된장 분리균 Bacillus sp. YB-1401의 Mannanase 생산성과 효소특성
전호진, 윤기홍*
우송대학교바이오식품과학전공
Received: March 17, 2014 / Revised: April 8, 2014 / Accepted: April 14, 2014
서 론
Mannan
다당류는mannose, glucose
와galactose
의결합 으로 이루어져 있는데,
구성하고 있는 당의 종류에 따라mannan, glucomannan, galactomannan
과galactogluco- mannan
으로구분된다.
이들의직쇄골격은mannose
잔기끼 리또는mannose
와glucose
잔기간에β-1,4
결합으로이루 어지며galactose
는mannose
의6
번탄소와 α-1,6
결합을하 여측쇄로존재한다.
β
-1,4-Endo-mannanase (mannanase)
는β-mannan
다당 류에서mannose
간의β-1,4-mannosyl
결합을무작위적으로 분해하여mannooligosaccharides (MOS)
로전환시키며,
이 것은 β-1,4-mannosidase
에 의해mannose
로 분해되는데,
mannan
을mannobiose
단위로절단하는exo-mannanase
의 존재도Bacteroides fragilis
로부터 보고되었다[7].
그리고mannose
와 결합하여 측쇄로 존재하는galactose
는 α- galactosidase
에의해분해된다. Mannan
의분해에가장중요한역할을하는
mannanase
는동물과식물을비롯한미생물에서두루발견되었으며펄프와제지표백공정
,
바이오매 스자원의당화공정,
커피가공공정을비롯하여세제와사료첨가용효소로산업적활용성이높다
[2].
특히곰팡이와세균유래의
mannanases
는그특성이다양하므로산업적으로활용하기적합한성질을갖는효소를개발하기위해산 성
,
알칼리성,
내열성 또는 호냉성 효소가 탐색되거나[5, 13, 19, 26],
변이 효소로 제조되었으며[22], Ca
2+ 이온이Clostridium thermocellum [3], Streptomyces thermolilacinus
과Thermobifida fusca [9]
유래의mannanases
의내열성을증 가시킨다는보고도있다.
알칼리성효소는세제나펄프제지 산업에서,
산성효소는식품과사료산업에서그가치가높으 며내열성은공통적으로요구되는특성이다.
미생물에서생Production and Characterization of Mannanase from a Bacillus sp. YB-1401 Isolated from Fermented Soybean Paste Ho Jin Joen and Ki-Hong Yoon*
Food Science & Biotechnology Major, Woosong University, Daejeon 300-718, Republic of Korea
A Bacillus strain capable of hydrolyzing locust bean gum was isolated as a producer of extracellular mannanase by way of an enrichment culture in an acidic medium from homemade soybean pastes. The isolate YB-1401 showed a biochemical identity of 61.1% with Brevibacillus laterosporus, while the nucleotide sequence of its 16S rDNA had the highest similarity with that of Bacillus amyloliquefaciens. The mannanase productivity of the Bacillus sp. YB-1401 was drastically increased by mannans.
Particularly, maximum mannanase productivity was reached at approximately 265 U/ml in LB medium supplemented with konjac glucomannan (4.0%). The mannanase was the most active at 55
oC and pH 5.5. Mannanase activity was completely maintained after pre-incubation at pH 3.5 to 11.0 for 1 h. The predominant products resulting from the mannanase hydrolysis were mannobiose and mannotriose for LBG, guar gum or mannooligosaccharides. A small amount of mannose was also detected in the hydrolyzates.
Keywords: Bacillus sp. enrichment culture, mannanase, productivity, property
*Corresponding author
Tel: +82-42-630-9742, Fax: +82-42-636-2676 E-mail: [email protected]
© 2014, The Korean Society for Microbiology and Biotechnology
산되는
mannanase
는대부분균체외로분비되며, glycosyl hydrolase (GH) family 5
와26
에 속한다. GH26
에는 주로세균성효소가대부분을차지하고
, GH5
에는곰팡이유래또는세균유래효소가포함된것으로알려져있다
.
