Purification and Characterization of Metalloprotease from Serratia marcescens PPB-26 and Its Application for Detergent Additive
Shikha Thakur, Nirmal Kant Sharma, Neerja Thakur, and Tek Chand Bhalla*
Department of Biotechnology, Himachal Pradesh University, Summer Hill, Shimla-171005, India
Received: September 5, 2018 / Revised: November 20, 2018 / Accepted: November 21, 2018
Introduction
Proteases (E.C 3.4) are enzymes that catalyse the hydrolysis of peptide bonds in proteins [1]. They are one of the largest and most diverse families of enzymes.
Although protease are ubiquitous in nature [2], however microbes are their most preferred source due to their large biochemical diversity, limited space requirement for growth and ease in genetic manipulations [3]. Micro- bial proteases, specially bacterial, constitute 60% of the total industrial enzyme sale in the world [4] owing to
their versatile applications in industries such as deter- gent, leather, food, textile and pharmaceutical [5−7].
Proteases are also useful in basic research as their selec- tive cleavage of peptide bonds helps elucidate the struc- ture-function relationships [8]. Besides this, they have extensive applications in development of eco-friendly technologies, bioremediation processes and in synthesis of peptides [9, 10].
Largest application of proteases is however in the detergent industry, where they are added in very small amounts to detergent preparations for increasing their cleaning ability [11]. Proteases have been added to laun- dry detergents for over 50 years and account for 30% of the worldwide production of enzymes [12, 13]. A deter- gent that does not contain an enzyme may not remove In this study, the extracellular metalloprotease from Serratia marcescens PPB-26 was purified to homoge- neity via ethanol fractionation and DEAE-cellulose column chromatography. Thus, a 3.8-fold purification was achieved with a 20% yield and specific activity of 76.2 U/mg. The purified protease was a 50-kDa mono- mer whose optimum pH and temperature for activity were 7.5 and 30℃ respectively; however, it was found to remain active in the 5−9 pH range and up to 40℃ for 6 h. The protease had a half-life of 15 days at 4℃, an optimum reaction time of 10 min, and an optimum substrate (casein) concentration of 0.25%. Furthermore, the Michaelis constant (Km) and reaction velocity (Vmax) of the protease were calculated to be 0.28% and 111.11 µmoles/(min·mg)-1,respectively. The protease was stable when subjected to metal ions (2 mM), show- ing increased activity with most (especially CoCl2 and MgSO4 (30.54% increase)). It was also stable when exposed to oxidizing agents, bleaching agents, and detergents (5% v/v for 60 min). It retained 93% of its activity in non-ionic detergents (Tween-20, Tween-80, and Triton X-100). Moreover, wash performance anal- ysis in commercial detergents (Ariel and Tide) showed that not only was the protease capable of protein stain removal, but also reduced cleaning time by 80% when added to detergents. Thus, the Serratia marc- escens PPB-26 metalloprotease appears to be a promising new candidate as a laundry additive in the deter- gent industry.
Keywords: Metalloprotease, ion-exchange chromatography, MALDI-TOF, N-terminal sequencing, detergent additive
*Corresponding author
Tel: +91-177-2831948, Fax: +91-177-2831948 E-mail: [email protected]
© 2019, The Korean Society for Microbiology and Biotechnology
the stains completely resulting in permanent residues [14]. The proteases form a key constituent in the com- mercial detergents since they enhance their efficiency, are biodegradable and increase the performance to cost ratios [15−17]. For an enzyme to be used as a detergent additive it should have a broad substrate specificity, sta- bility in the presence of surfactants, oxidizing agents, bleaching agents and compatibility with detergents [18].
However, very few bleach stable enzymes have been reported so far [19, 20]. The performance of an enzyme in detergents is also dependent on the detergent compo- sition, type of stains, water hardness, washing tempera- ture and procedure [14].
There is, thus, a huge industrial demand for proteases with novel and better properties. And although different proteases have been isolated from several bacteria and fungi, but only a few have such properties that make them a commercially viable choice for exploitation in the detergent industry. Keeping this view in the present research work the properties of metalloprotease of Serratia marcesence PPB-26 were evaluated. The prote- ase was purified to homogeneity and its biochemical characterization and stability studies were done aimed to determine possible eco-friendly application of the enzyme as a detergent additive.
