes in CBF compared to the control group during the first 6 hours, but the CBF dropped co- nsiderably from the 8th hour, ceasing after 24 hours.
Based on these findings, there appears to be three poss- ible reasons for the decrease. The first theory is based on the assumption that the β-toxin affects the sphingo- myelin of rabbits’ ciliated epithelial cells to cause injury to the cell membrane. Secondly, there is the possibility that the β-toxin forms pores in the ciliated epithelial cells at a certain concentration, thereby inducing the loss of ATP and the decrease in ciliary activity. In the present study, there was no time-based difference in the change in CBF at a β-toxin concentration of below 1 U/mL
Fig. 5. Light micrographs in 2 U/mL β-toxin-applied group. A:
Moderate cliary loss with prominent epithelial disruption and the infiltration of the inflammatory cells into the epithelium and lami- na propria were observed (H & E ×400). B:Severe exfoliation and the infiltration of the inflammatory cells into the epithelium and lamina propria were observed (H & E ×200).
A A AA
BbBbBb Bb BaBa
BaBa
compared with the control group, possibly because a co- ncentration below 1 U/mL was not sufficient to create pores on the ciliated epithelial cells, while concentrations of above 2 U/mL resulted in pores being formed on the cell membrane. The volume and size of the pores incre- ased in accordance with higher concentrations, and the CBF was found to decrease as the rate of ATP loss rose.
Thirdly, there is the possibility that ceramide, the disso- lved material of sphingomyelin, induces a inflammatory reaction on tissue. However, there has been no study on the sphingomyelin contents of cell membranes in rab- bits’ maxillary sinus mucosa, and there is the need for further study to determine the size of the pores β-toxin creates at each concentration.
With β-toxin decreasing the CBF of respiratory ep- ithelial cells at concentrations of more than 2.0 U/mL in the in vitro experiment, maxillary sinusitis was indu- ced by the lowest possible concentration, 2.0 U/mL. In a study by Lee et al.,21) following the infusion of S. pn- eumoniae into a group of rabbits under a status of natu- ral ostium occlusion, the occurrence of maxillary sinusitis was 100%, while in a group with just occlusion of the natural ostium and not infused with the bacteria, the rate of occurrence was 60%. The rate of occurrence fell to 10% in a group infused with bacteria under a status of opened natural ostium. Based on these findings, occ- lusion of the natural ostium appears to be more impor- tant than whether there is an infusion of S. pneumoniae in the pathogenesis of the maxillary sinusitis. Addition- ally, the possibility of sinusitis occurrence is considered to be low when there is only bacterial infusion under a status of opened natural ostium. This can be explained by the fact that the bacteria invading the paranasal sin- us are quickly expelled to the nasopharynx through the natural ostium by ciliary activity when the mucociliary transport is normal and the natural ostium is opened.
This provides only a short period of time for the bacteria to make contact with the mucosa and makes invasion into mucosa impossible, ultimately preventing an inflamm- atory reaction. But unilateral paranasal sinusitis is able to spread into the contralateral paranasal sinus of the pa- ranasal sinus whose natural ostium is opened, or paran- asal sinusitis may be induced in a paranasal sinus whose natural ostium is normal by inserting a foreign body. Fu- rthermore, odontogenic paranasal sinusitis and allergic paranasal sinusitis may develop even in cases where the maxillary sinus natural ostium is normal.22) Therefore,
it is difficult to clarify the etiology of the paranasal si- nusitis with only natural ostium occlusion and bacterial infusion. In the present study, maxillary sinusitis was induced only by percutaneous application of β-toxin without the natural ostium occlusion. The molecular we- ight of β-toxin is believed to range between 29,000 and 30,000, much lower than that of bacteria, allowing it to stay longer in the maxillary sinus and decrease CBF, which in turn reduces mucociliary transport and induces maxillary sinusitis.
Considering these results, ciliary motility decreased or stopped depending on the concentration of the toxin or the length of contact when mucosal epithelial cells of the nasal cavity or paranasal sinus are exposed to β-toxin.
Induction of the experimental maxillary sinusitis was po- ssible by β-toxin only without natural ostium occlusion or bacterial inoculation. Accordingly, the treatment of irreversible pathologic changes in the paranasal sinus mu- cosa may require the development and application of an antitoxin of β-toxin for paranasal sinusitis.
REFERENCES
1) Willett HP. Staphylococcus & Streptococcus pneumoniae. In: Jo- klik WK, Willett HP, Amos DB, Wilfert CM, eds. Zinsser Micr- obiology. 19th ed. East Norwalk: Prentice-Hall International Inc;
1988.
2) Zierdt CH. Long-term Staphylococcus aureus carrier state in hos- pital patients. J Clin Microbiol 1982;16:517-20.