한편균체 내효소이며GH113
에속하는mannanase
가Alicyclobacillus acidocaldarius
로부터보고되었다[25].
최근에는
MOS
가장내유익한미생물의성장을촉진하고 장관의 치료효과를 보이는 것으로 알려지면서[1, 17, 24]
glucomannan
이나galactomannan
을mannanase
로분해하 여MOS
또는glucomannooligosaccharides
를 생산하고자 하는연구도활발히진행되고있다[1, 3, 17].
Galactomannan
을함유한콩의발효식품은우리나라를비롯한대부분의아시아국가뿐아니라아프리카지역에서도 고유식품으로 이용되고 있으며
,
이로부터 여러 종류의Bacillus
속미생물이비전분성다당류의분해효소를생산하는균주로분리되고그효소의특성이연구되었다
[10, 15,
21].
본연구에서는산성조건에서된장발효미생물을증균하여이로부터
mannanase
를생산하는Bacillus
속균주를분 리하고효소생산성과특성을조사하였다.
재료 및 방법
Mannanase를 생산하는 내산성 균주의 탐색
가정에서제조된된장
25
점을수집하여pH
를4.5
로조절 한LB
배지에접종하고37
oC
에서2
일간진탕배양한후동 일한배지에3
일간격으로3
회계대배양을추가적으로실 시하였다. pH
가4.5
로조절된LB
평판배지(yeast extract, 5 g; tryptone, 10 g; NaCl, 5 g; agar, 15 g; water, 1 liter)
에계대배양액을도말하여37
oC
에서2
일간배양한후형성 된콜로니를0.5% locust bean gum (LBG)
이첨가된중성의LB
평판배지에접종하여하룻밤배양하고congo red
용액(0.2%)
으로 염색하여LBG
분해환을 생성하는 미생물을mannanase
생산균으로선발하였다.
분리균주의 동정분리균의형태적관찰을위해서는그람염색과포자염색을 실시하였으며
,
배양된균체현탁액을제조사의지침에따라API 20E
와API 50 CHB kits (Bioméreux
사, France)
에접 종하고37
oC
에서배양하면서1
일과2
일째각각관찰하여탄 수화물이용능과생화학적특성을판별하였다.
분리균의16S
rRNA
유전자염기서열을분석하기위해서분리균의총염색체
DNA
를주형으로하고,
세균의16S rRNA
유전자의보 존적지역의염기서열을primers
로사용하여중합효소연쇄반응
(PCR)
을실시함으로써얻은PCR
산물을정제하여그염기서열을결정하였다
[10].
Mannanase 조효소액 제조
Bacillus
속분리균을LB
배지에서전배양한후4.0%
곤약 이첨가된LB
배지에1% (v/v)
가되도록접종하여24
시간동 안진탕배양하였다.
배양액을원심분리하여얻은배양상등 액을ammonium sulfate (30-75%)
로 처리한 후, 12,000 rpm
에서30
분동안원심분리함으로써상등액을제거하였 다.
단백질침전물을10 mM citrate
완충용액(pH 5.5)
에현 탁하여동일한완충용액으로투석한후이를원심분리하고상등액을취해
mannanase
조효소액으로사용하였다.
Mannanase 활성 측정과 반응특성 분석
Mannanase
활성은0.5% LBG
와50 mM sodium citrate
완충액(pH 5.5)
을포함한반응액에효소를첨가하여50
oC
에서
15
분간 반응을 실시한 후 유리된 환원당을3,5-
dinitrosalicylic acid
방법[14]
으로측정하였으며,
효소활성 도1.0 unit
는위의조건에서1
분동안LBG
로부터1
μmol
의
mannose
에상응하는환원당을생성하는효소의양으로정의하였다
. Mannanase
활성에미치는반응온도와pH
의 영향을조사하기위하여30-65
oC
와pH 4.0-8.0
의범위에서 각각효소활성을측정하였다.
효소내열성은30-60
oC
에서mannanase
조효소액을1
시간동안방치한후잔존활성을측정하여결정하였고
, pH
안정성을조사하기위해서는pH 3.0-12.0
범위의완충용액을최종농도가50 mM
이되도록조 효소액과혼합하여4
oC
에서1
시간동안방치한후mannanase
잔존활성을측정하였다.
