- 14 -
The Role of Epithelium in the Pathophysiology of Chronic Rhinosinusitis : An Update
Jin-Young Min, MD, PhD1 and Hun-Jong Dhong, MD, PhD2
1Department of Otolaryngology-Head and Neck Surgery, Northwestern University Feinberg School of Medicine, Chicago, Illinois, USA,
2Department of Otorhinolaryngology-Head and Neck Surgery, Sungkyunkwan University School of Medicine, Samsung Medical Center, Seoul, Korea
ABSTRACT
Chronic rhinosinusitis (CRS) is characterized by persistent symptomatic inflammation of the nasal and parana- sal sinus mucosa resulting from the interaction of multiple host and environmental factors. Although the etiology of CRS remains a matter of vigorous debate, research into CRS has implicated that the epithelium plays an im- portant role as both a mediator and a regulator of innate and adaptive immune responses, as well as the transition from innate immunity to adaptive immunity. Recently, alterations of epithelial barrier functions, dysregulation of the innate immune system and inappropriate activation of the adaptive immune system have been identified as active processes in the pathogenesis of CRS. This review summarizes recent emerging evidence regarding the un- derlying pathogenic mechanism in CRS, which highlights the increasing recognition of the epithelium as a central factor in CRS etiology and pathogenesis.
KEY WORDS : Rhinosinusitis · Epithelium · Inflammation · Innate Immunity · Adaptive Immunity.
INTRODUCTION
Chronic rhinosinusitis (CRS) is a clinical syndrome as- sociated with persistent inflammation of the mucosa of the nose and paranasal sinuses resulting from the interaction of multiple host and environmental factors. There are 2 most common variants of this disease including chronic rhinosinusitis with nasal polyps (CRSwNP) and chronic rhinosinusitis without nasal polyps (CRSsNP), which have clinically and morphologically different characteristics.
The etiology and pathogenesis of CRS remain contro- versial; however, specific proposed mechanisms for persis- tent inflammation including obstruction of the osteomeatal complex, impaired mucociliary clearance, osteitis, atopy and microbial resistance have all been theorized to play a role.1) Recently, research into the workings of the role of epithelium in the aspect of immunity is emphasized. The concept of a host immune dysfunction forms the basis of
one line of current research into CRS pathogenesis. The upper airways and sinuses play an important filtering role.
Under optimal conditions, the epithelium of upper airways and sinuses serves as the site of interface with inhaled ir- ritants and pathogens without involvement of the adaptive immune system. If the barrier function of the epithelial layer fails, various materials may gain access to cells of the immune response in and below the lamina propria, further stimulating the inflammatory process and acute or chronic inflammation can result. Mucociliary clearance, physical exclusion, and the innate and adaptive immune responses are used to separate host from environment.
This article briefly summarizes recent insights to iden- tify emerging evidences underlying pathogenic mecha- nism in CRS highlighting the increasing recognition of epithelial host defects as central factors in CRS etiology and pathogenesis.
The use of epithelium to study pathophysiology of chronic rhinosinusitis
As surgical intervention is frequently necessary to man- age the disease of a significant proportion of people suf- fering from CRS, investigators have access to the substan- tial quantities of nasal tissues of CRS patients including Address correspondence and reprint requests to Hun-Jong Dhong, MD,
PhD, Department of Sungkyunkwan University School of Medicine, 50 Ilwon-dong Gangnam-Gu, Seoul 135-710, South Korea
Tel: +82-2-3410-3573, 3579 , Fax: +82-2-3410-3879 E-mail: [email protected]
Received for publication on April 25, 2013 Accepted for publicatoin on May 9, 2012
sinus mucosa, turbinate mucosa, and nasal polyps that are removed during the surgical procedures. In addition, the good primary source of normal control tissue is from patients undergoing transnasal resection of intracranial lesions. To study the immune and inflammatory process in CRS, it is also helpful to collect epithelial cell scrap- ings and nasal lavage samples. DNA, mDNA, and protein extracts of the tissues can be obtained from the samples.
Protein extracts and nasal lavage samples are useful to evaluate the production of cytokines, chemokines, host defense molecules, and immunoglobulins (Ig). Messenger RNA could be converted into cDNA for analysis of gene expression by real-time RT-PCR, and cells are cultured for comparative studies of responses in vitro. In addition, tis- sues can be used to perform immunohistochemistry.
