Bacillus cereus를 억제하는 Bacillus subtilis HH28의 항균물질 정제와 특성규명
차현아1,2, 정다은3, 홍성욱4, 정건섭1*
1연세대학교생명과학기술학부
2농촌진흥청국립축산과학원
3연세대학교원주의과대학
4세계김치연구소
Received: November 25, 2014 / Revised: December 9, 2014 / Accepted: December 10, 2014
서 론
Bacillus cereus
는자연환경에널리분포하며호기적,
혐 기적조건에서증식하는포자형성간균이다.
따라서식품제 조,
가공,
조리과정동안에포자형태로존재하다가적절한 환경에서영양세포로증식하여식품의부패를일으키는것 으로알려져있다[1].
또한B. cereus
는enterotoxin
과구토독소를생성하여설사형
,
구토형식중독을유발한다[28].
이러한미생물때문에발생하는식품부패와식중독을예방하기 위해식품보존제를첨가한다
.
식품보존제에는합성보존제 와천연보존제가있으며합성보존제는안전성의논란으로 인해소비자들이기피하는추세이다.
따라서천연보존제에 대한관심이증가하고있고이러한천연보존제에는유기산,
식물추출물,
미생물이생산하는항균물질등이있다.
미생물이생산하는항균물질은대부분이박테리아가생산
하는단백질또는
peptide
성물질이어서인간또는동물의소화효소에의해분해되므로식품에서안전한천연보존제로 사용가능하다
.
대표적인예로1928
년유산균인Lactococcus lactis
가생산하는nisin
이라고명명된bacteriocin
으로1998
년 미국FDA
의승인으로가공치즈의천연식품보존제로사용 되고있다.
이는유산균이생성하는bacteriocin
에대한연구Purification and Characterization of an Antimicrobial Substance from Bacillus subtilis HH28 Antagonistic to Bacillus cereus
Hyun A Cha
1,2, Dawn Chung
3, Sung Wook Hong
4, and Kun Sub Chung
1*
1
Division of Biological Science and Technology, Yonsei University, Wonju 220-710, Republic of Korea
2
National Institute of Animal Science, RDA, Suwon 441-706, Republic of Korea
3
Department of Obstetrics and Gynecology, Wonju College of Medicine, Yonsei University, Wonju 220-701, Republic of Korea
4
World Institute of Kimchi, Gwangju 503-360, Republic of Korea
A bacterium producing antimicrobial substance was isolated from cheonggukjang. The bacterium was identified as a strain of Bacillus subtilis by 16S rDNA sequencing and designated as Bacillus subtilis HH28. The antimicrobial substance produced from Bacillus subtilis HH28 was purified by 0-80% ammonium sulfate precipitation, DEAE-sepharose FF column chromatogra- phy, and Sephacryl S-200 HR gel chromatography. The molecular weight of the purified antimicrobial substance was estimated to be approximately 3,500 Da using Tricine sodium dodecyl sulfate-polyacrylamide gel electrophoresis and direct detection analysis. Antimicrobial substance from B. subtilis HH28 not only inhibited B. cereus, but also Listeria monocytogenes and Vib- rio parahaemolyticus. The purified antimicrobial substance was stable at 40-80
oC, and between pH 2 and 8. Antimicrobial activ- ity of the purified substance was completely destroyed by treatment of protease, proteinase K, and pronase E, indicating that it is proteinaceous.
Keywords: Purification, antimicrobial substance, Bacillus subtilis, Bacillus cereus
*Corresponding author
Tel : +82-33-760-2252, Fax: +82-33-760-2183 E-mail: [email protected]
© 2014, The Korean Society for Microbiology and Biotechnology
가활발히이루어지는발단이되었다
[12].
그러나유산균이 생성하는bacteriocin
보다Bacillus species
가생성하는항균 물질은다양한특징을가지고넓은범위의항균력을가지고 있다[2].
Bacillus species
가생성하는항균물질은다음과같이분 류한다. Class I
은subtilin, sublancin 168, subtilosin A
등 이속해있으며,
유전자가단백질로발현된후구조가변하 여(post-translational modification)
생성된다. Class II
는13 kDa
보다작은분자량으로열과pH
에안정하고리보솜에서합성되며번역후구조변화가없는선형펩타이드로 서
coagulin, thurincin H, lichenin
등이속한다. Class III
는30 kDa
보다큰분자량으로열에약한bacteriocin
이며, megacin A group
이이에 속한다[2].