이때pH
를조절하기위해서citrate
완충용액(pH 3.0-6.0), phosphate
완충용액(pH 6.0-8.0), Tris
완충용액(pH 8.0-9.0), KCl-borate
완충용액(pH 9.0- 10.0), carbonate
완충용액(pH 10.0-11.0)
과phosphate/NaOH
완충용액(pH 11.0-12.0)
이각각사용되었다. Mannanase
에의한가수분해산물을분석하기위해서는
0.5%
함량의LBG,
guar gum
또는중합도가2-6
인MOS
를각각기질로사용하 였으며,
이들기질과20 mM citrate
완충액(pH 5.5)
을포함 한반응액에과량의효소를첨가하여45
oC
에서5
시간반응 시킨후95
oC
에서5
분간열처리하고원심분리한다음상등 액을적정량취해chloroform, acetic acid
와증류수(4.3 : 5 : 0.7, (v/v))
혼합용액을전개용액으로하여silica gel-precoated thin layer plate (Merck, Kiesegel, No. 5748)
에서 박층크 로마토그래피를수행하였다.
전개된물질을발색시키기위 해서는ethanol 9 ml, p-anisaldehyde 0.5 ml, sulfuric acid 0.5 ml
와glacial acetic acid
몇방울을혼합한발색제용액 을뿌린후, 120
oC
에서10
분간방치하였다.
활성염색
조효소액을
SDS-PAGE
한 후polyacrylamide gel
에서SDS
를제거하기위하여25% isopropanol
를포함한50 mM
phosphate
완충액(pH 6.0)
으로세척한후동일완충액에15- 30
분간방치하였다. Polyacrylamide gel
을꺼내어동일한완 충용액에0.2% LBG
와1.5% agar
를녹여제조한gel
을중층 하여비닐로봉입하고50
oC
에서4
시간반응시킨후중층한gel
을congo red
로염색하여분해된부분을관찰하였으며polyacrylamide gel
은Coomassie brilliant blue
용액으로 염색하여단백질을관찰하였다.
결과 및 고찰
증균 배양을 이용한 mannanase 생산균의 탐색과 분리균 의 특성
된장으로부터
mannanase
를생산하는내산성Bacillus
속 균주를분리하기위하여pH
를4.5
로조절한LB
액체배지에 충청도지역의가정에서각각제조된25
점의된장시료를각 각접종하여진탕배양한후추가로3
회계대배양을실시 하였다.
그결과마지막계대배양액중에서오직1
점의시 료에서만균의성장을나타냈다.
이를LB
평판배지(pH 4.5)
에도말하고37
oC
에서2
일간배양함으로써2
종류의콜로니 가분리되었는데,
이로보아된장에존재하는내산성세균의 종류가적은것으로판단되었으며이로인하여된장시료25
점중오직1
점시료의접종액에서만균의성장을보였던 것으로추측된다.
한편분리된2
개의균주중YB-1401
로명 명한한개의균주가LBG
를분해하는균주로판명되어이 를mannanase
생산균으로선발하였다.
분리균
YB-1401
의형태적특성을조사한결과포자를형성하는그람양성간균으로확인되어
API 50 CHB
와20E
kit
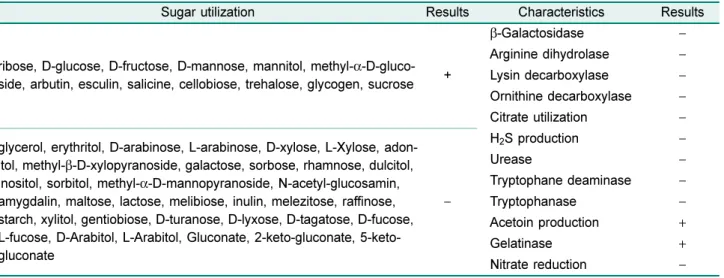
를사용하여생화학적특성을조사하였다. Table 1
에보 인바와같은YB-1401
의생화학적특성을Biomériux
의API
web (https://apiweb.biomerieux.com/jsp/login.jsp)
에서다른균 주와비교한결과Brevibacillus laterosporus
와유사도(61.1%)
가가장높았으며, Bacillus subtilis
및B. amyloliquefaciens
와 는26.9%, B. pumilus
와는11%
의유사도를보였다.