Alterations of epithelial barrier function in chron- ic rhinosinusitis
In CRS, evidences for alterations of epithelial barrier function include significantly diminished tight junction proteins (Zonula Occludens-1, ZO-1) and increased ion permeability.2) 3) More recently, other study demonstrated that serine peptidase inhibitor Kazal type 5 (SPINK5), which is one of many proteins involved in barrier main- tenance and encodes the protein Lympho-epithelial Ka- zaltype-related inhibitor (LEKT1), a protease inhibitor involved in maintaining epithelial barrier function, is sig- nificantly decreased in CRS.4) Various pathogens includ- ing dust mites, fungi and bacteria have significant intrinsic protease activity.5) Lower levels of protease inhibitors in CRS epithelium may result in increased susceptibility to endogenous and exogenous protease activity.6) 7)
In addition to mechanical barrier maintenance, protease inhibitors protect protease-activated receptors (PARs) from excessive stimulation by exogenous proteases. This excessive PAR stimulation could at least trigger increased cytokine and chemokine production, cellular recruitment and subsequent adaptive immune response.
Alterations in epithelial barrier function may, in turn, allow greater mucosal penetration of foreign materials and subsequent magnification of the inflammatory response through stimulation.7-9)
Dysregulation of innate immune system in chron- ic rhinosinusitis
The innate immune system refers to inborn resistance that is present before the first exposure to the antigen. The innate immune system is composed of cells and their as- sociated mechanisms which defense against pathogens through genetically encoded pathways with limited speci-
ficity for molecular structures.
Sinonasal epithelial cells express pattern-recognition receptors (PRRs) that recognize pathogen-associated molecular patterns (PAMPs) found in bacteria, viruses, parasites, yeast, and mycobacteria.10-12) One of the impor- tant PRRs in CRS is the Toll-like receptor (TLR) family that, when activated, trigger a pro-inflammatory response through the activation of nuclear transcription factors such as NF-κB, AP-1 and IRF3.13) TLRs including TLR2, TLR3, TLR4, and TLR9 are expressed in the airway and they play an important role in mediating host inflamma- tion and the development of CRS.14) TLR2 plays a promi- nent role in responses to fungi and gram-positive bacteria.
TLR3 responds to viral replication products and TLR4 responds to endotoxin. TLR5 recognizes bacterial flagel- lin, and TLR7 and TLR8 are activated by single stranded RNA.15) There are some studies suggesting increased TLR expression in CRS.12) 16)
In addition to PRRs, sinonasal epithelial cells express other various antimicrobial molecules.17-19) A recent study suggested that the S100 proteins might play a significant role in mediating some forms of CRS. The S100 proteins have multiple effects on barrier function and cell differen- ciation. Several members including S100A7 (psoriasin), S100A8/S100A9 (calprotectin) act as classic antimicro- bial proteins with direct antibacterial and antifungal ef- fects, recruit neutrophils and lymphocytes, and also aid in wound healing.20) 21) It is revealed that S100A7, S100A8 and S100A9 are significantly reduced in CRS when com- pared with controls and diminished S100 proteins in si- nonasal epithelial cells may predispose to development of CRS, possibly through (1) increased susceptibility to infection by microorganisms that are directly sensitive to these proteins, (2) diminished transepithelial migration of leukocytes, and (3) reduced repair and proliferation of epithelium.4) 5) However, the hypothesis that deficiencies of expression of S100 in CRS compromises maintenance of barrier function in the upper airways and sinuses is un- known and presently under evaluation.