이외에도많은 다른antimicrobial peptides
가있지만단백질과DNA
서열에대 한정보부족으로분류되지못하여bacteriocin - like inhibitory substances (BLIS)
로불린다. B. subtilis LFB112
가생성하 는BLIS
는그람양성균과그람음성균모두를저해하고[30], B. subtilis MJP1
이생성하는BLIS
는그람양성균과진균에 항균활성을가진다[32].
이와같이Bacillus species
가생성 하는항균물질은종류가다양하고항균범위가넓어식품산 업뿐만아니라다양한산업에적용할수있으며적용가능 한산업의종류는다음과같다.
첫번째의약품산업으로적용할수있다
.
현재사용하고 있는항생제내성균이발견되면서사람또는동물의미생물관련질병에
bacteriocin
으로대체하려는관심이증가하고있다
[13]. Staphylococcus aureus
의강한저해능을가지는mersacidin [20]
과살정제활성을가지고있어서천연피임 약으로사용할가능성이있는subtilosin A [25]
이다.
두번째로축산산업에적용할수있다
.
병원성미생물에 감염된가축의치료와성장촉진,
폐사율감소를위해항생제 를사료급여와함께사용하고있다.
그러나이러한사용은 항생제오남용으로내성문제를일으켜몇몇나라에서는치 료용이외의사용은금지하고있는실정이다.
따라서병원성 미생물의감염을억제하고성장촉진이가능한항생제대체 제가요구되고있다. Pattnaik
등[19]
의Bacillus licheniformis
가생산하는
lichenin
은가축의제1
위 과산증을일으키는Streptococcus bovis
에대한항균력을가지고,
다양한다당류 를가수분해할수있어반추위발효개선에적용될수있는 잠재력가지고있다고보고하고있다.
세번째로식품산업에적용할수있다
.
현대의소비자들 은합성보존제가 첨가되지않고최소의가공과정만거치는‘fresh foods’
을원한다.
따라서천연합성보존제로사용할수 있는bacteriocin
에대한관심이급증하고있다.
예를들어antimicrobial peptides P45 [26, 27], paenibacillin [6]
그리 고김치에서분리한Bacillus brevis
가생성하는항세균및항진균활성
peptide
등이있다[7].
네번째로환경산업에대한적용이다
. Bacillus species
가 생성하는항균,
항진균활성이있는bacteriocin
은‘biocontrol agents’
로사용할수있다.
예를들어대두의뿌리에서분리 한Bacillus thuringiensis NEB17
이생성하는bacteriocin
은근류형성을도와식물의성장을증진시키고,
식물의질 병저항성을높여준다[9].
따라서본연구에서는청국장에서분리한
Bacillus subtilis HH28
가생성하는항균물질의정제및특성을규명함으로써 여러가지산업에적용가능성을검토하고자하였다.
재료 및 방법
미생물 분리 및 배양
본실험의미생물은전통적으로제조한청국장및이를제 조할때사용한볏짚을이용하여분리하였다
.
미생물을분리 하기 위하여 청국장과볏짚 시료 각각을 멸균생리식염수[0.85% (w/v) NaCl]
와1:9
비율로혼합한후, homogenizer (Stomacher 400, Seward, England)
로10
분간균질화하여 현탁액을제조하였다.
이현탁액을멸균생리식염수로십진 희석한후tryptic soy agar (TSA; Difco, Detroit, MI, USA)
에100
μl
씩적가하여도말하고37
oC
에서14
시간동안배양한 후미생물집락을순수분리하였다.
항균물질 활성 측정
실험에사용된피검균
B. cereus KCCM 12667
는tryptic soy broth (TSB, Difco, MI, USA)
에1% (v/v)
접종하여37
oC
에서24
시간동안배양하였다.
분리미생물의B. cereus
에대한항균활성은paper disc
방법[29]
을사용하여확인하 였다. B. cereus
을0.4% (v/v)
접종한tryptic soy soft agar
를중층한평판배지에직경8 mm
의paper disc (Advantec, Toyo Roshi Kaisha Ltd., Japan)
를 올리고 항균물질을50
μl
을적가하여37
oC
에서12
시간동안배양한후생성된 투명환의형성유무를관찰하였다.
분리균주의 동정
항균활성을나타내는분리균주를동정하기위해서형태 학적
,
생화학학적동정과유전학적동정을실시하였다.
형 태학적인동정은그람염색과포자형성관찰을하였다.
생화 학적동정은API 50 CHB kit (bioMerieux, Marcy IEtoile,
France)
을사용하여49
개의탄소원에대한이용성을조사하고
, API 50 CHB database V3.0(http://apiweeb.biomerieux.
com)
을 통해 동정결과를 얻었다.
