193
세포외 Protease를 생산하는 내염성 Bacillus sp. SJ-10 균주의 분리 동정 및 효소 특성
김은영1·김동균1·김유리1·최선영2·공인수1*
1부경대학교 생물공학과, 2국군화생방방호사령부 화학방어 연구소 생물분석과
오징어 젓갈로부터 호염성 균주를 분리하여 형태적 , 생리·생화학적 특성 및 분자계통학적 분석을 통하여 동정
하였고 , 이 균주가 세포외로 생산하는 호염성 protease 를 정제하여 그 특성을 분석하였다 . 본 균주는 NaCl
0~14%, 20~55
oC 및 pH 5~8 에서 성장하였으며 , 35
±5
oC, pH 7 및 5% NaCl 에서 최적으로 성장하였다 . 주요 지방산 조성은 anteiso-C
15:0(44.70%), anteiso-C
17:0(15.76%) 및 C
16:0(13.91%) 이었고 , menaquinone 은 MK-7 이었으며 , DNA G+C 함량은 50.58 mol% 였다 . 16S rRNA 유전자 염기서열을 통한 계통분석 결과 Bacillus이었으며 Bacillus
sp. SJ-10 으로 명명하였다 . SJ-10 균주 배양액으로부터 40% 황산암모늄 침전과 Mono Q 컬럼크로마토그래피 과 정을 거쳐 collagen 과 gelatin 을 특이적으로 분해하는 약 40 kDa 의 세포외 protease 를 정제하였다 . 이 효소는
35
±5
oC, NaCl 5
±1% 및 pH 8 에서 최적 활성을 나타냈으며 , pH 5~10 범위에서 안정하였다 . Key words □ Bacillus sp., collagen degrading enzyme, halophilic bacteria
젓갈은 어패류 , 어패류의 알 또는 내장 등을 소금에 절여 발효 시킨 한국 전통 음식으로서 (13) 염장으로 인한 짠맛과 재료 고유 의 맛 , 그리고 내부에 존재하는 미생물이 생산하는 protease 와 재 료 사이의 상호작용에 의해 생산된 성분들로 인하여 특유의 풍
미가 있다 (3). 젓갈은 음식의 맛을 내기 위한 조미료와 같은 용
도로 이용되지만 젓갈 내부에 존재하는 다양한 미생물 또는 미 생물의 대사산물 때문에 다른 음식에 첨가하거나 그 자체로도
섭취되며 (7), 이러한 젓갈 발효에 관여하는 미생물이 생산하는
protease 는 식품산업에서 광범위하게 응용되고 있다 (6). 젓갈에
존재하는 미생물은 대부분 높은 NaCl 농도에서도 성장이 가능한
내염 또는 호염성균이므로 다양한 산업 분야에 적용하기 위한 호염성 미생물의 분리 및 이들 미생물이 생산하는 호염성 효소 의 생화학적 특성에 관한 연구가 다수 보고되고 있다 (8).
염장 산업에서의 protease 를 생성하는 호염성 미생물은 염장식 품의 풍미를 증가시키고 숙성 기간을 단축시키기 위한 starter 로
서 중요하다 . 또한 protease 에 의한 분해산물이 풍미를 증진시킬
뿐만 아니라 여러 가지의 생리활성이 있음이 밝혀짐에 따라 다 양한 분야에서 protease 의 활용을 위한 연구가 진행되고 있다 (2).
염장식품을 위한 미생물은 고염농도 뿐만 아니라 다양한 온도 범위 및 산성 또는 염기성 조건에서도 생장이 가능하며 강력한
protease 활성을 지닌 것이 보다 이용 가능성이 높다 .
한편 , 호염성 미생물 또는 이들 미생물이 생산하는 protease 는
염장식품 뿐만 아니라 수산어류의 껍질을 응용한 산업에서도 널
리 사용되어 (18) 호염성 protease 를 이용하여 어류의 외피에서
collagen 의 분해산물 또는 gelatin 을 얻을 수 있으며 어폐류 폐기 물을 처리하는 산업에 있어서도 이러한 protease 또는 protease 를 생산하는 미생물을 이용하는 방법이 강구되고 있다 .
