1. 서 론
건강에 대한 관심의 증대로 대부분의 식품 구매자들에게 식품의 기능성 소재 함유 여부가 중요 한 구매 결정 기준이 되어 가고 있다. 축산물을 제외한 국내 식품산업의 매출액은 2006년 통계 약 35조원으로서 전년대비 7.8%의 증가율을 보이며 꾸준한 성장세를 보였고, 이 중 2005년 대비 11 개 품목의 출하액은 증가한 반면, 9개 품목은 출하액이 감소한 것으로 나타났다. 이 중 2006년 도 식품 및 식품첨가물 생산 실적에 의한 국내 생산량 기준 국민 다소비 식품의 순위에서 밀가 루가 1위 그리고 곡류 가공품도 상위에 속해 있어 중요한 식품 소재로 이용되고 있는 실정이다 (2007 보건산업백서/한국보건산업진흥원).
밀가루와 곡류 가공품들은 최근 많은 관심을 받고 있는, 생리활성 소재들인 식이섬유류와 phytochemical류 등을 다량 함유하고 있어, 식생활의 서구화에 대비하여 소비가 지속적으로 증가 하고 있다. 또한 최근의 고식이섬유 식사에 대한 관심의 증대로 인해, 식이섬유 함량이 높은 통곡 물을 원료로 한 식품에 대한 관심과 소비가 증대되고 있고, 특히 도정이 덜 된 통곡물의 경우는 높은 식이섬유류, 미네랄 함량으로 건강 증진에 도움을 줄 수 있는 것으로 알려져 왔다. 그러나 실제로 이러한 대부분의 곡류 내의 무기질들은 많은 부분이 phytic acid와 같은 특정 유기물질과 강하게 결합하고 있어서 섭취 시 생체 이용성에 많은 어려움이 있는 것으로 보고되고 있다(Latta et al, 1980; Wodzinski et al, 1996; Sirkka, 1997). 특히 phytic acid는 단위 동물의 소화기관 내에는 이 의 분해를 위한 효소인 phytase가 없거나 제한적으로 활성을 나타내기 때문에(Henric et al, 2002) 소화가 어려운 난소화성 물질로 널리 알려져 있다.
Phytic acid(myo-inositol hexakisphosphate)는 곡류 또는 종자식물의 씨앗이 성장하는 중에 생성 되는 화합물로서, 곡류와 콩과식물의 씨앗 중 인과 inositol의 주요한 저장형태이다. 곡류 중 총 인의 70~80%가 phytic acid 상태로 존재하며 구조적으로는 단백질과 결합된 마그네슘염 또는 칼
식품 내 mineral 이용성 증진을 위한 새로운 phytase의 적용 가능성 탐색
어중혁 중앙대학교 식품공학과
슘염의 형태로 존재하고 있는 것으로 알려져 있다(Latta et al, 1980). 주요 곡물 내에는 중량 대비로 1~5%가량 존재하여 이들의 대부분은 1가 혹은 2가 양이온들(K+, Ca++, Mg++)과 강하게 결합하고 있다(Greiner, 2006). 이러한 독특한 형태의 결합물 형성은 특히 단위 동물의 영양에 중요한 미량의 다가 양이온들(Zn++, Ca++, Mg++, Fe+++)과의 불용성의 염 형성으로 이들의 생체 내 이용성을 저하시 키며, 이 외에도 아미노산, 비타민 등과도 결합하여 이들의 흡수를 저해하는 역할을 하는 것으로 보고되고 있다(Wodzinski et al, 1996). 따라서 이러한 고식이섬유 식품에 함유된 무기질의 생체 내 이용성 증진을 위해서는 phytate와 무기질과의 결합을 분해하여 이들의 생체 내 이용성을 높이는 것이 매우 중요하다 할 수 있다. 일반적으로 phytate를 제거하기 위한 방법으로는 열처리, 화학적 처리, 그리고 효소적 처리 방법(Frontela et al, 2008)이 있지만 열 및 화학적 처리방법은 다른 영양소 의 추가적인 손상을 초래하기 때문에 phytase와 같은 효소제의 사용이 가장 이상적인 방법으로 보고되고 있다(Martinez et al, 1996 Greiner and Konitzny, 2006).
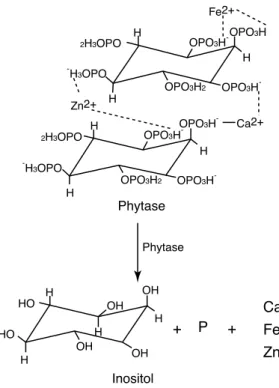
Phytase(myo-inositol hexakisphosphate phosphohydrolase)는 phytic acid를 분해하여 유리 phosphate 와 phosphate inositol 및 inositol을 생성하는 효소로서 인체 내에서는 분비되지 못하는 효소이다(그 림 1). Phytase는 최적 작용 pH에 따라 acid phytase와 alkaline phytase로 구분되고(Oh et al, 2004), 현 재까지는 주로 phytic acid에 의한 환경 오염을 줄이고, 동물 사료 내 미량 무기질의 이용성 증대
Phytase
+ P + Ca Fe Zn Phytase
2H3OPO
2H3OPO
H
H H
H H
H
H
H H
H HO
HO OH OH
OH OH
OPO3H
OPO3H2
OPO3H2
Fe2+
Ca2+
Zn2+
Inositol H3OPO
H3OPO
OPO3HOPO3H
OPO3H OPO3H OPO3H
-
-
-
- -
- -
그림 1 Phytate hydrolysis by phytase into inositol, phosphate, and other divalent cations.
를 위한 목적으로 축산 및 사료 산업에서 acid phytase의 형태만 이용되어 오고 있다(Nermin et al, 2006; Maria et al, 2008; Merete et al, 2007; Liu et al, 2008). 현재 산업적으로는 acid phytase의 일종인 Natuphos 만이 사료 산업에서 사용되고 있고, 식품 분야에서는 sour dough와 같은 산성 pH의 식 품에 활용하고자 하는 부분적인 시도의 사례만 보고되고 있는 실정이다(Porres et al, 2001; Greiner
& Konietzny, 2006). 특히 식품 산업에서는 아직까지 본격적인 사용이 시도되지 못하고 있으며, 이 는 현재까지 알려진 대부분의 phytase들이 산성 영역에서 작용하는 acid phytase들이었던 것이 가 장 큰 문제점이었던 것으로 보고되고 있다(Reddy & Shridhar, 2002). 따라서 본 연구에서는 본 연 구실에서 보유하고 있는 새로운 alkaline phytase와 기존의 acid phytase들의 특성을 비교하고 이들 의 식품에의 적용 가능성을 탐색하여, 이후 산업적 이용에 기초 자료로 활용하고자 하였다.