유전학적 동정은16S
ribosomal DNA
염기서열분석하는것으로분리미생물의chromosomal DNA
를template
로universal primer
인27
forward primer (5'-AGAGTTTGATCATGGCTCAG-3')
와1492 reverse primer (5'-GGATACCTTGTTACGACTT-3')
를사용하여16S rDNA gene
일부를증폭하였다.
증폭된약1,400 bp
의fragment
를T-vector (Invitrogen, Carlsbad, CA, USA)
에결합시켜형질전환한후T-vector sequencing primer
를통해염기서열을분석하고, BLAST search program
을 이용하여GenBank (NCBI)
의ribosomal DNA gene sequence
와비교동정하였다.
생육시기에 따른 항균 활성조사
분리 미생물을
glucose minimal medium (0.002%
ammonium molybdate, 0.003% CaCl
2·2H
2O, 0.0002%
FeSO
4·7H
2O, 1% Glucose, 0.7% K
2HPO
4, 0.15% KH
2PO
4, 0.05% MgSO
4·7H
2O, 0.004% MnSO
4·4H
2O, 0.5% yeast extract) 500 ml
에1% (v/v)
접종하여40
oC
에서180 rpm
으 로교반하여배양하였다. 0-144
시간동안진탕배양하는동 안spectrophotometer (VersaMax ELISA Microplate Reader, Molecular Devices Sunnyvale, CA, USA)
로660 nm
에서 흡광도를측정하여생육곡선을그리고,
생육시기에따른항 균활성을측정하였다.
활성역가는분리미생물배양액ml
당AU (activity unit)
로측정하였다. AU
는항균물질을2
배 씩점차적으로희석하여투명환을형성하는최대희석배수 의역수값을취하고,
이값을1 ml
로환산하는값에곱하여AU/ml
로정하였다.
항균물질의 분리 및 정제
선발 미생물을
glucose minimal medium 500 ml
에1%
(v/v)
접종하여40
oC
에서3
일동안배양하였다.
선발미생물 의배양상등액에황산암모늄을80%
의포화도까지첨가하 고, 4
oC
에서12
시간 동안방치한후원심분리(20,000
×g, 4
oC, 30
분)
하여 얻은침전물을10 mM Tris·HCl
완충용액(pH 8.0)
에용해시켰다.
그리고투석막(MWCO 6-8 kDa cut- off, Spectrum Laboratories, Inc.)
으로4
oC
에서12
시간동안 투석을 한 후DEAE-sepharose fast flow (Pharmacia, Biotech. AB Uppsala, Sweden) column (25
×400 mm)
과Sephacryl S-200HR (Pharmacia) column (15
×960 mm)
을이용하여정제하였다.
각분획의단백질함량은280 nm
로측정하였고,
각각항균활성을측정하여활성이존재하는 분획만취합하여동결건조하였다.
모든정제과정은단백질 성항균물질의변성을최소화하기위해서4
oC
에서수행하 였다.
단백질 함량 측정
항균물질단백질정량은
Lowry
방법을변형하여실시하였다
[17].
항균물질시료50
μl
에biuret reagent (0.75 mmol/l cupric sulfate, 94 mmol/l sodium hydroxide) 550
μl
를혼 합하고25
oC
에서10
분간반응시킨후, Folin and Ciocalteu’s phenol reagent (Sigma Co., St. Louis, MO, USA) 25
μl
를 넣고25
oC
에서30
분 동안 반응시켰다.
그리고spectro- photometer
로725 nm
에서흡광도를측정하여나타내었고,
표준곡선은bovine serum albumin (BSA; Sigma Co.)
를사 용하였다.
항균물질 분자량 측정
항균물질의분자량을측정하기위해
Tricine-sodium dodecyl sulfate polyacrylamide gel electrophoresis (Tricine-SDS- PAGE)
를수행하였다[21]. 20% polyacrylamide gel
를사용 하여100 V
로5
시간동안전기영동한후silver staining kit (Amersham Biosciences AB Inc., Sweden)
로염색했다.
분 자량marker
는ultra-low range molecular weight marker (1,060-26,600 Da, Sigma)
을사용했다. Tricine SDS-PAGE
를 수행한후 밴드가 항균물질인지 확인하기위해direct detection
을하였다[3]. Tricine SDS-PAGE
한polyacrylamide gel
을25% (v/v) isopropanol
과10% (v/v) glacial acetic acid
의혼합물에2
시간동안고정을하고멸균증류수로6
시 간동안wash
한후B. cereus KCCM 12667
가중층된배지 에올리고40
oC
에서6
시간배양하여투명환의유무를확인 하였다.