따라서 , 본 연구에서는 오징어 젓갈로부터 강력한 protease 를
생산하는 호염성 미생물을 분리하여 형태학적 , 생리·생화학적 및 분자계통학적 방법을 이용하여 동정하였으며 이 균주가 생산하 는 protease 를 분리·정제하여 그 특성을 파악하였다 .
재료 및 방법
Protease 생산균주의 분리분리원은 부산지역에서 시판되고 있는 오징어 젓갈을 사용하 였다 . 오징어 젓갈로부터 고형성분을 거른 다음 , 10% NaCl 을 첨 가한 marine agar (Difco, USA) 에 도말하여 37
oC 에서 16 시간 동 안 배양한 후 생성된 colony 를 skim milk (1%) 또는 gelatin (0.5%) 과 NaCl (5%) 이 포함된 marine agar 에서 동일한 조건으로 배양한 다음 콜로니 주변에 투명환을 형성하는 균을 선별하였다 .
선발된 균주는 다시 marine broth 에 옮겨 37
oC 에서 16 시간 동안 액체 배양하였으며 , 배양액 150~200 µ l 를 구멍 낸 gelatin (0.5%)
과 NaCl (5%) 이 함유된 agar 배지에서 동일조건으로 배양하였다 .
최종 선별된 균은 구멍 주변의 투명환을 관찰하여 가장 큰 활성 을 가지는 균으로 하였다 . 분리된 균은 동정 후 한국미생물보존 센터 (KCCM 90078) 및 일본미생물보존센터 (JCM 15709) 에 기탁 하였다 .
분리균의 배양 조건
변형된 marine 배지인 HM 배지 (NaCl 5%, yeast extract 1%,
*To whom correspondence should be addressed.
Tel: 82-51-629-5865, Fax: 82-51-629-5863
E-mail: [email protected]
분석 , G+C 함량 , 세포벽 지방산 성분분석 및 quinone 분석 등을 통하여 동정하였다 . 세포벽 지방산 성분분석은 Miller (12) 의 방
법에 따라 시행하였으며 , quinone 분석은 Kroppenstedt (10) 의 방 법을 이용하였고 , G+C 함량은 Tamaoka 등 (16) 의 방법에 따라 결정하였다 . 16S rRNA 염기서열을 결정하기 위하여는 일반적 방법에 따라 정제된 DNA 를 주형으로 하여 PCR 로 증폭하여 분 석하였다 . PCR 를 위한 primer 는 27F (5 ’ -AGAGTTTGATCMT GGCTCAG-3 ’ ) 와 1492R (5 ’ -TACGGYTACCTTGTTACGACTT- 3 ’ ) 을 사용하였고 , 94
oC 에서 45 초 , 55
oC 에서 60 초 , 72
oC 에서 60
초로 이루어진 과정을 35 회 반복하여 수행하였다 . 증폭된 DNA
단편을 Big Dye Terminator Cycle Sequencing kit (Applied Biosystems, USA) 로 정제한 다음 , DNA sequencer (Applied Biosystems model 3730XL, USA) 를 이용하여 염기서열을 결정
하였다 . 결정된 염기서열을 NCBI 의 GenBank database 및
Ribosomal Database Project (http://rdp.cme.msu.edu) 를 이용해 상
동성을 조사하였다 . 또한 이를 바탕으로 BioEdit 및 MEGA 4.1
을 이용하여 neighbor-joining 법으로 phylogenetic tree 를 작성하였 다 . Neighbour-joining 법 외에도 maximum-parsimony 법과 maxi mum-likelihood 법에 따른 phylogenetic tree 를 작성하여 유사한 결과를 얻었다 .