Recent study demonstrated that the expression levels of innate immune proteins of the palate lung and nasal epi- thelium clone (PLUNC) family were profoundly reduced in nasal polyps (NPs) compared to uncinate tissue from control subjects or patients with CRS.22) Decreased short PLUNC1 (SPLUNC1) and LPLUNC2 in NPs reflect a profound decrease in the number of submucosal glands and decreased glands may lead to a localized defect in the production and release of glandular innate defense mol- ecules.22)
An important inducer of innate immune responses is the cytokine IL-22 that is secreted by Th17 and Th1 cells and activates sinonasal epithelial cells via the IL-22 recep-
tors (IL22R). It is reported that the levels of IL22R1 were significantly decreased in CRSwNP when compared with controls, suggesting a diminished IL-22 response in CR- SwNP.23) This provides an interesting paradigm in which decreased IL-22R on sinonasal epithelial cells may impair innate immune responses in CRSwNP. Similarly, sinona- sal epithelial cells of CRS produce less IL-8 and respond less well to TLR2 ligands. Other studies focusing on IL-6, which activates Th17 cells and regulates the response of B lymphocytes and regulatory T cells, the soluble IL-6 receptor, and the soluble signaling molecule gp130 that suppresses IL-6 signaling have shown that all of these molecules are elevated in CRSwNP.24)
Because of the importance of the epithelium as a me- diator of immune defense, defects of epithelium-related genes could contribute to an innate immune dysfunction in patients with CRS. Genome-wide association studies focused on differences at the genomic level between cases and controls. Some of the mutation studies focused on single nucleotide polymorphisms (SNPs) in genes reflect- ing the absence of specific SNPs in TLR2 linked to CRS or the presence of mutations linked to the functionality of the newly discovered innate lymphoid cell (ILC)2.25-28) Inappropriate activation of adaptive immune sys- tem through epithelial-mediated mechanisms in chronic rhinosinusitis
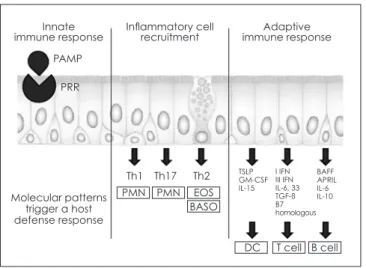
The integration of the innate and adaptive immune re- sponses in the sinonasal tract has not been extensively studied but likely begins with the recognition of PAMPs and cellular damage by multiple cell types that respond by secreting immune activating factors including cytok- ines that stimulate antigen presenting cells (APCs) and chemokines that attract the cellular components of the im- mune response (Fig.1). Damage to the epithelium likely
exposes more PRRs to PAMPs, amplifying the immune response; if the PAMP stimulus is sufficiently strong, an acquired immune response will result.
There are considerable evidences for the occurrence of epithelial cell activation in patients with CRS. Increased levels of cytokines, such as GM-CSF; chemokines, such as CCL5 (RANTES) and eotaxins; growth factor recep- tors, such as epidermal growth factor receptor; costimula- tory molecules, such as programmed death ligand 1 and 2;
and other molecules involved in inflammation and immu- nity are observed in epithelium in patients with CRS.5) Regulation of dendritic cells by epithelium
Tissue dendritic cells (DCs) are important in generation of adaptive immune response, acting as APCs gathering and processing peptides from the environment to present to T and B cells. After stimulation by PRRs through PAMP recognition, DCs become activated, cease phagocytic ac- tivity, and acquire chemokine receptors that lead them to migrate to lymph nodes where they present antigen to Th cells.29) Activation of epithelium could mobilize DCs for- mation and recruitment in the airways.
It was reported that some of the chemokines and cytok- ines induced were ones that would be expected to activate DCs, namely CCL20 (MIP-3a) and GM-CSF.10) GM-CSF is a powerful inducer of DCs formation from monocytes and CCL20 is a ligand for CCR6 and can recruit mature DCs.30) 31)
Thymic stromal lymphopoietin (TSLP), which is one of the key airway epithelial proteins can be strongly in- duced by common viruses. High levels of TSLP trigger DC-mediated polarization of the helper cell response in a Th2 direction.32) Furthermore, recent studies suggest that the absence of protease inhibitors including LEKT1 may increase TSLP production.33) This suggest the hypothesis
Fig. 1. Epithelium of the nasal and paranasal sinus mucosa plays an important role in innate and adaptive immune responses. Epithelial cells express pattern-recognition re- ceptors and recognize pathogen-associated molecular patterns. During immune responses, epithelial cells release specific chemokines that recruit the effector cells that are appropriate to the particular immune response. Finally, epi- thelial cells regulate the adaptive immune response by ex- pression of soluble and cell-surface molecules that alter the function of dendritic cells, T cells, and B cells. (Modified from Schleimer et al. J Allergy Clin Immuol. 2007;120:1279-84) PAMP, pathogen-associated molecular pattern; PRR, pat- tern-recognition receptor; PMN, polymorphonuclear leuko- cyte; EOS, eosinophil; BASO, basophil; TSLP, thymic stromal lymphopoietin; BAFF, B cell-activating factor of the TNF fam- ily; APRIL, a proliferation-inducing ligand.