항균 spectrum 조사
Bacillus subtilis, B. licheniformis, B. cereus
의배양배지 는TSB, Staphylococcus aureus, Salmonella Typhimurium, Pseudomonas aeruginosa, Shigella flexineri, S. sonnei, S. boydii, Escherichia coli O157:H7
의배양배지는nutrient broth (NB, Difco, MI, USA), Vibrio parahaemolyticus
는2.5 % (w/v)
염화나트륨이포함된TSB
배지를사용하였다. Streptococcus mutans, Listeria monocytogenes
는Brain Heart Infusion broth (BHI, Difco), Candida albicans
는Yeast Mold broth (YM, Difco, MI, USA), Aspergillus flavus, A. fumigatus, Rhizopus oryzae, Mucor circinelloides, Penicillum camemberti
는potato dextrose agar (PDA,
Difco)
배지를사용하여배양하였다.
피검균으로사용한균주들은생명자원센터
(KCTC),
한국농업미생물자원센터(KACC)
그리고한국미생물보존센터(KCCM)
에서분양받았다(Table
2).
배양온도는C. albicans
는30
oC, A. flavus, A. fumigatus,
R. oryzae, M. circinelloides, P. camemberti
는25
oC,
나머 지균들은37
oC
에서배양하였고이들균주에대한길항력 은분리균주배양상등액의황산암모늄침전액을사용하여측정하였고
paper disc
주변의투명환의직경으로비교하였다.
항균 작용 기전항균작용기전은
Nicolas
등[16]
의방법으로수행하였다. B. cereus KCCM 12667
를TSB
에5% (v/v)
접종한후2
시간진탕배양한배양액에항균물질
(32 AU/ml)
를첨가하였고,
항균물질을첨가하지않은
B. cereus
배양액을대조군으로 사용하였다.
항균물질첨가후12
시간동안 진탕배양하며2
시간간격으로생균수를측정하였다.
항균물질의 안정성
항균물질의
pH
안정성을 측정하기 위해pH 2.0-4.0
은0.1 M glycine-HCl
완충용액, pH 4.0-6.0
은0.1 M sodium acetate
완충용액, pH 6.0-8.0
은0.1 M sodium phosphate
완충용액, pH 8.0-10.0
은Tris-HCl
완충용액을사용하였다.
각각의pH
완충용액과정제항균물질을1:15
비율로혼합 하여배양온도인40
oC
에서12
시간방치한후잔존항균활 성을측정하였다.
대조군으로는항균물질을첨가하지않은 각각의pH
완충용액을사용하였다.
항균물질의온도안정성 을측정하기위하여정제한항균물질을40-80
oC
에서12
시 간, 100
oC
는1
시간방치한후잔존항균활성을측정하였다.
각종효소의영향을측정하기위해lysozyme (EC 3.2.1.17, Sigma),
α-amylase (EC 3.2.1.1, Sigma), lipase (EC 3.1.1.3, Type VII, Sigma), trypsin (EC 3.4.21.4, Sigma), protease (EC 3.4.24.31, Sigma), Proteinase K (EC 3.4.21.64, Sigma), pronase E (EC 3.4.24.4, Merck)
를10 mM sodium phosphate
완충용액(pH 7.0)
에4 mg/ml
이되도록준비하였다.
정제한 항균물질에각각의효소를2 mg/ml
첨가하여37
oC
에서30
분 동안방치한후항균물질의잔존활성을측정하였다.
결과 및 고찰
항균활성 균주의 분리 및 동정
전통적인방법으로제조한청국장과제조할때사용한볏 짚을멸균생리식염수로희석한후
, TSA
배지에서14
시간 동안배양한후,
미생물의집락을관찰하였고다양한미생물 의집락이관찰되었다. B. cereus
에대한항균활성실험결과,
항균활성이가장높은분리균주1
종을선발하였다.
분리한미생물은포자를형성하는그람양성간균이며
, API
50 CHB system
을이용한당이용성조사를통한동정결과는
Bacillus subtilis
로판정되었다.
보다정확한동정을위 하여16S rDNA
의염기서열을결정하고, GenBenk
에등록 된다른균주의염기서열과비교한결과B. subtilis
와98%
의상동성을보였다
.
따라서,
선발균주는B. subtilis
로동정 되었고, B. subtilis HH28
로명명하였다.