또한분리균
YB-1401
의16S rRNA
유전자를PCR
로증폭 한 후1,496 bp
크기의 염기서열을 결정하고(GenBank accession no. KJ57222)
이를미국NCBI
의BLAST
검색방 법으로16S rDNA database
와 비교하였다.
그 결과B.
amyloliquefaciens FZB42 (NR_075005)
와 가장 유사도가 높았으며 비교된1,493
개 염기중1
개 염기가 달랐고B.
subtilis 168 (NR_102783)
과는4
개염기가다른것으로확인 되었다.
그러나분리균과생화학적특성의유사도가가장높 았던B. laterosporus
의16S rDNA
염기서열(AB112720)
과는
89%
수준의낮은유사도를보였다.
그러므로현재의결과로는분리균이
Bacillus
속의어느균종에속한다고판단할 수 없다고 하겠다
.
한편 된장이나 청국장으로부터B.
licheniformis
가β-glucanase [4]
와 α-amylase [11]
의생산 균으로분리되었으며, mannanase
를생산하는B. subtilis
도 분리된 바 있으나[10, 15], mannanase
를 생산하는B.
laterosporus
또는B. amyloliqueaciens
는보고되지않았다. YB-1401
은된장을pH 4.5
의조건에서증균한후분리된 것이므로산성조건에서균의성장도를조사하기위해pH
가4.0
과4.5
로조절된LB
액체배지에서16
시간동안플라스 크배양을하면서배양액의흡광도를조사하였다.
그결과YB-1401
은pH 4.0
배지에서는전혀성장하지못하였고pH 4.5
배지에서는배양초기에균의성장이매우느렸지만배양 시간이10
시간정도에이르렀을때부터빠른성장을보였다.
한편된장에서B. subtilis
에속하는균주가mannanase
생 산균으로분리된바가있어[10, 15] B. subtilis
중실험균으Table 1. Characteristics of the isolate YB-1401.
Sugar utilization Results Characteristics Results
ribose, D-glucose, D-fructose, D-mannose, mannitol, methyl- α-D-gluco- side, arbutin, esculin, salicine, cellobiose, trehalose, glycogen, sucrose +
β-Galactosidase −
Arginine dihydrolase − Lysin decarboxylase − Ornithine decarboxylase −
Citrate utilization −
glycerol, erythritol, D-arabinose, L-arabinose, D-xylose, L-Xylose, adon- itol, methyl- β-D-xylopyranoside, galactose, sorbose, rhamnose, dulcitol, inositol, sorbitol, methyl- α-D-mannopyranoside, N-acetyl-glucosamin, amygdalin, maltose, lactose, melibiose, inulin, melezitose, raffinose, starch, xylitol, gentiobiose, D-turanose, D-lyxose, D-tagatose, D-fucose, L-fucose, D-Arabitol, L-Arabitol, Gluconate, 2-keto-gluconate, 5-keto- gluconate
−
H
2S production −
Urease −
Tryptophane deaminase −
Tryptophanase −
Acetoin production +
Gelatinase +
Nitrate reduction −
로널리사용되고있는
B. subtilis 168
를pH 4.5
배지에서배양한결과전혀성장하지못하여분리균
YB-1401
과는큰차이를나타냈다
(
결과미제시).
부가탄소원에 따른 mannanase 생산성
곰팡이와세균유래의
mannanase
는mannan
다당류를함 유한물질에의해그생산성이크게증가하는경우가많다. B. subtilis BS5 [5], B. subtilis WL-7 [10]
과B. subtilis WY34 [6]
는 곤약, Aspergillus niger BCC4525 [16]
와B.
halodurans PPKS-2 [19]
는copra meal
에의해mannanase
생 산성이증가하는것으로보고되었다.