2. 연구방법
2.1 Materials
Whole wheat flour(Bob’s Red Mill natural Food. Inc. USA)는 local market에서 구입하여 사용하였 고 enzyme은 실험실에 보유중인 alkaline phytase(DS11)와 wheat phytase(E.C. 3.1.3.26, Sigma, USA), Natuphos (BASF, Germany)를 사용하였다. Sodium phytate는 Sigma사의 시약을 구입하여 사용하 였고, 그 외의 시약들은 chemical grade의 것을 구매하여 사용하였다.
2.2 Alkaline phytase(DS11)의 분리 및 정제
본 연구실에서 보관중인 Bacillus amylolique f aciens 유래 phytase gene을 형질전환시킨 Escherichia coli BL21 균주를 37℃, LB-Kanamycin에서 배양하였다. 1mM IPTG(Isopropyl β-D-1- thiogalactopyranoside)를 induction 후, 배양액을 8,655 ×g에서 10분간 원심분리하여 균체를 취하고 5mM CaCl2/ 50mM Tris-HCl(pH 7.0) 완충액에 현탁하여 French press(SLM Instruments. INC)로 세 포를 파쇄하였다. 이후 16,400 ×g에서 30분간 원심분리하여 얻은 상등액을 70℃ 항온 수조에 25 분간 처리한 뒤 16,400 ×g에서 30분간 원심분리하여 상등액을 회수하여, 조효소액을 분리하였다.
준비된 조효소액을 Vivaspin 20(Satorius, Germany)을 사용하여 농축하고, 4℃에 보관하며 실험에 사용하였다.
2.3 단백질 농도 측정
단백질 농도는 Bio-Rad protein assay(Bio-Rad Lab., USA)를 사용하여 595nm에서의 흡광도를 측정 하였으며, bovine serum albumin을 기준 물질로 사용하여 농도를 계산하였다.
2.4 SDS-PAGE
Sodium dodecyl sulfate polyacrylamide gel electrophoresis(SDS-PAGE)는 Laemmli(Laemmli, 1970)의 방법에 따라 12% acrylamide gel을 사용하였으며 coomassie brilliant blue R-250을 사용하여 염색하 였다.
2.5 Phytase의 역가 측정
2mM sodium phytate를 이용하여 alkaline phytase(DS11)의 효소 역가를 측정하였다. 2mM sodium phytate/ 2mM CaCl2/ 50mM Tris-HCl(pH 7.0) 450µl에 alkaline phytase(DS11) 50µl를 첨가한 후 50℃
항온수조에서 90분간 반응시켰다. 반응액에 500µl의 발색시약(2.5% ammonium heptamolybdate, 0.175% ammonia, 0.1425% ammonium vanadate, 22.75% nitric acid)을 가하여 15분간 상온에서 반 응시킨 후 415nm의 흡광도로 해리되는 Pi의 함량을 측정하였다(Engelen et al. 1994). g효소의 1 unit은 pH 7.0, 50℃ 조건에서 sodium phytate로부터 1분당 1µM의 Pi를 해리시키는 효소량으로 정 하였다. Wheat phytase의 경우는 위와 동일하나, pH 5.15, 55℃ 조건에서 phytate를 1분당 1µM의 Pi 를 해리시키는 효소량으로 정하였다.
2.6 pH에 따른 phytase 반응 특성 분석
효소반응은 Engelen 등(1994)의 방법을 변형하여 사용하였다. 10ml 2mM sodium phytate/ 2mM CaCl2을 기질로 사용하였으며 pH는 50mM Tris-HCl(pH7.0)과 100mM sodium acetate(pH 5.0)를 사용하여 조절하였다. 기질 용액에 여러 수준의 정해진 phytase를 가하고 50℃ 항온수조에서 100 분까지 반응시켰다. 이때 20분 간격으로 반응액을 1ml씩 취하여 168µl의 20% TCA를 가하고 0℃
에 10분간 방치하여 효소 반응을 정지시켰다. Microwellplate에 용액을 희석한 반응액 100µl를 넣 은 뒤 100µl의 발색시약(2.5% ammonium heptamolybdate, 0.175% ammonia, 0.1425% ammonium vanadate, 22.75% nitric acid)을 가하여 15분간 반응시킨 후 Spectra max 190(Molecular devices USA) 로 415nm에서 흡광도를 측정하여 해리되는 Pi의 함량을 측정하였다.
2.7 Whole wheat bread 제조
빵 반죽은 통밀가루(100g), dry yeast(3.0g), 소금(2.0g), 물(57.3ml)에 phytase를 첨가하여 반죽하였 다. 10분간 반죽 후, 약 50g씩 나누어 모양을 형성하였다. 형성된 반죽은 29℃ 발효기에서 30분 혹 은 60분간 발효시킨 뒤 170℃ 오븐에서 27분간 baking하였다. 모든 빵은 상온에서 2시간 식힌 후 에 동결건조하여 이후 분석 시료로 사용하였다(Monica et al, 2001).
2.8 Phytate 분해산물의 분석
TLC
Cellulose 코팅된 glass plate는 Merck사(Darmstadt, Germany)의 것을 사용하였다. 시료는 plate 밑 에서 1.5cm의 간격을 두고 spot에 약 8µl가 되도록 spotting하였고, 이동상은 20ml 1-propanol : 25%
ammonia solution : water(5 : 4 : 1)를 사용하여 Camag사(Muttenz, Switzerland)의 double-through chamber(내부 크기: 30×27×10cm)에서 전개용매가 plate 상부의 5~10mm까지 올라가도록 약 12시 간 전개시켰다(Hatazack & Rasmussen, 1998). 전개가 끝난 plate를 후드에서 풍건한 후에 molybdate reagent(8mM ammonium heptamolybdate tetrahydrate, 0.1M HCl, 0.5M HClO4)를 분무하고, 85℃에 서 6.5분간 반응시킨 후 6.5분간 UV-light(254nm)를 사용하여 발색시켰다. Spot은 푸른색으로 나 타났으며 2시간 후에 최대 감도를 보였다.
HPIC
분석을 위한 시료는 Erika 등(1997), Pascale 등(2000)의 방법을 변형하여 전처리한 후 분석에 사용 되었다. Screw-cap test tube에 시료 0.2g을 칭량한 후 0.5M HCl을 10ml를 가하고 끓는 물에서 5분 간 교반하였다. 이것을 3,380×g에서 10분간 원심분리를 한 뒤 상등액을 회수하여 사용하였다.