분리균주의 생육시기에 따른 항균활성
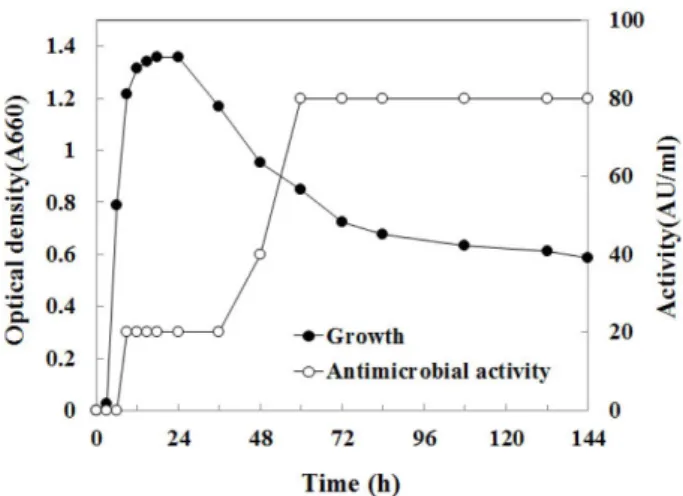
B. subtilis HH28
의생육에따른항균활성특성을조사하 였다(Fig. 1).
생육곡선측정결과,
배양12
시간까지대수증 식기이고24
시간까지정체기이며그이후사멸기에도달하 였음을알수있다. B. cereus
에대한항균활성은생육이대 수기인9
시간부터생성되어사멸기인60
시간에가장높은 활성(80 AU/ml)
을보였다.
많은항균물질이영양원이고갈되는정체기에축적되는것 과같이본연구의항균물질또한정체기인
12
시간이후부 터항균활성이나타나는것을알수있다[22].
또한항균활성 이가장높은시기인60
시간부터실험이종료된시점144
시 간(6
일)
까지활성이감소하지않고유지되는것을확인할수 있다.
반면Guo
등[5]
의B. licheniformis B11
이생산하는bacteriocin - like substance (BLIS)
는배양후20
시간이지 난후에항균활성이감소하고,
또한Yang
등[32]
의항균물 질도배양시간이길어짐에따라항균활성이감소하는것을 확인할수있다.
항균물질의 분리 및 정제
B. subtilis HH28
의배양상등액500 ml
을0-80%
황산암 모늄침전한후,
크로마토그래피방법으로정제하였다.
황산 암모늄침전시료를DEAE-sepharose fast flow
를사용하여 음이온교환크로마토그래피를실시하였고, 0-1 M
의이온강 도를사용하여항균물질을용출하였다(Fig. 2).
이온교환크 로마토그래피에서활성이있는분획143-156
번을취합한후sephacryl S-200HR
을사용하여겔크로마토그래피를하였다
(Fig. 3).
겔크로마토그래피의항균활성이있는분획46- 56
번을모아정제된항균물질로사용하였다.
항균물질정제 의결과를Table 1
에나타냈고, 19.7
배의정제도와38.4%
의 수율로정제되었다.
Fig. 1. Growth and antimicrobial activity of Bacillus subtilis
HH28 in the glucose minimal medium.
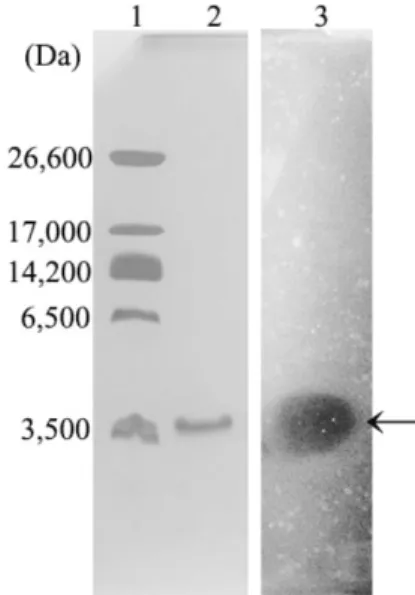
항균물질의 분자량 측정
정제한항균물질의분자량을측정하기위해
Tricine SDS-
PAGE
를실시하였고,
항균물질유무를확인하기위해direct
detection
방법을수행하였다.
정제된항균물질의전기영동과
direct detection
을한결과로밴드가하나이고그위치에 투명환이형성됨을확인하였다(Fig. 4).
또한marker
와시료 의이동성을비교하여항균물질의분자량이약3,500 Da
임 을예측하였다.