효소의 분리 및 정제
선발균이 생산하는 protease 분리를 위해 균을 최적배지조건
하의 37
oC 에서 24 시간 동안 배양한 후 원심분리 (4,500 rpm, 15 min, 4
oC) 하여 상등액을 회수한 다음 상등액을 10% 황산암모 늄으로 포화시켜 단백질을 침전시킨 후 얻어낸 2 차 상등액에 40% 황산암모늄으로 포화시켜 단백질을 침전시킨 다음 원심분리
(15,000 rpm, 45 min, 4
oC) 하여 얻은 침전물을 20 mM Tris-HCl (pH 8.0) 에 녹여 , 4
oC 에서 하루 동안 투석한 후 Mono-Q 컬럼크 로마토그래피를 이용한 HPLC 로 정제한 뒤 sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) 로 정제도 를 확인하였다 . 또한 silver staining 을 통해 순수분리 여부를 확 인하였다 . Gel 상에서 효소활성을 확인하기 위하여 0.5% gelatin
이 함유된 acrylamide gel 을 4
oC 에서 SDS-PAGE 한 후 세척완 충용액 (20 mM Tris-HCl; pH 8.0, 2.5% Triton X-100, 10 mM NaCl) 에 1 시간 , 배양완충용액 (20 mM Tris-HCl; pH 8.0, 10 mM CaCl
2, 0.02% NaN
3) 에 16 시간 반응시킨 후 Coomassie blue 염
색용액으로 염색한 뒤 탈색용액을 처리하여 zymogram 분석을 실시하였다 (17).
생성되는 1 µ M 의 leucine 양을 1 unit 으로 정의하였다 .
온도에 따른 효소활성 측정
Protease 활성에 미치는 온도의 영향을 검토하기 위해 정제 효
소액 100 µ l 에 0.2% gelatin 용액 300 µ l 와 반응완충용액 200 µ l
를 넣어 효소반응온도를 10
oC~60
oC 범위에서 10
oC 간격으로 하 여 1 시간 반응시켜 상대활성으로 표시하였다 . 또한 효소의 열 안 정성을 측정하기 위하여 각각의 온도에서 효소액과 반응완충액 을 1 시간 동안 10
oC~60
oC 범위에서 각각 정치시킨 후 37
oC 에서
1 시간 동안 0.2% gelatin 과 반응시켜 효소의 잔존활성도를 측정
하였다 . 상대활성의 기준은 가장 높은 활성을 가지는 온도의 활
성을 100% 로 하였다 .
pH에 따른 효소활성 측정
Protease 활성에 미치는 pH 의 영향을 검토하기 위해 정제 효
소액 100 µ l 와 0.2% gelatin 용액 300 µ l 에 각각 pH 의 범위를
4~10 으로 다르게 한 반응완충용액 200 µ l 를 넣어 37
oC 에서 1 시 간 반응시켜 활성을 측정하였다 . pH 안정성은 효소액과 pH 를 다르게 한 각각의 반응완충용액을 37
oC 에서 1 시간 동안 정치시 킨 후 0.2% gelatin 과 37
oC 에서 1 시간 동안 반응시켜 효소의 잔 존활성도를 측정하였다 . 결과 값은 효소가 가장 높은 활성을 보 이는 pH 의 활성을 100% 로 하여 상대활성으로 나타내었다 .
NaCl의 농도에 따른 효소활성 측정
Protease 활성에 미치는 NaCl 농도의 영향을 검토하기 위해 반 응완충용액에 NaCl 를 0%~16% 까지 첨가하여 효소액 및 gelatin
과 혼합하여 37
oC 에서 1 시간 반응시켜 효소의 활성을 측정하였 다 . 또한 NaCl 에 대한 안정성을 측정하고자 0%~16% 의 농도에 따라 NaCl 을 첨가한 반응완충용액과 효소를 37
oC 에서 1 시간 동 안 정치시킨 후 기질을 첨가하여 효소의 잔존활성을 측정하였다 .
상대활성은 가장 높은 활성을 가지는 NaCl 농도의 활성을 100%
로 하여 백분율로 나타내었다 .