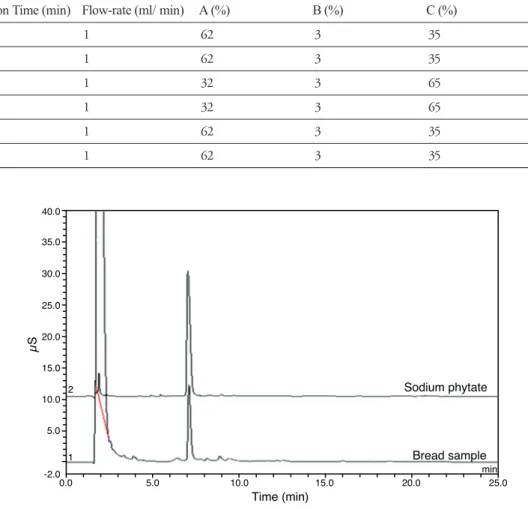
HPIC는 ICS-3000 system(Dionex, USA)과 conductivity detector(Dionex, USA), anion suppressor(ASRS 300 4-mm)를 사용하였고, Omnipac Pax-100(4×250mm, Dionex, USA) column 을 사용하여 분석하였다. 용매는 증류수(A), 50% isopropyl alcohol(B)(Burdick & Jackson, HPLC grade), 그리고 200mM sodium hydroxide solution(C)(Fisher Scientific, USA)을 표 1과 같은 조건으로 사용하였다(Erika et al, 1997).
표 1 Gradient elution program for the separation of phytic acid.
Elution Time (min) Flow-rate (ml/ min) A (%) B (%) C (%)
0.00 1 62 3 35
4.00 1 62 3 35
4.01 1 32 3 65
15.00 1 32 3 65
15.01 1 62 3 35
25.00 1 62 3 35
Time (min)
µS
Sodium phytate
-2.00.0 5.0
1 10.0 2 15.0 20.0 25.0 30.0 35.0 40.0
5.0 10.0 15.0 20.0
min 25.0
Bread sample
그림 2 Elution profile of phytic acid standard and the bread sample on an Omnipac Pax-100 column.
Inductively Coupled Plasma-Optical Emission Spectrometer
Minerals는 ICP-OES(inductively coupled plasma-optical emission spectrometer) optima 5300DV system(Perkin Elmer, USA)을 사용하여 분석하였다. 기준 물질은 Pi, Ca, Mg, Fe, Zn을 사용하였고, Pi와 Mg(50, 100, 200, 400, Ca: 10, 25, 50, 100ppm), Fe과, Zn(1, 3, 9, 27ppm)을 사용하여 검출 곡선 을 작성하였다.
Total minerals 함량은 건식 분해법을 사용하여 시료를 분해한 뒤 측정하였다. 시료 1g을 도가니 에 넣어 560℃ 회화로에서 4시간 회화시킨 후 상온에서 식힌 뒤에 35.5% HNO3로 가하고 다시 2 시간 동안 회화시켰다. 회화가 끝난 시료는 상온에서 식힌 뒤 4ml 17.5% HCl을 가한 후 2시간을 방치 후, 증류수로 희석하여 분석에 사용하였다. 효소의 작용에 의해 해리된 유리 미네랄의 함량 은 시료 0.5g에 10ml의 증류수를 가하고 5분간 교반한 후 여과하여 분석하였다.
2.9 통계처리
통계분석은 SAS(Statistical Analysis System, USA) program을 통하여 분석하였고, 처리구간의 평균 간 비교는 Duncan의 다중검정을 통하여 유의성 검정(p<0.05)을 실시하였다.
3. 결과 및 고찰
3.1 Alkaline phytase(DS11)의 부분 정제
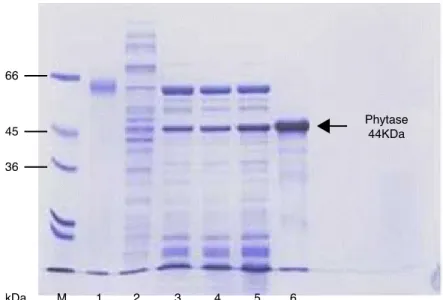
E. coli BL21에 발현시킨 B. amyloliquef aciens DS11 phytase는 내열성을 이용하여 부분 정제하였 다. 70℃에서의 열처리를 통하여 부분적으로 정제된 조효소액을 얻을 수 있음을 확인하였다(그 림 3). 열처리와 이후 농축 과정을 통해서 효소의 specific activity는 약 7배 증가(0.48 → 3.41unit/
mg protein)한 것을 확인하였고, 또한 Ni-NTA column으로 정제된 His-tagged phytase를 사용하 여 44kDa의 분자량을 가진 최종 활성 효소를 확인하였다. 기존에 보고된 phytase들의 경우는
Aspergillus 유래 phytase들은 A. terrus(214kDa), A. ficuum(60kDa)(Luis et al, 1997), A. niger(84kDa) (Anne and Gary, 2003)의 분자량을 가지는 것으로 알려져 있고, Bacillus species는 36~38kDa, E. coli 유래의 phytase 46kDa(Miksch et al, 2002), 그리고 Sporotrichum thermophile는 약 90kDa(Singh and Satyanarayana, 2009)의 분자량을 가지는 것으로 보고되고 있어, 다양한 분자량들을 가지는 것으 로 확인되고 있다. 본 연구에 사용된 Bacillus 유래의 phytase(DS11)는 44kda의 분자량을 가지며, 또한 Ca 존재 시에 약 90℃까지 열 안정성을 가지는 것으로 보고되고 있다(Kim et al, 1998). 지금 까지 보고된 대부분 phytase들의 열 안정성 한계 온도는 Enterobacter sp.의 60℃(Ashok et al, 2001), B. subtilis의 60℃(Tye et al, 2002), E. coli의 50℃(Miksch et al, 2002)로서 DS11 phytase에 비해 현저히 낮은 것으로 확인되고 있어 큰 차별성을 보이고 있다. 또한 이러한 열 안정성은 효소 정제 과정의 단순화와 이를 기반으로 한 산업적 이용에 있어 많은 장점을 가질 수 있을 것으로 기대된다.