Innate immune response
Molecular patterns trigger a host defense response
Th1 TSLPGM-CSF
IL-15
BAFFAPRIL IL-6IL-10 I IFN III IFN IL-6, 33 TGF-ß B7homologous
PMN
DC T cell B cell
PMN EOS
Th17 Th2 Inflammatory cell
recruitment Adaptive
immune response PAMP
PRR
BASO
that diminished LEKT1 levels in CRS may accentuate the TSLP response of epithelial cells to rhinovirus, account- ing for the heightened Th2 response characteristic of some forms of CRS.
Regulation of T cells by epithelium
Increased numbers of T cells have been reported in CRS. In addition, increased activation of the T cells that are present in nasal polyps has been reported.34) Several groups have investigated that the primary transcription factor in T-regulatory (Treg) cells, was decreased relative to controls along with transforming growth factor (TGF)-β in CRSwNP.35) 36)
Nasal polyps with a more eosinophilic infiltrate had higher levels of the Th2-specific transcription factor GATA-3, whereas the neutrophilic nasal polyps showed higher levels of Tbet, a transcription factor characteristic of Th1 cells.35)
Several recent studies have shown that airway epithelial cells express high levels of certain B7 family members, notably B7-H1 and B7-DC, which are important regula- tors of T-cell activation, function, survival, and differentia- tion.37) 38) Although B7-H1 and B7-DC levels are increased in CRS, their relevance to immune regulation and disease pathogenesis is under investigation. The mechanisms by which these various T cell populations are activated and recruited to the nasal mucosa are still controversial.
Regulation of B cells by epithelium
Sinonasal epithelial cells also appear to interact with B cells, which are critical mediators at mucosal sites and in a growing number of chronic inflammatory diseases.
Emerging research highlights a potentially important pathogenic role for B lymphocytes in the inflammation associated with CRS. B cells are a key component of the adaptive immune response. In the airway, localized differ- entiation of B cells into plasma cells and localized, lymph node independent, class switch recombination are thought to play particularly important roles in the pathogenesis of allergic diseases of the airway.39) In addition to their ability to produce antibodies that contribute to disease pathogenesis, B cells can function as antigen presenting or regulatory cells, and produce a variety of cytokines and chemokines that can influence inflammation. Recent study has investigated that B cell-activating factor of the TNF family (BAFF ; also called BLys,TNFSF13B) mRNA and protein levels are highly elevated in nasal polyp tissue in CRSwNP in comparison with CRSsNP and control tis- sues.40) BAFF is an important regulator of Ig class-switch recombination and production in B cells, independent
of T-cell help. BAFF also enhances T cell–dependent Ig class switching and production. Kato et al. reported that BAFF is produced by epithelial cells and could be in- duced by stimulation by several cytokines and TLR3.41) These findings suggest that dysregulated BAFF produc- tion by sinonasal epithelial cells may distort the adaptive immune response and play a role in polyp formation. In addition to BAFF, nasal polyps contain increased levels of the cytokine IL-6 and chemokines such as B-lymphocyte chemoattractant (CXCL13) and stromal cell–derived fac- tor 1a, which are known to play a role in B-cell recruit- ment and plasma cell differentiation.42) 43)
Several reports have demonstrated elevated levels of various isotypes of Igs including IgG, IgE and IgA in si- nus tissue from patients with CRS.44) 45) Hulse et al. re- ported that nasal polyp tissue from patients with CRS had abundant accumulation of inflammatory cells, especially B cells and plasma cells. In addition, they also found sig- nificantly elevated levels of several antibody isotypes in nasal polyp tissue.46)
Recent study reported the elevated levels of auto-antigen specific antibodies in nasal polyp tissue.47) The presence of autoantibodies suggests that the microenvironment of a nasal polyp promotes the expansion of self-reactive B-cell clones. Although the pathogenicity of these antibodies is unclear, the presence of increased anti-dsDNA antibody levels is associated with a clinically more aggressive form of CRS with nasal polyps.