B. subtilis
가생산하면서항균물질중약3,500 Da
분자 량을가지는것은lantibiotic
에속하는subtilin, subtilosin A, sublancin 168, ericin S
등이있다. Subtilin
은pentacyclic peptide
형태로3,319 Da
이고[24], ericin S
는3,442 Da
이며subtilin
과4
개의아미노산잔기만다르고매우유사한특성 을 가진 물질[23]
이다. subtilosin
은3,399 Da
으로post- translational modification
을통해macrocyclic
형태가되며[15], sublancin 168
은3,877 Da
이고두개의이황화결합으Fig. 2. Ion exchange chromatography of the antimicrobial
substance from Bacillus subtilis HH28 on the DEAE-sephar- ose FF. The DEAE-sepharose FF ion exchange column (25 × 400 mm) was equilibrated with 10 mM Tris·HCl buffer (pH 8). The column was eluted with a linear gradient of 0-1 M NaCl in 10 mM Tris·HCl buffer (pH 8) at a flow rate of 1 ml/min. The volume of each fraction was 5 ml.
Fig. 3. Gel chromatography of the antimicrobial substance from Bacillus subtilis HH28 on the sephacryl S-200HR. Seph- acryl S-200HR column (15 × 960 mm) equilibrated with 10mM Tris·HCl buffer (pH 8). The column was eluted with 10 mM Tris·HCl buffer (pH 8) at a flow rate of 0.5 ml/min. The volume of each frac- tion was 2 ml.
Table 1. Purification summary of antimicrobial substance produced by Bacillus subtilis HH28.
Steps
*Volume
(ml)
Total protein (mg)
Total activity (AU/ml)
Specific activity (AU/mg)
Purification (fold)
Yield (%)
Culture broth 500.0 1,159.6 40,000 34.5 1 100.0
AS 3.0 101.1 30,720 303.8 8.8 76.8
IEX 2.0 49.2 20,480 416.3 12.1 51.2
GF 1.5 22.6 15,360 678.5 19.7 38.4
*AS, ammonium sulfate precipitation; IEX, ion exchange chromatography; GF, gel chromatography
Fig. 4. Tricine SDS-PAGE and direct detection of purified anti- microbial substance from Bacillus subtilis HH28. Lane 1, Ultra-low range molecular weight marker (26,600 Da, Triosephos- phate isomerase from rabbit muscle; 17,000 Da, Myoglobin from horse heart; 14,200 Da, α-Lactalbumin from bovine milk; 6,500 Da, aprotinin from bovine lung; 3,496 Da, insulin chain B.); Lane 2, Purified antimicrobial substance eluted by sephacryl S-200HR;
Lane 3, Gel overlayed with Bacillus cereus KCCM 12667.
로연결된
lanthionine
이포함되어있는형태[18]
로알려져 있다.
본연구의항균물질의정확한구조를파악하기위해아미 노산서열을분석방법인
Edman
법[4]
을시행하였다.
그러 나N-terminal
이block
되어아미노산서열을분석하지못하 였고,
따라서추후의본연구의항균물질의자세한구조분 석이필요한것으로판단되었다.
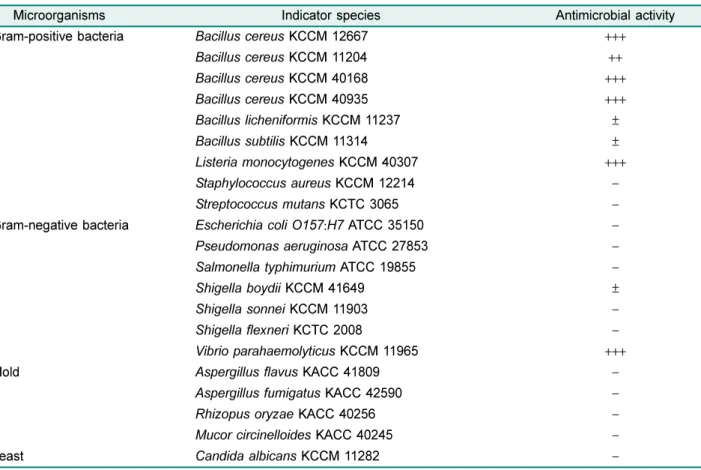
항균 spectrum
B. subtilis HH28
이생성하는항균물질의병원성세균및 곰팡이,
효모에대한항균효과는Table 2
에요약되어있다. Paper disk method
를통하여항균활성을측정한결과,
그 람 양성균인B. cereus KCCM 12667, KCCM 11204, KCCM 40168, KCCM 40935, B. licheniformis KCCM 11237, B. subtilis KCCM 11314, Listeria monocytogenes KCCM 40307
에대한 저해활성을나타내었으며,
특히B.
cereus
와L. monocytogenes
의활성이큰것을확인할수있 다.