3.2 효소의 작용 특성 및 pH에 따른 반응 특성
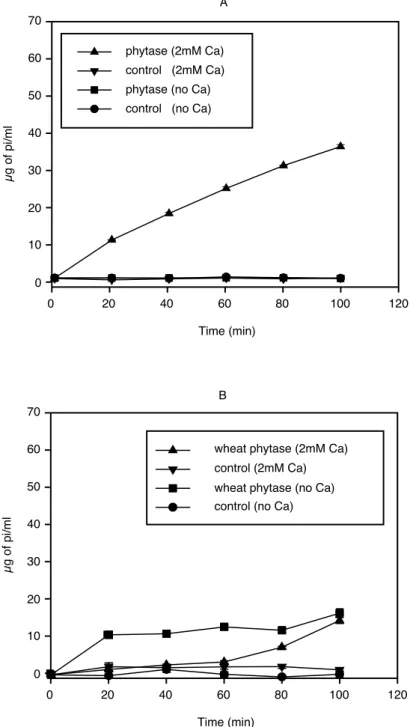
2mM phytate(50mM Tris-HCl pH 7.0)를 기질로 하여, alkaline phytase(0.018unit/mg phytate)와 wheat phytase(0.018unit/mg phytate)를 처리하고 시간별로 반응 추이를 확인하였다(그림 4). 2mM Ca을 cofactor로 첨가한 alkaline phytase 처리군에서만 반응이 진행되어, 100분 반응 후 약 35mg 의 Pi가 생성되어 alkaline phytase의 경우 Ca이 cofactor로서 반응에 중요한 역할을 하고 있음을 알 수 있었다. Wheat phytase는 일반적으로 metal을 cofactor로 이용하지는 않는 것으로 알려져 있으
나, 본 반응에서는 Ca의 존재 유무에 관계없이 모든 조건에서 alkaline phytase에 비해 약한 활성 을 나타내었다(그림 4B). 이러한 현상은 중성 부근에서 최적의 활성을 나타내는 alkaline phytase 에 비해 약산성(pH 5.15)의 조건에서 최적의 활성을 보이는 wheat phytase의 특성에 기인한 것으 로 판단되며, 100분 반응 후에 해리된 Pi의 양은 alkaline phytase의 50% 수준으로 매우 낮게 나타 났다. 그러나 phytase에 의한 Pi의 생성은 각 효소들의 작용 패턴에 의해서 상대적으로 다르게 나 타날 수 있다. 가령 wheat phytase는 phytate에 함유된 6개의 Pi 중 5개의 Pi를 해리시키고, inositol monophosphate를 최종 산물로 생성하는 것으로 알려져 있다. 이러한 특성을 기반으로 이후 alkaline phytase와 비교하면 더욱 정확한 정보를 얻을 수 있을 것으로 기대되며, alkaline phytase의 중성 pH에서의 작용 특성은 많은 일반 가공 식품에의 적용에 있어 매우 중요한 의미를 가짐을 기 대할 수 있었다.
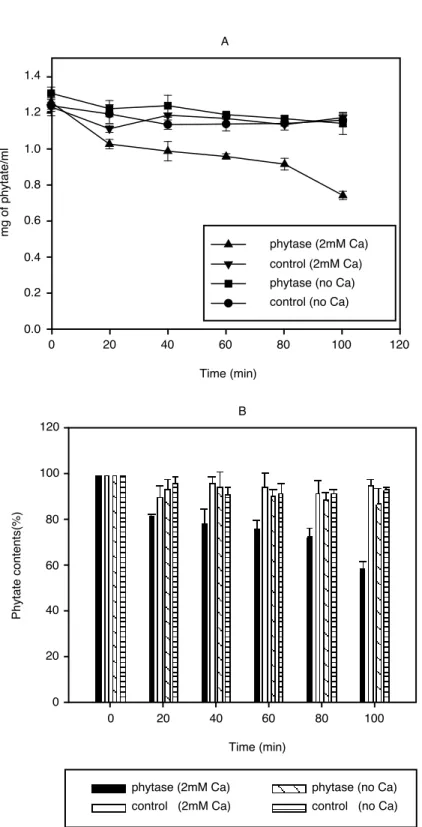
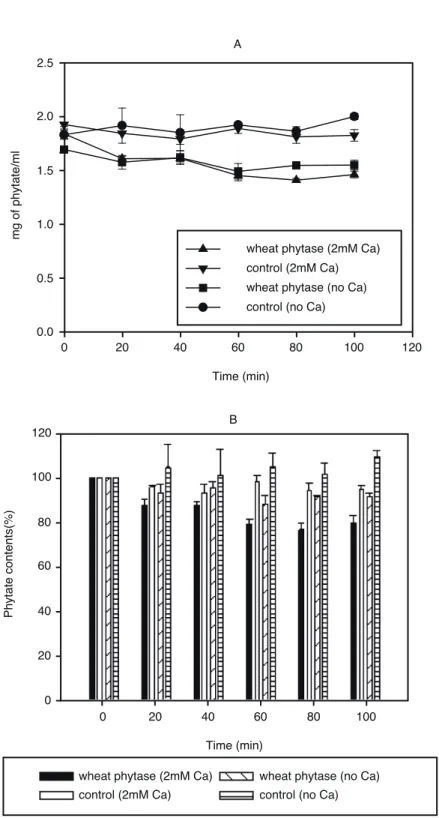
2mM sodium phytate(50mM Tris-HCl pH 7.0)에 phytase를 처리하고 시간별로 phytate의 양을 측 정하였다(그림 5, 6). Alkaline phytase(0.018unit/mg phytate)와 wheat phytase(0.018unit/mg phytate)를 가하고 50℃ 수조에서 시간별로 시료 내 phytate의 함량 변화를 측정하였다. 앞에서 확인한 Pi의 변화와 마찬가지로 alkaline phytase는 Ca이 함유되어 있는 기질에서만 반응이 일어났으며(그림 5), 상대적인 변화량을 비교하여 보면 시간에 따라 phytate가 효율적으로 분해되고 있음을 확인할 수 있었다(그림 5B). Wheat phytase의 경우에도 효소의 처리에 의해 약간의 phytate가 분해됨을 확 인할 수 있었으나, alkaline phytase에 비해 절대량 및 상대적 감소폭이 매우 적게 나타났다(그림 6).
또한 Pi의 변화와 마찬가지로 Ca의 존재 여부에 상관없이 기질인 phytate의 분해가 일어나고 있 음을 볼 수 있었다. 100분 반응 후, alkaline phytase 처리군에서는 약 40%의 phytate가 분해되었으 나, wheat phytase 처리군에서는 약 20%의 phytate만이 분해된 것으로 나타나 유의적인 차이를 보 였다. 이러한 현상 또한 지금까지 알려진 대부분의 phytase와 마찬가지로 약산성의 pH에서 주로 작용하는 wheat phytase의 작용 특성에 기인한 것으로서, 중성 및 약 알칼리 영역에서도 활성을 나타내는 alkaline phytase의 활용 가능성을 확인할 수 있었다.