Regulation of other cells by epithelium
The potential importance of mast cells has been con- sidered in pathogenesis of CRS because they produce various cytokines that activate eosinophils, molecules that promote tissue remodeling and chemical mediators that produce tissue edema.48) Recent study has demonstrated a unique localization of mast cells within the glandular epi- thelium of NPs and suggested that mast cells in NPs have distinct phenotypes that vary by tissue location. Glandular mast cells and the diverse subsets of mast cells may con- tribute to the pathogenesis of CRSwNP.48)
Chemokines mainly participate in the selective recruit- ment of inflammatory cells into tissue sites and several chemokines have also been reported to be involved in CRS. The regulation and production of chemokines in- volved in the recruitment of CCR3+ cells (eg, eosino- phils), such as RANTES (CCL5) and eotaxins (CCL11 [eotaxin-1], CCL24 [eotaxin-2], and CCL26 [eotaxin-3]) have been studied in patients with CRSwNP.49) 50) In addi- tion, levels of several other chemokines, including CCL2 (monocyte chemoattractant protein 1), CCL20 (liver and activation regulated chemokine), CXCL8 (IL-8), CXCL12
(stromal cell–derived factor 1a), and CXCL13 (B cell–at- tracting chemokine 1), have been reported to be increased in patients with CRS.51) 52) It has been reported that CCL23 which has chemotactic activity for monocytes, dendritic cells, and lymphocytes through the chemokine receptor CCR1, can induce endothelial cell migration, tube forma- tion, and angiogenesis through CCR1.
Poposki et al. reported that eosinophils produce the chemokine CCL23 and that patients with eosinophilic CRS have increased levels of CCL23 in polypoid tissue.53) In their study, CCL23 is identified as a novel marker of pa- tients with eosinophilic CRSwNP and the overproduction of CCL23 in NPs might contribute to the pathogenesis of eosinophilic CRSwNP. CCL18 is known to recruit naive T cells, B cells, and immature dendritic cells, as well as to activate fibroblasts. CCL18 is thought to be involved in Th2-related inflammatory diseases, including asthma and atopic dermatitis. Recently, Peterson et al. reported that M2 macrophages and mast cells produce the chemokine CCL18 and that the patients with CRSwNP have increased levels of CCL18 in sinonasal and polyp tissues. Their findings suggested that overproduction of CCL18 in NPs might contribute to the pathogenesis of CRSwNP through recruitment of lymphocytes and dendritic cells, activation of fibroblasts, and initiation of local inflammation.54)
CONCLUSION
The sinonasal epithelium provides a mechanical and innate immune barrier to various environmental agents.
Defects in the mechanical barrier and innate immune re- sponse also play a key role in regulating the adaptive im- mune response in the nose and paranasal sinus mucosa.
Alterations are reported in the various components of the epithelial innate immune system including epithelial ex- pressed PRRs and the levels of antimicrobial innate im- mune effector molecules. Recent studies show in CRS that epithelial-derived cytokines, chemokines and inducible surface proteins are involved in recruiting and activating cells of the adaptive immune system with development of the clinical symptoms characteristic of the disease. Fur- ther studies on host immune dysfunction in CRS will be necessary to generate a comprehensive understanding of the pathophysiology of CRS and to identify novel targets for drug development.
REFERENCES
1) Van Cauwenberge P, Van Hoecke H, Bachert C. Pathogenesis of chronic rhinosinusitis. Curr Allergy Asthma Rep 2006; 6:487–
2) Zuckerman JD, Lee WY, DelGaudio JM, Moore CE, Nava P, 94.
Nusrat A,et al. Pathophysiology of nasal polyposis: the role of desmosomal junctions. Am J Rhinol 2008; 22:589– 97.
3) Dejima K, Randell SH, Stutts MJ, Senior BA, Boucher RC. Poten- tial role of abnormal ion transport in the pathogenesis of chronic sinusitis. Arch Otolaryngol Head Neck Surg 2006; 132:1352–62.