그러므로부가탄소원 이Bacillus sp. YB-1401
의mannanase
생산성에미치는영 향을조사하기위해여러종류의탄수화물이0.5% (w/v)
가 되도록각각첨가된LB
배지에서배양온도를37
oC
로하여24
시간 동안 진탕 배양한 후 배양상등액에 존재하는mannanase
활성을조사하였다.
그결과곤약이첨가된배지에서효소생산성이가장높았으며
,
이외에도mannan
다당류를함유한
LBG
와guar gum
에의해서도그생산성이크 게증가하였다.
그러나mannan
다당류를함유한농산부산 물인palm kernel meal
에의한효소생산량의증가정도는높지않은것으로나타났다
(Table 2).
또한곤약의첨가량을달리하였을때그함량이높을수록효소생산량이크게증 가하여
4%
곤약을첨가하였을때최대생산성을보였다.
이는곤약을첨가한
LB
배지에서곤약의함량이0.5%
일때mannanase
생산성이최대이고그이상의함량을첨가해도mannanase
생산성이 더 이상 증가하지 않은B. subtilis WL-7
에비해크게다른점이라고할수있다[10]. B. subtilis WY34
도2%
곤약을첨가한배지에서mannanase
생산성이 가장높은것으로알려졌다[6].
실제곤약이첨가된LB
배지는점도가높아지고특히
2%
이상을첨가하였을때는배양 초기에액체배지가제대로진탕이되지않았으며, 4%
곤약을첨가한배지는
agar
를첨가하지않았지만거의고형의상태였다
.
그러나 배양이 진행되면서 분리균이 생산하는mannanase
에의해서곤약이분해됨으로써배양중기부터는배지의점질도가급격하게저하되고
24
시간배양후에는 배양액에점질도가없었다.
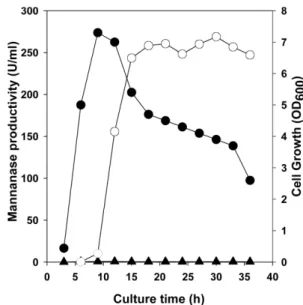
배양시간에따른
mannanase
생산성을조사하기위해곤 약(4%)
을첨가하거나첨가하지않은LB
배지에서36
시간동 안배양하면서mannanase
생산성과성장도를조사한결과,
곤약을첨가하지않은LB
배지에서9
시간배양후에균의 성장이최대에이르렀으나효소생산성은1 U/ml
미만으로 매우낮았다.
곤약을첨가한배지에서는배양시간이9
시간 이 되었을때mannanase
생산성이10.2 U/ml
이었으나, 12
시간에서는155.7 U/ml
로급격하게증가되었고18
시간이후에는약
265 U/ml
수준의최대생산성을보였으며36
시간까지그생산성이유지되었다
(Fig. 1).
한편LB
배지에 서측정한배양균액의흡광도가12
시간이후부터는지속적 으로감소하는것으로보아사멸기에이른것으로판단되었 는데,
곤약을첨가한배지에서는배지의점도가너무높아 배양초기에균이원활하게성장하지못하였을것을감안하 면사멸기에이르는시간이LB
배지에서보다오래걸릴것Table 2. Effects of additional carbon sources on the mannanase production of Bacillus sp. YB-1401.
Additional carbon sources
(0.5%)
Mannanase productivity
(U/ml)
Additional amount (%)
of konjac glucomannan
Mannanase productivity
(U/ml)
None 0.2 0.3 11.3
Glucose 0.3 0.5 12.1
Xylose 0.7 1.0 16.1
α
-Cellulose 0.9 1.5 20.1
Locust bean gum 8.4 2.0 36.9
Guar gum 4.6 2.5 41.4
Konjac glucomannan 12.2 3.0 113.0
Palm kernel meal 1.3 3.5 128.7
Wheat bran 0.4 4.0 217.8
Fig. 1. Production of mannanase by Bacillus sp. YB-1401 and its growth. The isolate was grown respectively in LB broth sup- plemented with or without konjac glucomannan (4%) for mannan- ase production. Cell growth (closed circles) and mannanase productivity (closed triangles) were observed in the media without additional carbon source. Culture medium containing konjac glu- comannan was used for mannanase production (open circles).