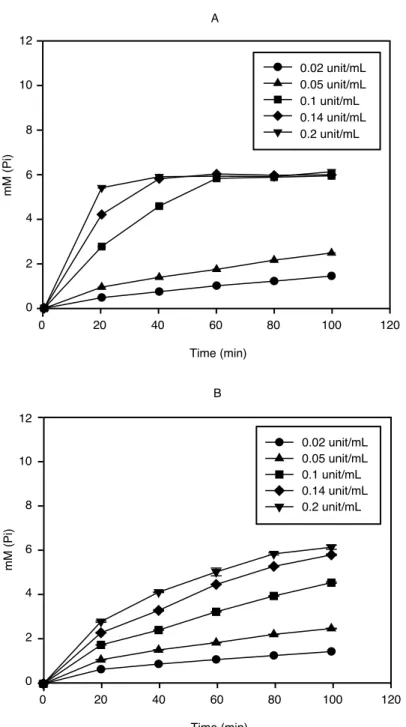
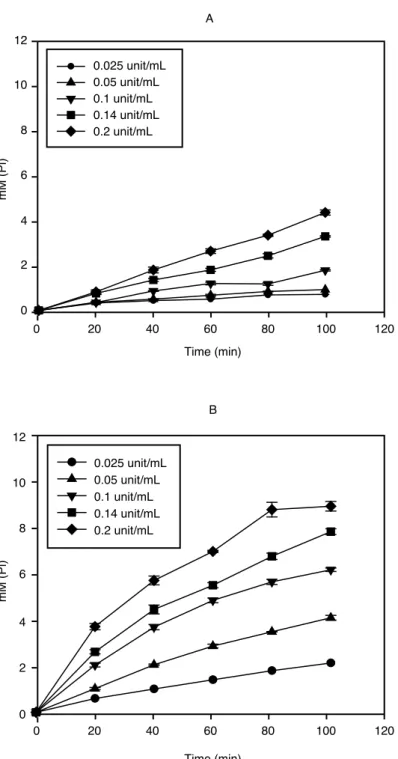
이러한 pH에 따른 효소들의 작용 특성을 산성(pH 5.0)과 중성(pH 7.0) 영역에서의 반응에 의한 Pi 생성 정도를 기준으로 비교, 측정해 보았다. Alkaline phytase는 pH 5보다 pH 7에서 더 높은 활 성을 나타내었고(그림 7), 이는 akaline phytase의 안정한 pH 범위가 4~8이고 pH 7에서 가장 좋은 활성을 가진다는 보고(Kim et al, 1998)와 일치하는 경향을 보였다. 효소에 의해 유리된 Pi함량으 로부터 alkaline phytase는 1M의 phytate에서 3M의 Pi를 생성하는 것을 확인할 수 있었고, 이것은 5M의 Pi를 유리시킨다고 알려져 있는 wheat phytase와 차별화되는 점이었다. 산성 영역에서 작용 하는 것으로 알려진 wheat phytase와 Natuphos 는 기존의 보고와 같이 pH 7보다 pH 5에서 높은 활성이 유지되었다. Wheat phytase는 pH 5.15의 산성 조건에서 최적의 활성을 나타내고 pH 6 이상 에서는 활성이 급격히 떨어진다는 연구(Peers, 1953)와 유사하게 pH 7에서 효소에 의한 유리 Pi의
kDa M 1
Phytase 44KDa
6 5 4 3 2 66
45 36
그림 3 SDS-PAGE analysis of Alkaline phytase (DS11) at each purification step from E.
coli BL21. The arrows indicated the phytase band. Lane M: M.W. maker; lane 1: BSA (66 kDa); lane 2: Cell extract; lane 3, 4: Heated at 70℃; lane 5: Concentrated; lane 6: Purified 6xHis-tagged phytase.
양이 유의적으로 낮게 나타났다(그림 8). 현재 사료 산업에서 이용되고 있는 또다른 acid phytase 의 하나인 Natuphos 도 pH 7보다는 pH 5에서 유리 Pi의 양이 높게 검출된 것으로 측정되어 산성 영역에서 활성이 높은 것으로 확인되었다(그림 9). pH 5에서 2mM의 phytate와 반응할 경우, 최고 10mM의 유리 Pi가 측정되었는데 이것은 Natuphos 는 본 연구에 사용된 alkaline phytase와 다르게 phytate를 분해하여 1개의 IP1(Inositol monophosphate)와 5개의 유리 Pi를 생성함을 보여준 것이다 (그림 9).
phytase (no Ca) phytase (2mM Ca)
control (no Ca) control (2mM Ca)
Time (min) A
µg of pi/ml
0 20 40 60 80 100 120
0 10 20 70 60 50 40 30
Time (min)
wheat phytase (no Ca) wheat phytase (2mM Ca)
control (no Ca) control (2mM Ca)
B
µg of pi/ml
0 20 40 60 80 100 120
0 10 20 70 60 50 40 30
그림 4 Time course analysis of Pi liberation from phytate.
(A) alkaline phytase(DS11); (B) wheat phytase.
Time (min)
mg of phytate/ml
phytase (2mM Ca) control (2mM Ca) phytase (no Ca) control (no Ca) A
0 20
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4
40 60 80 100 120
Time (min)
phytase (2mM Ca) phytase (no Ca) control (no Ca) control (2mM Ca)
B
Phytate contents(%)
20
0
0 20 40 60 80 100
40 60 80 100 120
그림 5 Time course analysis of phytate hydrolysis by alkaline phytase (0.018 unit/ mg phytate).
Time (min)
mg of phytate/ml
wheat phytase (2mM Ca) control (2mM Ca) wheat phytase (no Ca) control (no Ca) A
0 20
0.0 0.5 1.5
1.0 2.5
2.0
40 60 80 100 120
B
Phytate contents(%)
20
0
0 20 40 60 80 100
40 60 80 100 120
Time (min)
0 20 40 60 80 100
Time (min)
wheat phytase (2mM Ca) wheat phytase (no Ca) control (no Ca) control (2mM Ca)
그림 6 Time course analysis of phytate hydrolysis by wheat phytase (0.018 unit/mg phytate).
0.02 unit/mL 0.05 unit/mL 0.1 unit/mL 0.14 unit/mL 0.2 unit/mL
Time (min)
0 20 40 60 80 100 120
A
mM (Pi)
0 12 10 8 6 4 2
0.02 unit/mL 0.05 unit/mL 0.1 unit/mL 0.14 unit/mL 0.2 unit/mL
Time (min)
0 20 40 60 80 100 120
B
mM (Pi)
0 12 10 8 6 4 2
그림 7 Time course analysis of Pi liberation from phytate by alkaline phytase.
(A) pH 7; (B) pH 5.
0.025 unit/mL 0.05 unit/mL 0.1 unit/mL 0.14 unit/mL 0.2 unit/mL
Time (min)
0 20 40 60 80 100 120
A
mM (Pi)
0 12 10 8 6 4 2
0.025 unit/mL 0.05 unit/mL 0.1 unit/mL 0.14 unit/mL 0.2 unit/mL
Time (min)
0 20 40 60 80 100 120
B
mM (Pi)
0 12 10 8 6
4 2
그림 8 Time course analysis of Pi liberation from phytate by wheat phytase.
(A) pH 7; (B) pH 5.
0.025 unit/mL 0.05 unit/mL 0.1 unit/mL 0.14 unit/mL 0.2 unit/mL
Time (min)
0 20 40 60 80 100 120
A
mM (Pi)
0 12 10 8 6 4 2
0.025 unit/mL 0.05 unit/mL 0.1 unit/mL 0.14 unit/mL 0.2 unit/mL
0.025 unit/mL 0.05 unit/mL 0.1 unit/mL
0.14 unit/mL 0.2 unit/mL B
Time (min)
0 20 40 60 80 100 120
mM (Pi)
0 12 10 8 6 4 2
그림 9 Time course analysis of Pi liberation from phytate by Natuphos (A) pH 7; (B) pH 5.