기질에 따른 효소활성 측정
기질에 대한 protease 활성을 검토하기 위해 효소반응액에
0.2% gelatin 용액 대신 azocasein, β -casein, Z-GPLGP, Z- GPGGPA, collagen (type I, II, III, V) 및 gelatin 을 각각 1 mg/
ml 의 농도로 첨가하여 각 기질에 대한 효소의 활성을 측정하였
다 . Gelatin 을 기준으로 하여 각 기질의 상대활성을 백분율로 나
타내었다 .
결과 및 고찰
균주 분리 및 동정본 연구에서는 호염성 미생물을 분리하기 위하여 젓갈로부터
0.5% gelatin 이 함유된 평판배지 상에서 투명환의 크기를 통해
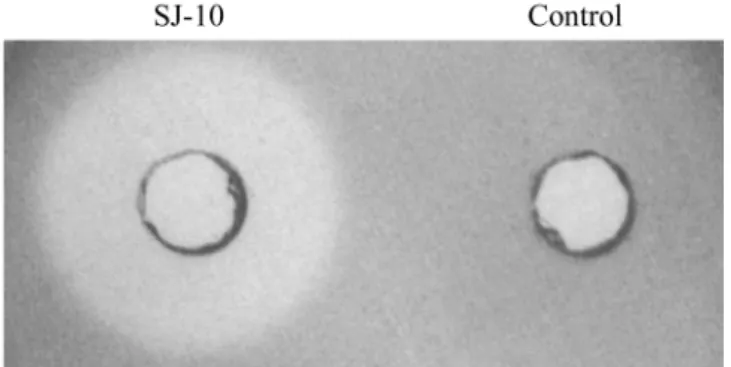
gelatin 분해능이 우수한 SJ-10 균주를 분리하였다 (Fig. 1). 분리
균은 그람양성의 간균으로 포자를 형성하고 , 운동성을 지니고 있
는 등 전형적인 Bacillus 속 균주의 형태학적 특징을 지니고 있
었다 (Fig. 2). 또한 menaquinone 은 seven isoprene units (MK-7) Fig. 1. The protease activities of strain SJ-10 on 1.2% agar plate
containing 0.5% gelatin. The isolated SJ-10 was cultured at 37
oC for 16 h. The control was marine broth. The amount of dropped sample was 150 µ l.
Fig. 2. Transmission electron microscope showing the morphology of cell, SJ-10, incubated for 16 h on HM medium at 37
oC. Bar means 200 nm.
Fig. 3. Phylogenetic location of the strain SJ-10 based on 16S rRNA gene sequences. The tree was constructed by neighbor-joining method.
Numbers at nodes are bootstrap percentage based on 1,000 resampled datasets. GenBank accession numbers are given in parentheses. Bar, 0.005
changes per nucleotide.
판단하였으며 , 16S rRNA 염기서열 (GenBank/EMBL/DDBJ accession number : FJ185224) 분석을 통해서도 Bacillus 속 균주
임을 확인할 수 있었다 . 또한 분리균은 16S rRNA 염기서열의
상동성 조사 결과 B. velezensis , B. substilis , B. amyloliquefaciens
등과 99% 이상의 유사도를 보였으며 phylogenetic tree 작성 결 과 B. velezensis 와 가장 가까웠다 (Fig. 3). B. velezensis 의 경우
menaquinone 은 MK-7 으로 분리균과 동일하였으며 DNA G+C 함
량은 약 46 mol% 로 분리균에 비해 낮은 수치를 보였다 . 주요한
세포벽 지방산 성분의 경우 anteiso-C
15:0(32.70%), iso-C
15:0(29.86%), C
16:0(13.41%) 으로 보고되었으며 이는 분리균과 같은
FA-type2 이나 분리균에 비해 iso-C
15:0의 함량이 높고 anteiso-C
17:0의 함량은 낮은 차이를 보였다 (5).
