호알칼리성 Bacillus sp. DK1122 균주가 생산하는 알칼리성 단백질 분해효소의 정제 및 특성
이형재, 유지승, 배동훈*
단국대학교식품공학과
Received: June 14, 2016 / Revised: July 7, 2016 / Accepted: July 14, 2016
서 론
미생물유래의각종효소는일반화학반응대비특이성이 높고, 가격경쟁력이우수하며, 지구환경을보전하는친환 경적생물소재이다.이중 protease의경우, 원시생물체도 자신이 생산한 단백질을 분해, 대사에 재사용하기 위해 protease가필요했을것이므로, protease는생물학적진화과 정중가장초기에존재했을것으로추정된다. 미생물에있
어서 protease의역할은사람이나동물이소화기관인장내
로이효소를분비하는경우와유사하다. 즉, 세포외부의단 백질을세포내로흡수가능한아미노산으로분해하기위해
protease를분비한다. 특히고분자단백질의흡수기작이없
는미생물은 protease 분비를통해열량원및질소원까지얻
을 수 있다[19]. Protease가 작용하는 위치에 따라서
exopeptidase와 endopeptidase로분류[28]하며, 미생물이세 포 외로 분비하거나 동물이 장내로 분비하는 대부분의 protease는 endopeptidase에해당한다[19]. 최근 생물공학 기술의발달과함께식품제조, 제약, 유전자조작등여러방 면에서새로운효소가개발되고있으며, 효소제품의사용도 계속증가하고있다. 그중단백질분해효소(protease)는세 계효소생산의약 25%를차지한산업적으로중요한효소이 다[6, 22].
호알칼리성 protease로는 1958년에알칼리성단백질분해 효소 subtilisin이 Bacillus subtilis로부터 분리되었다[12].
1971년에는호알칼리성세균인 Bacillus sp.로부터알칼리 성단백질분해효소[13]가발견되었고, 이후연육가공, 세 제, 탈모공정등에알칼리성 protease의탁월한유용성에주 목하여, 많은연구자들이활성이높고, 안정된성질을갖는 Purification and Characterization of an Alkaline Protease Produced by Alkalophilic Bacillus sp. DK1122
Hyungjae Lee, Ji-Seung Yoo, and Dong-Hoon Bai*
Department of Food Engineering, Dankook University, Cheonan 31116, Republic of Korea
An alkaline protease was purified and characterized from an alkalophilic microorganism, Bacillus sp.
DK1122, isolated from soil in central Korea. The optimum temperature and pH for the growth of the pro- ducer strain were 40℃ and pH 9.0, respectively. The protease was produced aerobically at 40℃ after 24 h incubation in modified Horikoshi I medium (pH 9.0) containing 0.5% (w/v) glucose, 0.8% (w/v) yeast extract, 0.5% (w/v) polypeptone, 0.1% (w/v) K2HPO4, 0.02% (w/v) MgSO4·7H2O, 1% (w/v) Na2CO3, and 3% (w/v) NaCl.
The alkaline protease was purified by 70% ammonium sulfate precipitation of the culture supernatant of Bacillus sp. DK1122, followed by CM-Sepharose chromatography. The molecular weight of the enzyme was estimated to be 27 kDa on the basis of SDS-PAGE. The optimum temperature and pH for the protease activ- ity were 60℃ and pH 9.0, respectively. Addition of CaCl2 increased the thermal stability of the purified pro- tease, where 90% of protease activity was retained at 60℃ for up to 3 h. Consequently, it is expected that the alkaline protease from this study, exhibiting stability at pH 7–9 and 60℃, may be promising for application in the food and detergent industries.