3.3 TLC
효소 처리에 의한 기질 분해 패턴을 TLC를 이용하여 분석하였다. 현재까지 phytate(Inositol 6-phosphate, IP6)와 그 분해 산물들인 IP1, IP2, IP3, IP4, IP5, IP6의 물질은 명확한 분석법이 개발되 어 있지 못하나, TLC와 conductivity를 이용한 LC 분석법이 가장 유용한 방법으로 평가되고 있 다. 시료를 Ca 첨가군과 미첨가군으로 나눠서 alkaline phytase(0.018unit/mg of phytate)와 wheat phytase(0.018 unit/mg of phytate) 그리고 Natuphos 처리 후 50℃ 항온수조에서 반응을 시키고 20분 간격으로 샘플링하여 분석하였다. Reddy 등(2002)에 의하면 phytase는 phytate 분해 시에 모든 Pi를 동시에 분해하는 것이 아니라, penta-, tetra-, tri-, di-, monophosphate의 형태를 거치면서 분해한다. 일 반적으로 acid phytase는 여러 단계를 거쳐 최종 산물로 IP1과 5개의 Pi로 분해하는 것으로 보고되 고 있다. 본 연구에 사용한 alkaline phytase의 시간에 따른 분해 양상은 그림 10에 나타난 바와 같 이, Ca이 존재할 경우에만 기질의 분해가 일어났으며, 반응의 진행에 따라 Pi와 IP3가 생성되고 있 음을 확인할 수 있었다. 반면에 wheat phytase는 Ca의 존재 유무와 관계없이 반응 시간의 증가에 따라 phytate는 감소하고, Pi와 함께 IP2 혹은 IP3의 생성이 증가함을 볼 수 있었다(그림 11).
Kerovuo 등(2000)에 의하면 Bacillus 유래 phytase는 phytate를 분해하여 IP5, IP4의 isomer형태를 거치면서 최종적으로 IP3가 되는 것으로 확인되었으나, acid phytase는 여러 단계를 거쳐 최종 산 물로 IP1과 5개의 Pi를 생성하는 것으로 알려져 있다. Alkaline phytase(DS11)는 보고된 Bacillus 유 래 phytase와 유사한 반응 패턴을 보이는 것을 확인할 수 있었다. Natuphos 는 wheat phytase와 유 사하게 Ca 유무와는 상관없이 시간이 지남에 따라 phytate는 분해되었고, IP2 혹은 IP와 같은 분 해물들을 생성하는 것으로 확인되었다(그림 12).
Pi Phytate
2mM CaCl2 Control
1 2 3 4 5 6 7 8 9 10 11 12 13 14
그림 10 TLC analysis of phytate hydrolysis reaction catalyzed by alkaline phytase
Pi
Phytate
1 2 3 4 5 6 7 8 9 10 11 12 13 14
2mM CaCl2 Control
Ins P3
Ins P2
그림 11 TLC analysis of phytate hydrolysis reaction catalyzed by wheat phytase (5 µl of the reaction mixture were withdrawn and subjected to TLC.)
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Pi
Phytate
2mM CaCl2 Control
Ins P3
Ins P2
그림 12 TLC analysis of phytate hydrolysis reaction catalyzed by Natuphos (5 µl of the reaction mixture were withdrawn and subjected to TLC.)
3.4 Phytase에 의한 whole wheat flour의 phytate 분해
표준 기질에서의 반응 패턴을 기반으로 실제 식품에서의 기질인 whole wheat flour(통밀가루)에 효소를 처리하고 그 반응 양상을 관찰하였다. 앞에서 효율적인 phytate의 분해를 보여주었던 수 준의 효소를 처리한 경우에는 phytate의 분해를 관찰할 수 없어서, 더 높은 수준의 효소 처리 수 준에서 phytate의 분해 정도를 측정하였다. 50mM Tris buffer(pH 7.0)에 suspend시킨 통밀가루에 alkaline phytase(1.538unit/mg phytate, 20unit/g wheat flour)를 첨가한 후 시간별로 phytate 함량의 변화를 측정하였다.
Monica 등(2001)의 보고에 의하면, 제빵과정에서 밀가루에 자연적으로 존재하는 phytase에 의 하여 일부 phytate가 분해되는 것으로 알려져 있으며, 본 연구에서도 효소를 첨가하지 않은 대조 구에서도 phytate 함량이 유의적으로 감소하고 있는 것을 관찰할 수 있었다. 그러나 효소의 첨가 에 의해 좀 더 많은 phytate가 효율적으로 분해되어 그 함량이 감소하고 있음을 확인할 수 있었 다(그림 13). Alkaline phytase 처리에서 특징적인 현상은 Ca 첨가군이 비첨가군에 비해 phytate 분해 율이 낮게 나타난 것인데, 이것은 밀가루 자체에 함유되어 있는 Ca에 의해 추가의 Ca이 공급될 경 우 효소의 활성이 반대로 제한을 받고 있음을 보여주는 것이라 할 수 있다. 이 점은 이후 효소 처 리의 최적점 설정 시 반드시 고려하여야 할 요인으로 생각된다.
3.5 Phytase 처리에 의한 whole wheat bread의 phytate 함량 변화
Phytic acid를 검출하는 전통적인 방법으로는 Heubner와 Stadler의 absorptiometric 방법(Heubner
& Stadler, 1914)과 phytate의 분리 후 ferric chloride를 이용하여 검출하는 방법(Latta & Eskin, 1980), 그리고 철과 phytate phosphorus의 비율로 측정하는 방법 등이 있지만 낮은 민감도로 인해 정확 한 정량에 어려움이 있는 것으로 알려져 있다(Graf & Dintzis, 1982). 이를 극복하기 위하여 LC 를 이용한 분석법이 개발되었으나, pre 혹은 post column 유도체화, 또는 추가의 전처리 과정으 로 인해 분석법으로서의 단점을 가지고 있다(Skoglund et al, 1997). 이에 비해 최근에 널리 사용되 기 시작한 ion chromatography를 이용한 분석법은 추가의 전처리 과정이 필요 없어 phytate의 분 석에 가장 좋은 방법으로 추천되고 있다. 본 연구에서 사용된 conductivity 검출기를 이용한 ion chromatography 분석법은 Talamond 등(2000)이 보고한 conductivity 검출기를 이용한 분석 방법을 변형하여 사용하였다. 그림 14에 나타난 바와 같이 제빵 과정에서 phytate의 변화를 손쉽게 측정 할 수 있었고, 제빵이 진행되면서 phytate의 함량이 유의적으로 감소하고 있는 것을 확인할 수 있 었다.