Enzyme activities in the culture filtrates were determined in the
reaction condition of pH 5.5 and 50
oC. Each curve represents the
average of three independent experiments.
으로여겨졌다
.
배양상등액을
ammonium sulfate
로처리하여제조한조효소액을사용하여
SDS-PAGE
를수행한후활성염색을실시한결과
Fig. 2
에보인바와같이분자량이약38.0 kDa
정도로추정되는
mannanase
활성단백질이관찰되었으며이는
SDS-PAGE
에서38-40 kDa
의분자량을보인B. subtilis WY34 [6], B. subtilis BCC41051 [18]
과B. subtilis WL-3 [15]
의mannanases
와유사하다고하겠다.
Mannanase의 반응특성
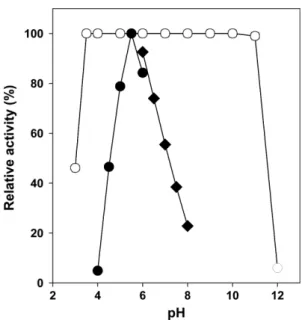
조효소액을사용하여반응
pH
와온도가효소활성에미치 는영향을조사하기위해반응온도를50
oC
로고정하고여러pH
에서mannanase
활성을측정한결과pH 5.5
에서최대 활성을보였으며pH 5.0-6.5
범위에서최대활성의70%
이 상의활성을나타내었다(Fig. 3).
또한반응액의pH
를5.5
로 하고반응온도에따른효소의활성을조사한결과55
oC
에 서활성이가장높게나타났다(Fig. 4).
따라서YB-1401
의mannanase
는반응온도50-70
oC,
반응pH 5.0-7.0
의범위에 서 최대활성을 보이는 대부분의Bacillus
속 균주 유래mannanases [10, 15, 18, 26]
와유사한것을알수있다.
반 면B. subtilis BS5 [5], B. subtilis G1 [20]
과B. halodurans PPKS-2 [19]
로부터유래된mannanases
는최적반응온도가 낮거나알칼리조건에서최대활성을보이는것으로각각보 고되었다.
pH 3-12
의범위에서mannanase
의안정성을조사한결과pH 3.5-10
에서1
시간동안방치하였을때실활되지않았으Fig. 2. Zymogram (A) and SDS-PAGE (B) of the crude enzyme produced by Bacillus sp. YB-1402. After SDS-PAGE of the con- centrated culture filtrate of isolate YB-1401 grown in LB broth con- taining konjac glucomannan (4%), protein exhibiting mannanase activity, which was indicated with arrow, was analyzed by activity staining with LBG as substrates. Molecular size is shown in kilo- daltons to the right side of the gel.
Fig. 3. pH Optimum and pH stability of the crude enzyme.
Buffers (50 mM) used were as follows: sodium citrate (pH 3-6), sodium phosphate (pH 6-8), Tris (pH 8-9), KCl-borate (pH 9-10), sodium carbonate (pH 10-11), and sodium phosphate/NaOH (pH 11-12). Mannanase activity (closed symbols) was assayed at pH 4-8 using sodium citrate (circles) and sodium phosphate (dia- monds) buffers with constant temperature of 50
oC. pH Stability (open symbols) was determined by measuring the residual activ- ities after pre-incubations for 1 h in various pHs at 4
oC. Each curve represents the average of three independent experiments.
Fig. 4. Temperature optimum and thermostability of the crude
enzyme. The enzyme reaction was performed at various tempera-
tures (closed symbols) for 15 min in 50 mM sodium citrate buffer
(pH 5.5). Thermostability (open symbols) was determined by mea-
suring the residual activities after pre-incubation for 1 h at different
temperatures. Each curve represents the average of three inde-
pendent experiments.
며
, pH 11.0
에서도99%
의잔존활성을보였다(Fig. 3).