Keywords: Alkaline protease, alkalophilic, Bacillus, purification, characterization
*Corresponding author
Tel: +82-41-550-3562, Fax: +82-41-559-7868 E-mail: [email protected]
© 2016, The Korean Society for Microbiology and Biotechnology
알칼리성 protease를생산하는균주선별을하기위해많은 노력을해왔다[10, 20, 26]. 또한 Protease의생산[8], 구조분 석[27] 및구조안정화[30], 열안정성[29], 생리학적역할[14], 분자생물학적연구[15]과관련된다양한연구가진행되었 다. 식품, 세제 등 산업적으로 주목을 받는 호알칼리성
protease는알칼리범위의 pH에서최적활성을나타내며대
부분활성부위에 serine 가존재하여, serine과특이반응을 하는물질[31]에의해쉽게불활성화된다고보고되어있다 [1]. 이후최근까지 B. licheniformis MP1 [16], B. horikoshii [17], B. firmus CAS 7 [2], Pseudomonas aeruginosa MN1 [4], Streptomyces clavuligerus strain Mit-1 [33], Aspergillus clavatus [34] 등다양한미생물에서호알칼리성 protease를 분리, 정제및특성연구를통한관련산업으로의응용성을알 아보고자하는연구가많이보고되었다. 현재고온성, 호염 성, 호알칼리성등특수환경미생물자원은기초학문으로서 의생태학적측면은물론산업적으로도새로운효소, 항생 제, 항암제등각종고부가가치생물신소재로응용될수있 다. 이중호알칼리성 protease는식품및세제산업에실제 응용이되고있기에새로운호알칼리성 protease를찾는것 은산업적으로매우중요하다고할수있다.
본연구에서는선행연구를통해토양에서분리, 동정한효 소생산균주로부터호알칼리성 protease의최적생산조건을 수립하고, 생산하여해당효소를정제하고특성을알아보았다.
재료 및 방법
균주 및 배지
선행연구를통해토양에서분리, 동정한균주, Bacillus sp.
DK1122를 protease 생산균주로사용하였다[25]. 생산배지 최적화의기본배양배지는균주분리에사용한 Horikoshi I 배지(0.5% (w/v) polypeptone, 0.5% (w/v) yeast extract, 0.1% (w/v) K2HPO4, 0.02% (w/v) MgSO4· 7H2O, 1% (w/v) Na2CO3, 2% (w/v) agar (pH 10.2)에별도로멸균된 1% (w/v) glucose를혼합제조)를사용하였다[25].
Protease 활성
Protease 활성은 azocasein법을변형하여사용하였다[24].
효소의기질로 azocasein (Sigma, USA)을최종농도 2% (w/v) 로 10 mM sodium phosphate buffer (pH 9.0)에녹인 후, 기질 250 µl에효소액 150 µl를첨가하여 60℃또는지정된 온도에서 10분간 반응시켰다. 이후 600µl의 20% (w/v) trichloroacetic acid를가하여효소를불활성화후, 9,000 × g 에서 10분간원심분리를통해미반응 azocasein은침전, 제 거하였다. 700 µl의 1 N NaOH를원심분리상등액 450 µl에 혼합하고 spectrophotometer (UV-1201, Shimadzu, Japan)
를이용하여 440 nm에서흡광도를측정하였다. 효소활성
도 1 unit은 10분간효소반응에의하여유리되는 azo기에의
해 440 nm에서의흡광도를 0.001 증가시키는양으로정하
였다.