Time (min) A
mg of phytate/ml
phytase control
0 20 40 60 80 100 120
0 2 4 6 8 10 12 14 16
Time (min)
phytase (no Ca) phytase (2mM Ca)
control (no Ca) control (2mM Ca) B
Phytate contents(%)
0 20 40 60 80 100
0 120 100 80 60 40 20
그림 13 Hydrolysis of phytate in whole wheat flour by alkaline phytase (1.538 unit/ mg phytate).
Time (min)
µS
µS
Proofed dough Mixed dough Phytate
-2.00.0 5.0
1 2 3
10.0 15.0 20.0 25.0 30.0 35.0 40.0
5.0 10.0 15.0 20.0
min 25.0 Bread
그림 14 Analysis of phytate degradation at different stages of bread preparation by ion chromatography.
완성된 빵에서 발효 시간과 pH 변화에 따른 phytate의 감소 정도를 그림 15에 정리하였다. 원 료 flour에 비하여 30분 발효과정을 거친 alkaline phytase 첨가군은 pH 7에서 42%, pH 5에서 35%
감소하였고, 60분의 발효과정을 거친 경우는 pH 7에서 42%, pH 5에서 40% 감소하여 pH 7에서 유의적으로 많은 감소폭을 보였다. 반면에 acid phytase 첨가군은 원료 flour에 비하여 pH 7에서 37%, pH 5일 때 42% 더 낮은 함량이 측정되었다. pH 7에서 제조된 빵의 경우, acid phytase 처리군 은 발효 시간의 증가에 따라 phytate의 함량이 유의적으로 감소하였으나, alkaline phytase(DS11)는 30분과 60분 발효한 경우에 phytate 함량에 차이가 없는 것으로 측정되었다. 또한 pH 5와 pH 7에 서의 결과를 비교하면, pH 7에서 alkaline phytase를 처리한 경우에 가장 많은 phytate가 분해되는 것을 관찰할 수 있었다. 일반적으로 제빵과정에서는 밀가루에 자체적으로 함유된 phytase에 의해 일부 phytate가 분해되는 것으로 보고되고 있다(Monica et al. 2001). 본 연구에서도 대조구에서 일 부 phytate가 분해되는 것을 관찰할 수 있었으나, 효소 첨가군과 비교할 경우 매우 큰 차이를 보 였다.
A
µg of phytate/g of whole wheat flour
flour 0 25,000
20,000
15,000
10,000
5,000
ug of phytate/g of whole wheat bread
control 30min control 60min DS11 30min DS11 60min Natuphos 30min Natuphos 60min B
pH 5 pH 7
0 25,000
20,000 a
b c d
a b
d d c d 15,000 de e
10,000
5,000
그림 15 Effects of different of phytases on phytate degradation in bread.
(A) total phytate content in whole wheat flour;
(B) µg of phytate/g of whole wheat bread (significant at p<0.05, n = 3).
3.6 Phytase 처리에 의한 whole wheat bread의 minerals 함량 분석
빵에 있는 총 mineral과 유리 minerals 함량을 ICP-OES를 사용하여 측정하고 결과를 표 2에 정리 하였다. Alkaline phytase(DS11) 첨가군(pH 7)에서 유리 minerals 함량은 Pi 860.0±7.2µg, Ca 97.4±
0.9µg, Fe 5.4±0.0µg, Zn 2.4±0.0µg으로 나타나, 대조구의 유리 minerals 함량인 Pi 549.4±26.3µg, Ca 34.4±0.6µg, Fe 0.51±0.1µg, Zn 1.32±0.0µg과 매우 큰 유의적 변화를 보였다. 이들 중 특히 Fe은 효소의 처리에 의해 약 10배 증가하는 큰 변화를 나타내었다. Fe의 변화는 pH 7뿐 아니라, pH 5의 산성 영역에서도 약 8배가량 증가하여, phytate의 분해에 따라 생성된 inositol triphosphate(IP3)의 Fe과의 affinitiy가 가장 크게 변하는 것으로 추측되었다. Acid phytase 첨가군은 효소의 최적 작용 조건인 pH 5에서 유리 minerals 함량은 Pi 1129.7±22.7µg, Ca 65.0±2.0µg, Fe 1.1±0.0µg, Mg 785.4±
12.3µg, Zn 2.75±0.0µg이었고, 역시 대조구에 비해 유의적인 증가 양상을 나타내었다. Acid phytase 의 처리에 의한 mineral의 변화는 전체적인 유리 mineral의 양은 증가하였으나, mineral들 간의 증 감 양상은 대조구와 유사하게 나타났다. 이는 밀가루에 본래 존재하는 wheat phytase와 실험에 사용한 Natuphos 의 기질 분해 패턴이 앞서 기술한 바와 같이 동일하기 때문이고, 반면에 alkaline phytase는 기질의 분해 패턴이 이들과 차이가 나기 때문에, 유리된 mineral의 패턴도 다르게 나타 나는 것으로 생각된다. 대조군의 경우 pH 7보다 pH 5에서 유리 minerals 함량이 높게 나타난 것 은 역시 밀가루에 함유된 wheat phytase의 산성 영역에서의 상대적인 높은 활성 때문으로 생각된 다. 일반적인 wheat phytase의 최적 pH는 5.15이고 pH 6이 넘어가게 되면 급격히 활성이 떨어지기 때문에 pH 7에서 반죽한 빵보다 높은 minerals 함량을 나타낸 것으로 추측되었다(Peers, 1956). 또 한 pH 5에서 acid phytase 첨가군이 alkaline phytase(DS11) 첨가군에 비하여 Pi함량이 높게 나타난 것은 acid phytase의 phytate 분해 최종 생성물이 IP1과 5개의 Pi이나, alkaline phytase의 경우 IP3와 3 개의 Pi를 최종 산물로 생성하기 때문으로 생각되었다.
표 2 Effects of different phytases on the content of free minerals in bread.
Minerals
(µg/g) Total content pH 5 pH 7
Control DS11 Natuphos® Control DS11 Natuphos®
Pi 3359.5±26.5 775.2±10.2d 697.7±2.6e 1129.7±22.7a 549.4±26.3f 860.0±7.2c 892.2±4.0b Ca 382.5±1.52 53.2±2.1d 68.5±1.0b 65.0±2.0c 34.4±0.6f 97.4±0.9a 40.0±0.6e Fe 110.7±1.25 0.52±0.1d 3.86±0.0b 1.1±0.0c 0.51±0.1d 5.4±0.0a 0.5±0.0d Mg 1648.2±11.59 760±7.2b 424.8±3.8f 785.4±12.3a 626.1±4.1d 526.1±3.7e 650.8±4.5c Zn 55.2±0.15 1.95±0.05c 1.67±0.01e 2.75±0.07a 1.32±0.02f 2.4±0.01b 1.82±0.0d Different letters in a row indicate significant difference (p<0.05).