그람음성균에서는Shigella boydii KCCM 41649
에약 한 저해 활성을 나타내었고, Vibrio parahaemolyticus
KCCM 11965
에서큰저해활성을나타내었다.
그리고곰팡이와효모에서는활성이없었다
.
대부분의
Bacillus spp.
가생산하는bacteriocin
은그람음 성균을억제하지못하고그람양성균만을억제하였다[8].
예 를들어 청국장에서 분리한B. subtilis W42
의 경우도B.
cereus
와L. monocytogenes
에큰항균활성을나타내지만그 람음성균에는활성을없었다[11].
그러나본연구의항균물 질은그람음성균의V. parahaemolyticus
에높은활성을보 였는데이균은해산물에의한식중독을일으키는균으로잘 알려져 있으며,
중국 전통 콩 발효 식품에서 분리한B.
subtilis NT-6
가이에대한항균활성이있음을보고한바있 다[31].
또한그람양성균중장류의식중독균인B. cereus
와 샐러드와같은최소가공처리를한채소에서식중독을일으 키는L. monocytogenes
에대한항균활성이매우우수하여천 연식품보존제로사용할경우큰효과를나타낼것으로사 료되었다.
그러나장류발효시유용하게작용하는B. subtilis
와B. licheniformis
에대한항균활성을가지고있지만그활 성이매우작아큰영향을미치지못할것으로사료되었다.
항균 작용 기전
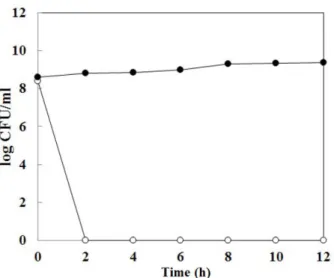
2
시간동안배양한B. cereus
에정제한항균물질을첨가Table 2. Inhibitory spectrum of antimicrobial substance from Bacillus subtilis HH28.
Microorganisms Indicator species Antimicrobial activity
Gram-positive bacteria Bacillus cereus KCCM 12667 +++
Bacillus cereus KCCM 11204 ++
Bacillus cereus KCCM 40168 +++
Bacillus cereus KCCM 40935 +++
Bacillus licheniformis KCCM 11237 ±
Bacillus subtilis KCCM 11314 ±
Listeria monocytogenes KCCM 40307 +++
Staphylococcus aureus KCCM 12214 −
Streptococcus mutans KCTC 3065 −
Gram-negative bacteria Escherichia coli O157 : H7 ATCC 35150 −
Pseudomonas aeruginosa ATCC 27853 −
Salmonella typhimurium ATCC 19855 −
Shigella boydii KCCM 41649 ±
Shigella sonnei KCCM 11903 −
Shigella flexneri KCTC 2008 −
Vibrio parahaemolyticus KCCM 11965 +++
Mold Aspergillus flavus KACC 41809 −
Aspergillus fumigatus KACC 42590 −
Rhizopus oryzae KACC 40256 −
Mucor circinelloides KACC 40245 −
Yeast Candida albicans KCCM 11282 −
−, no inhibition zone; ±, below 10 mm; +, below 13 mm; ++, 13-15 mm; +++, above 15 mm.
한후
2
시간만에B. cereus
가사멸한것을확인할수있었 다(Fig. 5). 2
시간 동안 배양한B. cereus
의생균수8.4 log (CFU/ml)
에서B. subtilis HH28
이생성하는항균물질을첨 가한2
시간후에는B. cereus
의콜로니를확인하지못했다.
이 러한결과를보아본연구에서정제한항균물질은B. cereus
에대한살균작용(bactericidal action)
이있음을확인하였다. Kindoli
등[10]
의청국장에서분리한B. subtilis H27
의항균 물질 또한살균작용을하며, 1,600 AU/ml
을처리했을 때12
시간동안서서히L. monocytogenes
가사멸되는것을확 인할수있었다.
그러나본연구의항균물질은32 AU/ml
을 처리하여2
시간만에B. cereus
가모두사멸되고12
시간동 안유지되는것으로보아항균물질의살균작용이높다는것 을확인할수있었다.
항균물질의 안정성
pH
안정성을측정하기위해pH 2-10
까지각각완충용액으로정제된항균물질을희석하여배양온도인
40
oC
에서12
시 간방치후잔존항균활성을측정하였다.