이로 보아내산성균주로분리된YB-1401
가생산하는mannanase
는최적반응pH
는Bacillus
속균주유래mannanases
들과 유사하였지만, pH 4-9
의범위내에서안정한것으로알려진B. subtilis B23 [26]
과B. subtilis G1 [20]
의mannanases
보다산성뿐아니라알칼리성의pH
범위에서도그안정성 이우수한것으로확인되었다.
그러나YB-1401
의mannanase
는pH 3.0
에서는1
시간방치하였을때잔존활성이46%
수준을유지하여
pH 3-10
의범위에서안정한것으로보고된Thielavia arenaria XZ7
유래의곰팡이효소보다는산성조건에서안정성이낮은것으로나타났다
[13]. Mannanase
의열안정성을조사한결과
45
oC
이하에서1
시간동안방치하 였을때실활되지않았으나, 50
oC
이상에서는실활되기시 작하여50
oC
에서는52%, 55
oC
에서는15%
정도의잔존활성 을보였다.
이로보아YB-1401
의mannanase
는B. subtilis BCC41051 [18], B. subtilis G1 [20]
과B. subtilis WL-3 [23]
을포함한다수의Bacillus
속균주유래mannanases
에 비해서열에대한안정성이낮았으며, B. subtilis BS5 [5]
나Sphingomonas
속균주[27]
유래의mannanases
보다는열에 안정하였다.
반응산물
Mannanase
에의한가수분해산물을조사하기위해중합도가
2-6
인MOS, LBG
와guar gum
을기질로하여효소반 응을실시한후가수분해산물을TLC
로분석한결과Fig. 5
에나타낸바와같이
mannobiose
는가수분해되지않았으며이보다중합도가큰
MOS
는가수분해되어최종산물로mannotriose
와mannobiose
가 다량 생성되었고 동시에mannose
도소량생성되었다.
그러나mannotriose
는이보다 중합도가큰MOS
에비해가수분해정도가미약한것으로 확인되었으며 이러한 결과는B. subtilis WL-3 [23]
과Bifidobacterium adolescentis [8]
의효소와유사하였다.
한 편B. subtilis WY34 [6]
의mannanase
는mannotriose
는분해하지 못하고
mannotetraose
이상의 중합도를 갖는MOS
를분해한다고알려졌다.
LBG
를mannanase
로가수분해하였을때도mannotriose
와mannobiose
가주된반응산물이며mannose
는소량생성되었는데
guar gum
의 가수분해 산물로는 매우 미량의mannotriose
와mannobiose
만 관찰되었다. Guar gum
과LBG
는galactomannan
이지만mannose
와 측쇄 결합한galactose
의양에차이가있으며, guar gum
에서galactose
의 함량이많은것으로알려져있다. Mannanase
가galactomannan
의직쇄골격인mannose
간의β-1,4
결합을가수분해할 때mannose
에측쇄결합된galactose
가반응을저해하게되어galactose
함량이높은guar gum
의가수분해산물이LBG
의 분해산물에비해적게관찰된것으로판단된다.
이러한현 상은 대부분의mannanase
가LBG
에 대한 활성이guar
gum
에대한활성보다약4-10
배정도높다는사실에서도알수있다
[12, 23].
요 약
가정에서제조된된장을산성조건에서계대배양한후균 체외
mannanase
를생산하는Bacillus sp. YB-1401
를분리 하였다.
분리균의 당이용능을 비롯한 생화학적 특성은Brevibacillus laterosporus
와유사도(61.1%)
가가장높은반 면에16S rRNA
유전자의염기서열은B. amyloliquefaciens
와유사도가가장높았다.
분리균YB-1401
의mannanase
생산성은
mannans
에의해급격하게증가하였으며,
특히곤약(4%)
이첨가된배지에서약265 U/ml
로최대생산성을보였 다.
분리균의mannanase
는55
oC
와pH 5.5
반응조건에서최 대활성을보였으며pH 3.5-11.0
의범위에서1
시간방치하였 을때실활이거의일어나지않았다.
또한LBG
와guar gum
및mannooligosaccharides
를mannanase
로분해하였을때 주된반응산물로mannobiose
와mannotriose
가생성되었으 며mannose
도소량생성되었다.
References