3) Wilson R, Cole PJ. The effect of bacterial products on ciliary fun- ction. American Review of Respiratory Diseases 1988;138:49-53.
4) Harada T, Saida S, Majima Y, Ukai K, Sakakura Y. Effect of lipo- polysaccharide of Hemophilus influenzae on ciliary activity of the human nasal mucosa. Acta Otolaryngol (Stockh) 1987;103:307-11.
5) Hingley ST, Hastie AT, Kueppers F, Higginns ML, Weinbaum G, Shryock T. Effect of ciliostatic factors from Pseudomonas aerugi- nosa on rabbit respiratory cilia. Infect Immun 1986;51:254-62.
6) Wilson R, Roberts D, Cole P. Effect of bacterial products on hu- man ciliary function in vitro. Thorax 1985;40:125-31.
7) Rautiaimen M, Yoshitsugu M, Matsune S, Nuutinen J, Happonen P, Ohyama M. Effect of exogenous ATP and physical stimulation on ciliary function impaired by bacterial endotoxin. Acta Otolar- yngol (Stockh) 1994;114:337-40.
8) Steinfort C, Wilson R, Mitchell T, Feldman C, Rutman A, Todd H, et al. Effect of Streptococcus pneumoniae on human respiratory epithelium in vitro. Infect Immun 1989;57:2006-13.
9) Rayner CFJ, Jackson AD, Rutman A, Dewar A, Mitchell TJ, An- drew PW, et al. Interaction of pneumolysin-sufficient and –def- icient isogenic variants of Streptococcus pneumoniae with human respiratory mucosa. Infect Immun 1995;63:442-7.
10) Settipane GA. Nasal polyps and immunoglobulin E (IgE). Allergy Asthma Proc 1996;17:269-73.
11) Saano V, Nuutinen J, Virta P, Joki S, Karttunen P, Silvasti M. The effect of ATP on the ciliary activity of normal and pathological hu- man respiratory mucosa in vitro. Acta Otolaryngol (Stockh) 1991;
111:130-4.
12) Roberta Cocchiara, Antonella Bongiovanni, Giuseppe Albeggiani, Antonina Azzolina, et al. Inbibitory effect of neuraminidase on SP-induced histamine release and TNF-amRNA in rat mast cells:
evidence of a receptor-independent mechanism. Journal of Neur- oimmunology 1997;75:9-18.
13) Andrew J Winter, Spiro D Comis, Michael P Osborne, Michael J Tarlow, et al. A role of Pneumolysin but not Neuraminidase in the Hearing Loss and Cochlear Damage Induced by Experimental Pn- eumococcal Meningitis in Guinea Pigs. Infection and Immunity 1997;65:4411-8.
14) Lenia Scanlon K, Warren F Diven, Robert H Glew. Purification and properties of Streptococcus pneumoniae Neuraminidase. Enzyme 1989;41:143-50.
15) Bramley AJ, Patel AH, Reilly MO, Foster R, Foster TJ. Roles of Alpha-Toxin and Beta-Toxin in Virulence of Staphylococcus aure- us for the Mouse Mammary Gland. Infection and Immunity 1989;
57:2489-94.
16) Richard JO Callaghan, Michelle C Callegan, Judy M Moreau, Linda C Green, Timothy J Foster, et al. Specific Roles of Alpha- Toxin and Beta-Toxin during Staphylococcus aureus Corneal in-
fection. Infection and Immunity 1997;65:1571-8.
17) Wikinson PC. Inhibition of Leukocyte locomotion and chemotaxis by lipid-specific bacterial toxins. Nature 1975;255:485-91.
18) Kennett FF, Schenkein HA, Ellis TM, Rutherfold RB. Phospholi- pid composition of human monocytes and alterations occurring due to culture and stimulation by C3b. Biochim Biophys Acta 1984;
804:301-7.
19) Iwan Walev, Ulrich Weller, Susanne Strauch, Timothy Foster, Su- charit Bhkdi. Selective Killing of Human Monocytes and Cytokine Release Provoked by Sphingomyelinase (Beta-Toxin) of Staphyl- ococcus aureus Infection and Immunity 1996;64:2974-9.
20) Hannun Y. The sphingomyelin cycle and second messenger func- tion of ceramide. J Biol Chem 1994;269:3125-8.
21) Lee BJ, Kim SY, Min YG. An experimental study on the pathog- enesis and histopathological changes of acute maxillary sinusitis.
Korean J Otolaryngol 1991;34:489-504.
22) Kumlien J, Schiratzki H. Blood flow in rabbit sinus during expe-rimentally induced
chronic sinusitis. Acta Otolaryngol 1985;99: 630-6.