호알칼리성 Protease 생산 배양조건: 온도, 배양시간, pH, 산소공급
균체생육에미치는온도의영향을검토하기위하여 20 ml
의 Horikoshi I 배지에 종배양액을 0.5% 접종한 다음 20, 25, 30, 35, 40, 45, 50℃조건에서 250 rpm으로 24시간진 탕배양한후균체의생육도와배양액내의효소활성을측 정하였다. 배양시간에따른균체의생육과효소생산성을검 토하기위하여 20 ml Horikoshi I 배지에종배양액 0.5% (v/v) 를접종한다음 40℃, 250 rpm으로진탕배양하였으며 4시 간마다배양액을취하여균체의생육도와배양상등액의효 소활성을측정하였다. 균체의생산에미치는 pH의영향을 검토하기위하여 Horikoshi I 배지에서 Na2CO3를제거하여 배지를 조제하였다. 여기에 100 mM sodium phosphate (pH 5−7), Tris-HCl (pH 7−9), sodium carbonate (pH 9− 12) buffer로 pH를조정하여사용하였다. pH를재조정한배 지에종배양액 0.5% (v/v)를접종하여 40℃, 250 rpm에서 24 시간진탕배양하여균체의생육도와효소활성을확인하였 다. 산소공급에따른균체의생육과효소생산성을검토하 기 위하여 250 ml Erlenmeyer flask에 pH 9로 조정한 Horikoshi I 배지를 10−100 ml까지 10 ml 단위로제조하였 다. 각각의배지에종배양액 0.5% (v/v)를접종한다음 40℃,
250 rpm으로 24시간배양하여균체의생육도와효소활성을
측정하였다.
호알칼리성 Protease 생산 배지 최적화: 탄소원, 질소원, NaCl 농도
균체의생육과효소생산에미치는탄소원의종류와농도 에따른영향을검토하기위해 Horikoshi I 배지(pH 9.0)에 1% (w/v) glucose 또는 glucose 대신 maltose, lactose, sucrose, dextrin, xylose, galactose, fructose, starch를 각 1% (w/v) 농도로배지에첨가하여종배양액 0.5% (v/v)를접 종한후 40℃, 250 rpm에서 24시간진탕배양하면서생육 과효소활성을측정하였다. 이중생육과효소활성이가장 좋은탄소원을선별하여 Horikoshi I 배지에 0.5, 1, 1.5, 2,
5, 10% (w/v) 농도가되도록첨가한배지를제조하여사용
하였다. 이후, 각배지에종배양액 0.5% (v/v)를접종하여 40℃, 250 rpm으로 24시간진탕배양하여균체의생육도와 효소활성을측정하였다. 질소원의영향을확인하기위해, 배 지내에질소원으로사용하는 yeast extract와 polypeptone 을제거한후 glucose를 0.5% (w/v) 첨가하여 pH를 9.0으로
조정한 Horikoshi I 배지 20 ml에 yeast extract, polypeptone, bactopeptone, neopeptone, beef extract, tryptone, urea, NH4Cl, (NH4)2HPO4, (NH4)2SO4, NaNO3를각각 1% (w/v) 가되도록첨가하였으며, polypeptone과 yeast extract 혼용 배지의경우는각각 0.5% (w/v) 첨가하여총 1% (w/v)가되 도록액체배지를조제하였다. 각배지에종배양액 0.5% (v/v) 를접종하여 40℃, 250 rpm으로 24시간진탕배양하여배양 액내의생육도와효소활성을측정하였다. NaCl의영향을 검토하기위하여 Horikoshi I 배지에 0.5% (w/v) glucose를 첨가하고 pH 9.0으로조정하여 0, 1, 2, 3, 4, 5, 7, 10, 15, 20% (w/v) NaCl를각각첨가한배지를제조하였다. 여기에 종배양액 0.5% (v/v)를접종하여 40℃, 250 rpm으로 24시간 진탕배양하면서균체의생육도와효소활성을측정하였다.
호알칼리성 protease 정제
Bacillus sp. DK1122의 배양액을 원심분리기(Hanil Supra 22K, Hanil Science Industrial, Korea)로 7500 × g 에서 20분간원심분리하여균체를분리하였다. 4℃에서미 리냉각시켜놓은배양상등액에 ammonium sulfate를 70%
포화용액이되도록첨가한후, 저온에서 30분간교반시키며 효소단백질을침전시켰다. 7500 × g에서 30분간원심분리 후상등액을제거하여침전된효소단백질에서수분을제거 후, 4℃에서냉각된 10 mM sodium-phosphate buffer (pH 6.0)에용해시켜조효소액을준비하였다. 수용성조효소액을 10 mM sodium-phosphate buffer (pH 6.0)로 평형화된 CM-Sepharose column chromatography (35 × 150 mm;
Sigma)를사용하여정제하였다. 이를통해얻은활성효소분
획을 PM-10 membrane을사용하여 Amicon ultrafiltration cell (Millipore, USA)에서 10 mM Tris-HCl buffer (pH 9.0) 로치환하였다.