4. 요 약
Alkaline phytase(DS11)를 이용한 식품의 mineral 이용성 증진 가능성을 탐색해 보았다. 제빵과정 에 적용해 본 결과, 기존의 acid phytase에 비해 효율적인 phytate 분해능을 확인할 수 있었고, 또한 alkaline phytase의 작용에 의해서 phytate와 결합하고 있는 다양한 mineral들이 유리될 수 있음을 확인하였다. 이러한 결과는 이후 중성 pH의 식품에서 phytate의 저감화를 통한 mineral 이용성 증 진의 소재로 이용될 수 있음을 시사하는 것으로 alkaline phytase의 식품 산업에의 이용에 기초 자 료로 활용될 수 있을 것으로 기대되었다.
참고문헌
Anne C, Gary W. Purification and characterization of extracellular phytase from Aspergillus niger ATCC 9142.
Bioresource Technol. 86: 183-188 (2003)
Ashok P, George S, Carlos RS, Jose APL, Vanete TS. Production, purification and properties of microbial phytase.
Bioresource Technol. 77: 203-214 (2001)
Bijender S, Satyanarayana T. Characterization of a HAP-phytase from a thermophilic mould Sporotrichum thermophile. Bioresource Technol. 100: 2046-2051 (2009)
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein untilizing the principle of protein-dey binding. Anal. Biochem. 72: 248-254 (1976)
Engelen AJ, Van der HFC, Randsdorp PHG. Simple and Simple and rapid determination of phytase activity. J.
AOAC Int. 77: 760-764 (1994)
Frontela C, Garcia-Alonso FJ, Ros G, Martinez C. Phytic acid and inositol phosphates in raw flour and infant cereals: the effect of processing. J. Food Composition Anal. 21: 343-350 (2008)
Graf E, Dintzis FR. Determination of phytic acid in foods by high-performance liquid chromatography. J. Agr.
Food Chem. 30: 1094-1097 (1982)
Greiner R. Advances in phytase research. Tagung Schweine-und Geflügelernährung, 28~30 (2006) Greiner R, Konietzy U. Phytase for food application. Food Technol. Biotechnol. 44: 125-140 (2006)
Heubner W, Stadler H. Uber eine titrationsmethode zur bestimmung des phytins (Titration method for the determination of phytins). Biochem. Z 64: 422-437 (1914)
Kerovuo J, Rouvinen J, Hatzack F. Analysis of myo-inositol hexakisphosphate hydrolysis by Bacillus phytase:
indication of a novel reaction mechanism. J. Biochem. 352: 623-628 (2000)
Kim YH. Effect of silk peptide on physicochemical properties of bread dough. Korean J. Food Sci. Technol. 36: 246- 254 (2004)
Kim YO, Kim HK, Bae KS, Yu JH, Oh TK. Purification and properties of a thermostable phytase from Bacillus sp.
DS11. Enzyme Microbial. Technol. 22: 2-7 (1998)
Laemmli. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685 (1970)
Latta M, Eskin M. A simple and rapid colorimetric method for phytate determination. J. Agr. Food Chem. 28: 1313-
1315 (1980)
Liu Z, Wang H, Wang XE, Xu H, Gao D, Zhang G, Chen P, Liu D. Effect of wheat pearling on flour phytase activity, phytic acid, Iron, and zinc content. Swiss Society of Food Sci. Technol. 41: 521-527 (2008)
Luis P, Monika H, Markus W, Michel T, Adolphus PGM, van Loon. Gene cloning, Purification, and characterization of a heat-stable phytase from the fungus Aspergillus fumgatus. Appl. Environ. Microbiol. 62: 1696-1700 (1997) Martinez C, Ros G, Periago MJ, Lopez G, Orutuno J, Rincon F. Phytic acid in human nutrition. J. Food Sci.
Technol. 2: 201-209 (1996)
Miksch G, Kleist S, Friehs K, Flaschel E. Overexpression of the phytase from Echerichia coli and its extracellular production in bioreactors. Appl. Microbiol. Biotechnol. 59: 685-694 (2002)
Monica H, Cristina MR, Carmen B. Use of fungal phytase to improve breadmaking performance of whole wheat bread. J. Agr. Food Chem. 49: 5450-5454 (2001)
Nermin B, Adem E, Selman T. Effect of various phytase sources on phytic acid content, mineral extractability and proten digestibility of tarahana. Food Chem. 98: 329-337 (2006)
Nielsen MM, Damstrup ML, Thomsen AD, Raasmussen SK, Hansen A. Phytase activity and degradation of phytic acid during rye bread making. Eur. Food Res. Technol. 225: 173-181 (2007)
Oh BC, Choi WC, S Park, Kim YO, Oh TK. Biochemical properties and substrate specificities of alkaline and histidine acid phytases. Microbiol. Biotechnol. 63: 362-372 (2004)
Oh BC, Chang BS, Park KH, Ha NC, Kim HK, Oh BH, Oh TK. Calcium-dependent catalytic activity of novel phytase from Bacillus amyloliquef aciens DS11. Biochem. 40: 9669-9676 (2001)
Palacios MC, Haros M, Sanz Y, Rosell CM. Selection of lactic acid bacteria with high phytate degrading activity for application in whole wheat breamaking. Swiss Society of Food Sci. Technol. 41: 82-92 (2008)
Peers FG. The phytase of wheat. Biochem. J. 53:102-110 (1953)
Porres JM, Etcheverry P, Miller DD, Lei XG. Phytase and citric acid supplementation in whole-wheat improves phytate-phosphorus release and iron dialyzability. J. Food Sci. 66: 614-619 (2001)
Reddy NR, Sathe SK. Food Phytase. CRC PRESS (2002)
Siekka P. Myoinositol phosphates: analysis, content in foods and effects in nutrition. Lebensm. Wiss. Technol. 30:
633-647 (1997)
Skoglund E, Carlsson NG, Sandberg AS. Determination of isomers of inositol Mono- to Hexaphosphates in selected foods and intestinal contents using High-Performance Ion Chromatography. J. Agric. Food Chem. 45: 431-436 (1997)
Soren KR., Freank H. Identification of two low-phytate barley (Hordeum vulgareL.) grain mutant by TLC and genetic analysis. Hereditas 129: 107-112 (1998)
Talamond P, Doulbeau S, Rochette I, Guyot JP, Trech S. Anion-exchange high performance liquid chromatography with conductivity detection for the analysis of phytic acid in food. J. Chromatogr. A 871: 7-12 (2000)
Tye AJ, Siu FKY, Leung TYC, Lim BL. Molecular cloning and the biochemical characterization of two novel phytases from B. subtilis 168 and B. licheniformis. Appl. Microbiol. Biotechnol. 59: 190-197 (2002)
Wodzinski RJ, Ullah AHJ. Phytase. Advances in Appl. Microbiol. 42: 263-302 (1996)
Xin GL, Jesus MP. Phytase enzymology, applications, and biotechnology. Biotechnol. 25: 1787-1794 (2003)
보건산업백서. 한국보건산업진흥원. (2007)