4) Richer SL, Truong-Tran AQ, Conley DB, Carter R, Vermylen D, Grammer LC, et al. Epithelial genes in chronic rhinosinusitis with and without nasal polyps. Am J Rhinol 2008; 22:228–34.
5) Tieu DD, Kern RC, Schleimer RP. Alterations in epithelial barrier function and host defense responses in chronic rhinosinusitis. J Allergy Clin Immunol 2009; 124:37–42.
6) Kauffman HF, Tamm M, Timmerman JA, Borger P. House dust mite major allergens Der p 1 and Der p 5 activate human airway- derived epithelial cells by protease-dependent and protease-inde- pendent mechanisms. Clin Mol Allergy 2006; 4:5.
7) Briot A, Deraison C, Lacroix M, Bonnart C, Robin A, Besson C, et al. Kallikrein 5 induces atopic dermatitis-like lesions through PAR2-mediated thymic stromal lymphopoietin expression in Netherton syndrome. J Exp Med 2009; 206:1135–47.
8) Rudack C, Steinhoff M, Mooren F, Buddenkotte J, Becker K, von Eiff C, et al. PAR-2 activation regulates IL-8 and GRO- alpha synthesis by NF-kappaB, but not RANTES, IL-6, eotaxin or TARC expression in nasal epithelium. Clin Exp Allergy 2007;
37:1009–22.
9) Vroling AB, Fokkens WJ, van Drunen CM. How epithelial cells detect danger: aiding the immune response. Allergy 2008;
63:1110–23.
10) Sha Q, Truong-Tran AQ, Plitt JR, Beck LA, Schleimer RP. Acti- vation of airway epithelial cells by toll-like receptor agonists. Am J Respir Cell Mol Biol 2004; 31:358–64.
11) Lane AP, Truong-Tran QA, Myers A, Bickel C, Schleimer RP.
Serum amyloid A, properdin, complement 3, and toll-like recep- tors are expressed locally in human sinonasal tissue. Am J Rhinol 2006; 20:117–23.
12) Claeys S, de Belder T, Holtappels G, Gevaert P, Verhasselt B, van Cauwenberge P, et al. Human beta-defensins and toll-like recep- tors in the upper airway. Allergy 2003; 58:748–53.
13) Vroling AB, Fokkens WJ, van Drunen CM. How epithelial cells detect danger: aiding the immune response. Allergy 2008;
63:1110–23.
14) Kato A, Schleimer RP. Beyond inflammation: airway epithelial cells are at the interface of innate and adaptive immunity. Curr Opin Immunol 2007; 19:711–20.
15) Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immu- nol 2004;4: 499-511.
16) Wang CS, Dong Z. Expression of toll-like receptor mRNA in epi- thelial cell of nasal mucosa. Zhonghua Er Bi Yan Hou Ke Za Zhi 2003;38:243-6.
17) Kim ST, Cha HE, Kim DY, Han GC, Chung YS, Lee YJ,et al.
Antimicrobial peptide LL-37 is upregulated in chronic nasal in- flammatory disease. Acta Otolaryngol 2003; 123:81–5.
18) Lee HM, Park IH, Woo JS, Chae SW, Kang HJ, Hwang SJ. Up- regulation of surfactant protein A in chronic rhinosinusitis. Laryn- goscope 2006; 116:328–30.
19) Psaltis AJ, Bruhn MA, Ooi EH, Tan LW, Wormald PJ. Nasal mu- cosa expression of lactoferrin in patients with chronic rhinosinus- itis. Laryngoscope 2007; 117:2030–5.
20) Eckert RL, Broome AM, Ruse M, Robinson N, Ryan D, Lee K.
S100 proteins in the epidermis. J Invest Dermatol 2004; 123:23–
21) Meyer JE, Harder J, Sipos B, Maune S, Klöppel G, Bartels J,et 33.
al. Psoriasin (S100A7) is a principal antimicrobial peptide of the human tongue. Mucosal Immunol 2008; 1:239–43.
22) Seshadri S, Lin DC, Rosati M, Carter RG, Norton JE, Suh L,et al. Reduced expression of antimicrobial PLUNC proteins in na- sal polyp tissues of patients with chronic rhinosinusitis. Allergy 2012;67:920–28.