단백질 정량과 전기영동
단백질의 농도는 UV-1201 spectrophotometer (Shimadzu Co.)를이용하여 Bradford 법[7]에준하여측정하였다. SDS- PAGE는 Laemmli의방법[23]에준하여행하였다. 전기영동 running gel은 15% (w/v)를사용하였고 41 mA에서 3시간 전기영동한 후, 0.05% (w/v) Coomassie brilliant blue R-
250 (Sigma)으로염색후탈색하여단백질가수분해효소단
백질의정제정도를확인하였다.
호알칼리성 protease 최적반응 조건: 반응온도, 열안정성, pH, 무기염류
효소반응의최적온도를정하기위해효소를 30−70℃범 위에서 10℃단위로 10분간효소반응을하여최적반응온도 를정하였다. 효소의열안정성을검토하기위하여효소를 30−
70℃범위에서 10℃단위마다 0−5시간동안 1시간마다효소 활성을측정하였다. 효소의활성에미치는반응용액의 pH 영향을확인하기위하여 sodium phosphate buffer (pH 5−7), Tris-HCl buffer (pH 7−9), sodium carbonate buffer (pH 9−11)를 10 mM 농도가되도록첨가하여 60℃에서 10분간 반응시킨후, 잔존활성을측정하였다. 무기염류가효소의열 안정성에미치는영향을확인하기위하여열에불안정한 60℃ 에서효소에 CaCl2나 MgCl2를첨가하여활성을측정하였 다. 추가로 CaCl2를 2, 10 mM의농도로첨가하여 60℃에서 0−5시간반응시키며 1시간마다효소활성을측정하였다.
결과 및 고찰
호알칼리성 protease 생산조건 최적화: 온도, 배양시간, pH, 산소공급
Protease 생산 시, 배양온도의 영향을 확인하기 위해,
Horikoshi I 배지를사용하여종배양액을 0.5%씩접종한후 배양온도를 20−50℃로각기달리하여 250 rpm, 24시간동 안진탕배양후균체생육과효소활성을측정한결과, 균체 생육은 25℃에서서서히증가하여 40℃에서최대를보였으 며 45℃이상에서는생육이이루어지지않았다(Fig. 1A). 균 체로부터의단백질분해효소활성도 40℃에서최대의활성 을보였다. 본균주는 30−40℃의좁은온도범위에서만생 육하며, 배양액중효소활성은 40℃에서만최적의활성을보 이고이외의온도에서는매우낮은활성을보이므로이후의 실험에는배양온도를 40℃로하였다.
배양 시간에 따른 효소 생산을 검토하기 위하여 40℃,
250 rpm으로진탕배양후배양액을 4시간간격으로채취하
여균체의생육도와효소활성을측정하였다. Fig. 1B와같이 균체의생육이 4시간이후부터서서히증가하기시작하여 24 시간이후에최대의생육을보였고, 이후에서서히감소하였 으나그차이가현저하게나타나지는않았다. 또한효소활성 은 20시간과 24시간에서가장높았으며그이후효소의활 성이최대치의약 80%를유지하는 것으로확인되었다. 따 라서이후의실험에서는종배양액을접종하여균체생육과효 소활성이최대치를갖는 24시간에서배양액을회수하였다.