분리균의 protease 생산을 위한 최적 배양 조건을 규명하기 위
해 NaCl 농도 , 온도 , pH 등의 조건을 달리하여 배양한 결과 , NaCl 농도는 0~14%, 온도는 20~55
oC 그리고 pH 는 5~8 범위에
서 성장하였으며 35
±5
oC, pH 7, 5% NaCl 에서 가장 좋은 성장 을 보였다 . 이미 동정된 B. velezensis 의 경우 NaCl 농도 (0%~
12%) 및 온도 (15
oC~45
oC) 는 분리균보다 낮은 성장범위를 가졌으 며 , pH (5~10) 는 더 넓은 성장범위를 보여 , 분리균과는 다소 차 이가 있음을 알 수 있었다 .
효소의 정제
균체를 제거한 배양상등액을 40% 황산암모늄으로 포화시켜 침전시킨 후 원심분리하여 얻어낸 침전물을 20 mM Tris-HCl (pH 8.0) 에 녹여 , 4
oC 에서 하루 동안 투석하였으며 Mono-Q 컬 럼크로마토그래피를 이용하여 HPLC 로 정제한 결과 , 비활성 2,100 U/mg protein, 수율 0.58%, 정제배수 2.8 배로 효소를 정제 하였다 (Table 1).
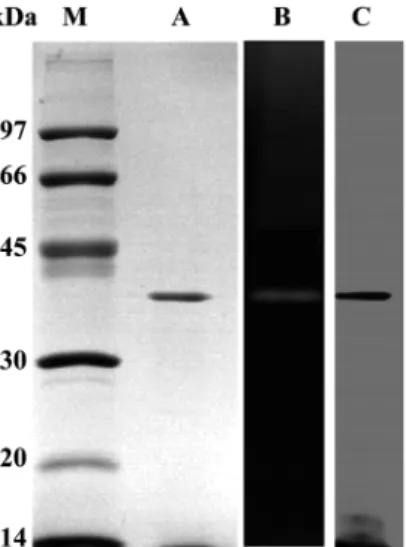
정제한 효소의 SDS-PAGE 분석 결과 분자량은 약 40 kDa 이
었다 . 기존에 보고된 collagenolytic protease 생산균주
Clostridium histolyticum (116 kDa), Vibrio alginolyticus (82 kDa), Geobacillus collagenovorans (105 kDa), B. cereus (105 kDa),
Streptococcus gordnii (98 kDa) 등이 큰 분자량을 가지는 반면 본 연구에서 정제한 효소의 경우는 Alicyclobacillus sendaiensis (37 kDa), Porphyromonas gingivalis (37.8 kDa) 에서 분리된
collagenolytic protease 와 비슷한 분자량을 보였다 (17). Zymogram
분석 결과 또한 약 40 kDa 에서 gelatin 이 분해되어 coomassiae blue 용액이 탈색된 모습을 보였다 (Fig. 4).
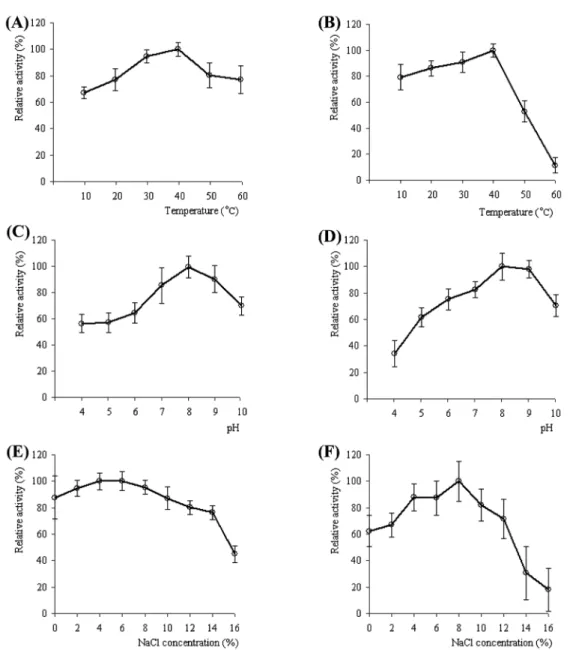
효소활성 최적 조건 및 기질 특이성
분리정제된 효소는 35
±5
oC, pH 5~10, NaCl 0%~10% 에서 높
은 활성을 나타냈으며 , 안정성의 경우 온도 40
oC, pH 8, NaCl 8% 에서 가장 높았으나 그 이상의 조건에서는 감소하였다 (Fig. 5).