본연구의항균물 질은pH 2-8
까지100%
의항균활성을유지하였고, pH 9
에 서는50%
로pH 10
에서는25%
로항균활성이 감소하였다(Table 3). pH
의영향으로B. cereus
의성장을저해한것인 지확인하기위하여대조군으로완충용액의항균활성실험 을실시하였는데완충용액으로인해B. cereus
가저해되지 않았다.
이결과로보아본연구의항균물질은염기성보다산성일때안정성이더높다는것을확인할수있었다
. Lee
등
[14]
의B. subtilis SC-8
이생성하는항균물질은pH 3
에서항균활성이감소하고
pH 4-10
은항균활성이유지됨을알수 있다.
본연구의항균물질과반대로산성에서안정성이낮고 염기성에서안정성이높다.
온도안정성측정은정제한항균물질을
40-80
oC
는12
시간 동안방치하였고, 100
oC
는1
시간방치후의잔존항균활성 을측정하였다. 40-60
oC
에서12
시간동안방치한후, 100%
의잔존항균활성을유지하였고
, 80
oC
에서12
시간동안방 치한경우, 12.5%
로잔존항균활성이감소하였다(Table 3).
Lee
등[14]
에서80
oC
와100
oC
에서1
시간방치하였을때B.
cereus
에대한항균활성이소실되었다. Kindoli
등[11]
의경 우60-80
oC
에서15
분동안방치하였을때항균활성이50%
가 감소하였고
, 100
oC 10
분 방치하였을 때20%
로L.
monocytogenes
의항균활성이감소하였다.
이러한결과로보 아본연구의항균물질은온도에대한안정성이높음을알 수있다.
단백질분해효소인
trypsin
의처리로50%
로항균활성이 감소하였고, protease, proteinase K, pronase E
의 처리시 항균활성이소실되었다. Lysozyme,
α-amylase, lipase
를처리하였을경우
,
모두100%
활성이유지됨을알수있었다(Table 3).
단백질분해효소에의해항균활성이소실되므로본연구의항균물질을단백질성물질임을확인하였다
. Fig. 5. Inhibition mode of antimicrobial substance from Bacil-
lus subtilis HH28. Opened circle, Bacillus cereus KCCM12667 culture with purified antimicrobial substance; closed circle, Bacillus cereus KCCM12667 culture without purified antimicrobial sub- stance.
Table 3. Effect of enzyme treatment, heat and pH on the anti- microbial activity of B. subtilis HH28.
Treatment Residual activity (%) pH
2 100
4 100
6 100
8 100
10 25
Heat
40
oC 100
60
oC 100
80
oC 12.5
100
oC 25
Enzyme
lysozyme 100
α
-amylase 100
lipase 100
trypsin 50
protease 0
proteinase K 0
pronase E 0
요 약
청국장으로부터
Bacillus cereus
에대한항균활성이가장높은균주를분리하여형태학적
,
생화학적특성과16S rDNA
염기서열결정을통해
Bacillus subtilis HH28
으로동정및 명명하였다. B. subilis HH28
의생육시기에따른항균활성 을측정해본결과,
생육이대수증식기인9
시간부터생성되 어사멸기인60
시간에가장높은활성(80 AU/ml)
을나타내 었고144
시간(6
일)
까지항균활성을유지하였다.
항균물질의 정제는 황산암모늄 침전과DEAE-sepharose fast flow, sephacryl S-200HR
를 이용하였고, 19.7
배의 정제도와38.4%
의수율로정제되었다. Tricine SDS-PAGE
와directed detection
을통해항균물질의분자량이약3,500 Da
임을알 수있었다.
또한본연구의항균물질을B. cereus
뿐만아니 라Listeria monocytogenes, Vibrio parahaemolyticus
의식 중독균에서도우수한항균활성을나타내었고,
항균작용의 기작은미생물을사멸시키는살균작용이였다.
또한온도안 정성실험에서40-80
oC
까지안정했고, pH
안정성실험에서 는pH 2-9
까지안정하여비교적온도와pH
에안정하였다.
효소에대한영향실험에서는단백질분해효소에의해항균 활성이실활되어본연구의항균물질은단백질성임을알수 있었다.
위와같은특성으로보아B. subtilis HH28
이생산 하는항균물질은천연식품보존제및사료보존제,
항생제 대체의약품으로사용할수있는잠재력을가지고있으며,
향후이항균물질의정확한구조및특성규명등의연구가 필요하다.
Acknowledgments
This study was supported by the 2012 Korea Institute of Plan- ning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (iPET), Ministry of Agriculture, Food and Rural Affairs (MAFRA).
References