균체생육과효소생산에미치는 pH의영향을검토하기 위하여배양액에서균체생육과효소활성을측정한결과는 Fig. 1C와같다. 균체는 pH 7 이상에서생육하여 pH 9에서
최대생육을보였으며 pH 11 이상에서균체생육이급격히
저하됨을확인하였다. 효소활성은 pH 8 이하와 pH 10 이상
에서현저히낮았으며, pH 9에서최대활성을나타내었다.
또한 pH 9에는 Tris-HCl buffer를첨가하여배양한균주와 sodium carbonate buffer를첨가하여배양한균주의생육도 는비슷하였다. 반면, 효소활성에있어서는 sodium carbonate
buffer를 사용한 배지에서의 균체 효소활성이 Tris-HCl
buffer를첨가하여배양한균체의효소활성보다 20% 정도
높은 효소활성을 나타내었다. 따라서 이후의 실험에서는 sodium carbonate를첨가한 Horikoshi I 배지를 pH 9로재 조정하여사용하였다.
효소생산에미치는산소공급영향을검토한결과균체 의생육도는산소의공급량에크게영향을받지않고생육 을보였으나, 효소의활성에는산소공급조건에따라급격 한차이를나타내었다(Fig. 1D). 배지의양이 20 ml일때균
체의효소활성이최대를보였으며배지의양이 20 ml보다
많아질수록균체의효소활성은급격히감소되었다. 따라서 Bacillus sp. DK1122가생산하는 protease는산소공급에따 라효소활성에큰영향을미치는것으로확인되었다. 호알칼리성 protease 생산 배지 최적화: 탄소원, 질소원, NaCl 농도
효소생산에미치는탄소원의영향을검토하기위하여배 양후균체의생육도와효소활성을측정한결과 glucose를 첨가하였을때최대의생육도와효소활성을보였으나, 다른 탄소원에서는효소활성이거의없었다(data not shown). 한
편최대의활성을보인 glucose의농도변화에따른효소의
생산성을 검토하기 위하여 Horikoshi I 배지에 0, 0.5, 1, 1.5, 2, 5, 10%의 농도의 glucose를첨가하여 배양한결과
0.5% glucose농도에서가장높은활성을나타내었다. 따라
서이후의실험에서는 0.5% 농도의 glucose를첨가한배지 를사용하였다.
균체생육과효소생산에미치는질소원의영향을검토한 결과 polypeptone과 yeast extract를혼합하여첨가한배지 에서다른질소원을첨가했을때보다높은 생육과효소활 성을 보였다(data not shown). 각각 yeast extract와
polypeptone의농도를변화시키면서효소생산을검토한결
과 yeast extract는 0.8%의농도에서가장높은활성을나타 냈으며, polypeptone은 0.5% 농도에서최대의효소활성을보 였다(data not shown).
본균주에있어서 NaCl 농도가균체생육과효소활성에
미치는영향을검토한결과생육도는 10% NaCl 농도까지
높은생육을보였으나 15% 이상에서급격히감소하기시작
하여그이상의고농도염을첨가하면균체가자라지못하 는것으로 확인되었다(data not shown). 또한효소활성은 3%의 NaCl 농도에서최대의활성을보였으며, 그이상의농 도에서는균체의생육이높더라도활성이급격히감소하여 7% 이상의농도에서는활성을거의찾아볼수없었다. 따라 Fig. 1. Effect of culture temperature (A), culture time (B), and initial pH (C) on the growth and enzyme production from Bacillus sp. DK1122. (D) Effect of medium volume on the enzyme production from Bacillus sp. DK1122.
서본균주는 3% 농도에서최대의활성을갖으며 10%의염 농도에서도생육하는내염균(halotolerant)으로판단되었다. 이전연구에서 5% NaCl에서최적성장을나타내는 Bacillus sp. 균주가생산하는호알칼리성 protease에대한연구가있 었다[18].