23) Ramanathan M, Spannhake EW, Lane AP. Chronic rhinosinusitis with nasal polyps is associated with decreased expression of mu- cosal interleukin 22 receptor. Laryngoscope 2007; 117:1839–43.
24) Schleimer RP, Kato A, Peters A, Conley D, Kim J, Liu MC,et al.
Epithelium, inflammation, and immunity in the upper airways in humans. Proc Am Thorac Soc 2009;6:288–94.
25) Tewfik MA, Bossé Y, Hudson TJ, Vallée-Smejda S, Al-Shemari H, Desrosiers M. Assessment of Toll-like receptor 2 gene poly- morphisms in severe chronic rhinosinusitis. J Otolaryngol Head Neck Surg 2008;37(4):552–8.
26) Castano R, Bossé Y, Endam LM, Desrosiers M. Evidence of asso- ciation of interleukin-1 receptor-like 1 gene polymorphisms with chronic rhinosinusitis. Am J Rhinol Allergy 2009;23(4):377– 84.
27) Endam LM, Bossé Y, Filali-Mouhim A, Cormier C, Boisvert P, Boulet LP, et al. Polymorphisms in the interleukin-22 receptor alpha-1 gene are associated with severe chronic rhinosinusitis.
Otolaryngol Head Neck Surg 2009;140 (5):741–7.
28) Van Drunen CM, Mjösberg JM, Segboer CL, Cornet ME, Fok- kens WJ. Role of Innate Immunity in the Pathogenesis of Chronic Rhinosinusitis: Progress and New Avenues. Curr Allergy Asthma Rep 2012;12:120–6.
29) Kern RC, Conley DB, Walsh W, Chandra R, Kato A, Tripathi-Pe- ters A, et al. Perspectives on the etiology of chronic rhinosinusitis:
An immune barrier hypothesis Am J Rhinol 2008;22: 549–59.
30) Leonard WJ. TSLP: finally in the limelight. Nat Immunol 2002;3:605–7.
31) Kato A, Favoreto S, Avila PC, Schleimer RP. TLR3- and Th2 cy- tokine-dependent production of thymic stromal lymphopoietin in human airway epithelial cells. J Immunol 2007;179:1080–7.
32) Liu YJ. TSLP in epithelial cell and dendritic cell cross talk. Adv Immunol 2009; 101:1–25.
33) Moniaga CS, Jeong SK, Egawa G, Nakajima S, Hara-Chikuma M, Jeon JE, et al. Protease activity enhances production of thymic stromal lymphopoietin and basophil accumulation in flaky tail mice. J Exp Med 2009; 206:1135–47.
34) Sanchez-Segura A, Brieva JA, Rodriguez C. T lymphocytes that infiltrate nasal polyps have a specialized phenotype and produce a mixed TH1/TH2 pattern of cytokines. J Allergy Clin Immunol 1998; 102:953–60.
35) Zhang N, Van Zele T, Perez-Novo C, Van Bruaene N, Holtappels G, DeRuyck N, et al. Different types of T-effector cells orches- trate mucosal inflammation in chronic sinus disease. J Allergy Clin Immunol 2008; 122:961–968
36) Van Bruaene N, Pérez-Novo CA, Basinski TM, Van Zele T, Hol- tappels G, De Ruyck N, et al. T-cell regulation in chronic parana- sal sinus disease. J Allergy Clin Immunol 2008; 121:1435–41.
37) Kurosawa S, Myers AC, Chen L, Ni J, Plitt JR, Heller NM, et al.
Expression of the costimulatory molecule B7-H2 by human air- way epithelial cells. Am J Respir Cell Mol Biol 2003;28:563-73.
38) Kim J, Myers AC, Chen L, Pardoll DM, Truong-Tran QA, Lane AP,et al. Constitutive and inducible expression of B7 family of ligands by human airway epithelial cells. Am J Respir Cell Mol Biol 2005;33:280-9.
39) Wise SK, Ahn CN, Schlosser RJ. Localized immunoglobulin E expression in allergic rhinitis and nasal polyposis. Curr Opin Oto- laryngol Head Neck Surg 2009; 17:216–22.