일반적으로 collagen 분해효소의 최적 활성 온도는 30
oC~37
oC 로 보고되고 있는데 본 연구에서의 결과도 유사하게 나타났으며
(14), pH 4~5 의 경우 효소의 pH 안정성이 pH 8 의 활성에 대하
여 30%~60% 수준의 잔존활성을 보였으며 pH 에 대한 활성 또
한 60% 정도였다 . pH 5~10 까지 비교적 넓은 범위에서 높은 효
소 활성과 안정성을 보였으며 pH 8 에서 활성과 안정성 모두 가
장 높은 값을 나타냈는데 이러한 결과는 최적 성장 조건과 최대 효소 생산 범위가 유사하다는 보고와 일치하였다 (11).
현재까지 보고된 collagen 분해 효소의 경우 최적 활성 조건은
대부분 pH 7.5, 37
oC 이었다 (1). 그러나 본 연구에서 분리한
protease 의 경우 pH 8, 35
±5
oC 로 다른 큰 차이를 보이지 않았지
만 , 온도 (10
oC~60
oC) 와 pH (4~10) 범위에서 활성 차이가 크지
Table 1. Purification steps of protease produced by Bacillus sp. SJ-10
Steps Total protein (mg) Total activity (U
a) Specific activity (U
a/mg) Yield (%) Purification (fold)
Culture supernatant 48 36,250 755 100 1
40% Ammonium sulfate 0.16 271 1,700 0.75 2.3
Mono Q ion exchange 0.10 210 2,100 0.58 2.8
a
One unit was defined as the amount of 1 µ M leucine released
Fig. 4. Polyacrylamide electrophoresis and zymogram of the purified
protease. Lanes: M, Molecular markers; A, 12% SDS-PAGE followed
by Coomassie staining; B, 12% SDS-PAGE (containing 0.5% gelatin)
by zymogram; C, 12% SDS-PAGE followed by silver staining.
않고 안정성이 높았으며 , 14% NaCl 에서도 활성이 높은 점 등에
서 차이를 보였다 . 이러한 분리 효소의 특성은 기존에 보고된
collagen 분해 효소와 비교해 볼 때 NaCl 농도 , 온도 및 pH 등
이 다변적인 환경에의 이용에 유리할 것으로 보인다 . 또한 기질
에 따른 효소의 활성을 측정한 결과 gelatin 을 기질로 한 효소의
활성을 100% 으로 하였을 때 azocasein, β -casein 이 각각 29% 와
21% 을 나타내 효소활성이 70% 이상 감소하는 것으로 나타났으
며 gelatin 에 대한 높은 특이성을 보여주었다 . 또한 collagen type II (102%) 와 type III (99%) 효소의 활성이 gelatin 과 유사하게 높은 것으로 나타났다 (Table 2).
다양한 환경에서 분리된 Bacillus 는 여러 가지 산업용 단백질
분해 효소를 생산하며 , 빠른 성장 속도 및 다양한 기능성 물질을 생산하는 유용한 미생물로 사용된다 (4, 9, 15). Bacillus 가 생산하
Fig. 5. The activity and stability of purified enzyme. Effect of temperature on enzyme activity (A) and stability (B); Effect of pH on enzyme activity (C) and stability (D); Effect of NaCl concentration on enzyme activity (E) and stability (F).
Table 2. Substrate specificity of enzyme
Substrate Specific activity (U
a/mg) Relative activity (%)
Azocasein 30 29
β -Casein 22 21
Z-GPLGP 48 46
Z-GPGGPA 79 75
Collagen type I 62 59
Collagen type II 107 102
Collagen type III 104 99
Collagen type V 72 69
Control (gelatin) 105 100
a