위의실험결과를토대로한 Bacillus sp. DK1122의최적 화된배양조건은균주스크리닝에사용한 Horikoshi I 배지 와 차이가 있었다. 최적배지를 modified Horikoshi I medium로 명명하였고, 조성은 0.5% (w/v) glucose, 0.8%
(w/v) yease extract, 0.5% (w/v) polypeptone, 0.1% (w/v) K2HPO4, 0.02% (w/v) MgSO4· 7H2O, 1% (w/v) Na2CO3, 3% (w/v) NaCl이고 pH 9.0으로조정하였다. 이후의실험에 서는 modified Horikoshi I medium으로 Bacillus sp.
DK1122를배양하였다.
호알칼리성 protease 정제 및 특성
Bacillus sp. DK1122가생산하는알칼리성단백질분해효 소를분리, 정제하기위하여효소최적생산조건에서균체를 배양하였다. 배양후균체를원심분리시켜회수한배양상등
액으로부터 효소 단백질을 침전하기 위하여 ammonium
sulfate 농도를달리하여효소침전조건을검토하였다. 균
체를제거한상등액에 ammonium sulfate의포화농도를각
각 50, 70, 80%로하여가하고, 4℃에서 4시간방치하였다. 침전된 효소 단백질을 10 mM sodium-phosphate buffer
(pH 6.0)에용해시켜효소활성을측정하였으며, 효소량을구
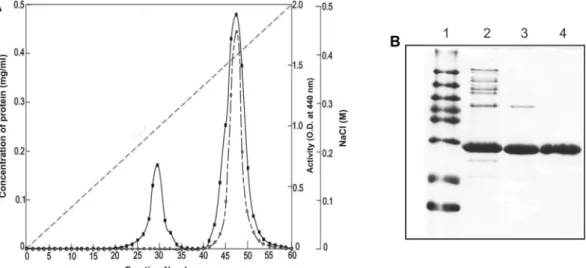
하여배양상등액의효소량과비교하여회수율을계산하였 다. 그결과 70%의포화농도에서가장높은효소침전회수 율(41.8%)을보였다(data not shown). Ammonium sulfate 에의하여침전시킨효소단백질을 10 mM sodium phosphate buffer (pH 6.0)에 현탁시킨 후 CM-Sepharose와 DEAE- sepharose에흡착시켜본결과효소활성이 DEAE-Sepharose의 경우비흡착부분에서효소활성을보였고, CM-Sepharose의 경우에는흡착된부분에서효소활성을보였다. 따라서효소 액을 10 mM sodium phosphate buffer (pH 6.0) 조건에서 CM-Sepharose에흡착시키고 0−0.5 M NaCl 범위에서농도 별로증가시키며효소를용출한결과효소활성은 No. 41−52 분획관에서용출되었다(Fig. 2A). CM-Sepharose column에 의해서는배양상등액보다 2.8배의정제도를보였으며회수
Fig. 2. Purification of alkaline protease from Bacillus sp. DK1122. (A) Chromatogram of an alkaline protease from Bacillus sp. DK1122 on CM-Sepharose chromatography. ---: concentration of NaCl (M);■: concentration of protein (mg/ml); ○: enzyme activity (optical den- sity at 440 nm). (B) SDS-PAGE of an alkaline protease from Bacillus sp. DK1122. Lane 1, molecular weight markers of 97.4, 66.2, 55.0, 42.7, 40.0, 31.0, 21.5, and 14.4 kDa; lane 2, culture broth; lane 3, precipitant of 70% ammonium sulfate; lane 4, active fractions of CM-Sep- harose chromatography.
Table 1. Purification table of protease from Bacillus sp. DK1122.