40) Kato A, Peters A, Suh L, Carter R, Harris KE, Chandra R, et al.
Evidence of a role for B cell-activating factor of the TNF family in the pathogenesis of chronic rhinosinusitis with nasal polyps. J Allergy Clin Immunol 2008; 121:1385–1392.
41) Kato A, Xiao H, Chustz RT, Liu MC, Schleimer RP. Local release of B cell-activating factor of the TNF family after segmental aller- gen challenge of allergic subjects. J Allergy Clin Immunol 2009;
123:369–75.
42) Patadia M, Dixon J, Conley D, Chandra R, Peters A, Suh LA, et al. Evaluation of the presence of B-cell attractant chemokines in chronic rhinosinusitis. Am J Rhinol Allergy 2009;24:11-6.
43) Peters AT, Kato A, Zhang N, Conley DB, Suh L, Tancowny B, et al. Evidence for altered activity of the IL-6 pathway in chronic rhinosinusitis with nasal polyps. J Allergy Clin Immunol 2010;125:397-403.
44) Bachert C, Zhang N, Holtappels G, De Lobel L, van Cauwen- berge P, Liu S, et al. Presence of IL-5 protein and IgE antibodies to staphylococcal enterotoxins in nasal polyps is associated with comorbid asthma. J Allergy Clin Immunol 2010; 126:962-8.
45) Sanchez-Segura A, Brieva JA, Rodriguez C. Regulation of im- munoglobulin secretion by plasma cells infiltrating nasal polyps.
Laryngoscope 2000; 110:1183-8.
46) Hulse KE, Norton JE, Suh L, Zhong Q, Mahdavinia M, Simon P, et al. Chronic Rhinosinusitis with Nasal Polyps is Characterized by B Cell Inflammation and EBI2 Expression. J Allergy Clin Im- munol 2013;131:1075-83.
47) Tan BK, Li QZ, Suh L, Kato A, Conley DB, Chandra RK, et al.
Evidence for intranasal antinuclear autoantibodies in patients with chronic rhinosinusitis with nasal polyps. J Allergy Clin Immunol 2011; 128:1198-206.
48) Takabayashi T, Kato A, Peters AT, Suh LA, Carter R, Norton J, et al. Glandular mast cells with distinct phenotype are highly el- evated in chronic rhinosinusitis with nasal polyps. J Allergy Clin Immunol 2012;130:410-20.
49) Beck LA, Stellato C, Beall LD, Schall TJ, Leopold D, Bickel CA, et al. Detection of the chemokine RANTES and endothe- lial adhesion molecules in nasal polyps. J Allergy Clin Immunol 1996;98:766-80.
50) Jahnsen FL, Haye R, Gran E, Brandtzaeg P, Johansen FE. Gluco- corticosteroids inhibit mRNA expression for eotaxin, eotaxin-2, and monocyte-chemotactic protein-4 in human airway inflamma- tion with eosinophilia. J Immunol 1999;163:1545-51.
51) Marcella R, Croce A, Moretti A, Barbacane RC, Di Giocchino M, Conti P. Transcription and translation of the chemokines RANT- ES and MCP-1 in nasal polypsand mucosa in allergic and non- allergic rhinopathies. Immunol Lett 2003;90:71-5.
52) Meyer JE, Bartels J, Gorogh T, Sticherling M, Rudack C, Ross DA, et al. The role of RANTES in nasal polyposis. Am J Rhinol 2005;19:15-20.
53) Poposki JA, Uzzaman A, Nagarkar DR, Chustz RT, Peters AT, Suh LA, et al. Increased expression of the chemokine CCL23 in eosinophilic chronic rhinosinusitis with nasal polyps. J Allergy Clin Immunol 2011;128:73-81.
54) Peterson S, Poposki JA, Nagarkar DR, Chustz RT, Peters AT, Suh LA, et al. Increased expression of CC chemokine ligand 18 in patients with chronic rhinosinusitis with nasal polyps. J Allergy Clin Immunol 2012;129:119-27.
55) Schleimer RP, Kato A, Kern RC, Kuoerman D, Avila PC. Epithe- lium: at the interface of innate and adaptive immune responses. J Allergy Clin Immunol 2007;120:1279-84.