Step Total protein
(mg)
Total activity (unit)
Specific activity (unit/mg)
Yield
(%) Fold
Culture broth 7,725.0 1,427,100 184.7 100 1
70% Ammonium sulfate 1,401.6 596,300 425.4 41.8 2.3
CM-Sepharose chromatography 659.8 341,700 517.9 23.9 2.8
율은 23.9%였다(Table 1). 정제효소의순도를확인하기위 하여 SDS-PAGE를행하였다. 그결과정제된단일 band를 확인하였고, 분자량은 27 kDa로추정되었다(Fig. 2B). 다른 Bacillus spp.에서생산하는호알칼리성 protease의분자량 이 30 kDa [5], 28 kDa [9, 21]인것과 유사한것으로판명 되었다. 그러나 halotolerant Bacillus sp.에서유래호알칼 리성 protease (40 kDa)와는분자량이상이하였다[18]. SDS- PAGE 분석에의하여 CM-Sepharose column chromatography 에의한정제분획물로부터단일단백질밴드를나타내어순 수정제된것으로판단되었다. 일부 Bacillus spp. 경우 2개 의 monomeric 단백질로호알칼리성 protease를생산한다고
보고된것과는상이한결과를나타냈다[11, 32].
호알칼리성 protease 최적반응 조건: 반응온도, 열안정성, pH, 무기염류
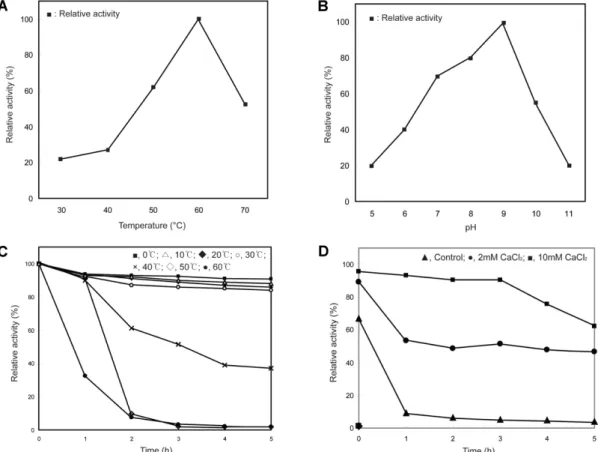
효소반응온도를 30−70℃범위로온도를조정하여정제 된효소활성최적반응온도를측정하였다. 그결과 40℃부 터효소활성이점차증가하여 60℃에서최대효소활성을보 였으며그이상의온도에서는급격히감소하였다(Fig. 3A).
따라서효소활성측정시반응온도를 60℃에서행하였다.
다른 Bacillus spp.에서분리된효소의경우최적활성온도
는 60℃ [9, 11]로동일하거나, 40℃ [32]로본연구에서정 제한효소의최적활성온도보다낮았다.
효소의활성에미치는반응용액의 pH 영향을확인하기위 하여 sodium phosphate buffer (pH 5−7), Tris-HCl buffer (pH 7−9), sodium carbonate buffer (pH 9−11)를 10 mM 농도가되도록첨가하여 60℃에서 10분간반응시켰다. 그결 과효소활성이 pH 7−9의범위에서높게나타났으며 pH 9.0 에서최적의효소활성이나타남을확인하여정제된효소는 alkaline protease로판명되었다(Fig. 3B). 기존보고된호
알칼리성 protease의경우와동일하거나유사한 최적 pH
(pH 8.0−10.0)를나타냈다[3, 11, 34]. 따라서효소활성측 정시기질로사용하는 azocasein 용액은 pH 9.0으로조정 하여사용하였다. 이를통해세제산업등알칼리성에서활
성을나타내는 protease 수요를충족시키는데본효소나이
를기반으로추후개량된효소를활용할수있을것으로판 단된다.
정제된효소의열안정성을검토하기위하여효소를 0−60℃ 까지 0−5시간동안반응시키며 1시간마다효소활성을측정 하였다. Fig. 3C에서나타난바와같이 0−30℃까지는 5시간
Fig. 3. Effect of temperature (A), pH (B), and inorganic salts (D) on the activity of the purified protease from Bacillus sp. DK1122.
(C) Thermal stability of the activity of the purified protease from Bacillus sp. DK1122.