Materials and Methods
Chemicals and media components
The media ingredients were obtained from Hi-Media, India. Pre-packed ion-exchange column (HiPrepTM DEAE Cellulose) and reagents of polyacrylamide gel electrophoresis (PAGE) were purchased from Sigma Aldrich, USA. Sodium dodecyl sulphate (SDS) and native PAGE protein molecular size markers were obtained from GeNei (Merck bioscience), India. All other chemicals and reagents were of analytical grade and procured from standard companies.
Microorganism and its culture conditions
The bacterial strain, Serratia marcescens PPB-26 (accession number KJ735909) used in present work is a hyper producer of protease, screened, isolated and char- acterized in our previously published research work [21].
It was grown in optimized media (2.5% dextrose, 1%
casein, 1.5% tryptone, 1% yeast extract, 0.5% potassium phosphate, 0.25% calcium chloride and 0.5% magnesium sulphate), pH 7 at 30℃ for maximum protease produc- tion.
Protease assay
Protease activity was measured according to the meth- odology of Manachini et al. [22] using casein as sub- strate. To 4 ml substrate solution i.e., 0.8% (w/v) casein in 0.15 M Tris-HCl (pH 7.5), 1 ml enzyme was added and incubated at 30 ºC for 10 min. The reaction was stopped by adding 5 ml stopping reagent i.e., 5% (w/v) of Trichlo- roacetic acid (TCA). The reaction mixture was then vor- texed and centrifuged at 10,000 × g for 5 min and optical density (OD) of the supernatant measured at 275 nm.
One unit of protease activity was defined as the amount of enzyme required to release 1 µg of tyrosine per minute under standard assay conditions.
Purification of S. marcescens PPB-26 protease
All purification steps were performed at room tem- perature. S. marcescens PPB-26 was grown overnight in 500 ml media and then centrifuged. Crude enzyme in the supernatant was precipitated using three different fractionation methods: ammonium sulphate, ethanol and acetone. These were added to 10 ml supernatant at different saturation levels (10% to 90%) and kept over- night at 4℃ with slow stirring. Precipitates were col- lected by centrifugation at 10,000 ×g for 30 min and then solubilised in minimum volume of 0.15 M Tris-HCl buffer (pH 9) and assayed for protease activity. Satura- tion with which highest protease activity is measured was thus selected for large scale (500 ml) fractionation.
Precipitates obtained from this were then subjected to ion exchange chromatography using DEAE Cellulose column. The column (20 ml bed volume) was washed with 1 M KCl (10 ml) and then pre-equilibrated with 0.15 M Tris-HCl buffer (pH 7.5, 20 ml) containing 0.1 M NaCl. Protein precipitate sample was loaded on the col- umn and allowed to bind. Elution was done with 100 ml linear gradient of 0.0 M to 1.0 M NaCl in Tris-HCl buffer (0.15 M, pH 7.5). Fractions of 3 ml each were collected at a flow rate of 0.5 ml/min and assayed for protease activ- ity. The active fractions were pooled together and used as purified enzyme preparation.
Polyacrylamide gel electrophoresis
The pooled fractions were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) on 12% polyacrylamide gel using a medium range marker (10−250 kDa). Native-PAGE electrophoresis was also done on 8% polyacrylamide gel using high molecu- lar weight markers (29−205 kDa). Coomassie brilliant blue R-250 was used for gel staining. Destaining was done with 10% methanol and 10% acetic acid. For SDS- PAGE gel silver staining was also done.
MALDI-TOF and N-terminal sequence analysis
Matrix assisted laser desorption ionization-time of flight (MALDI-TOF) analysis of the purified protease was performed at Central Instrumentation Facility, School of Life Sciences, Jawaharlal Nehru University, New Delhi. The protein band was excised from silver stained gel and destained with 100 mM sodium thiosul- phate and 30 mM potassium ferricyanide. The destained band was washed with ammonium bicarbonate: acetoni- trile (1:1) and incubated in 100% acetonitrile for 1 h at room temperature for dehydration. The band was digested with trypsin and incubated at 37℃ for 16 h.
Digested peptides were dried and resuspended in 0.1%
trifluoroacetic acid. This suspension (0.5 ml) was mixed with same amount of cyano-4-hydroxycinnamic acid (10 mg/ml in 50% acetonitrile, 0.1% trifluoroacetic acid) and spotted onto a MALDI-target plate for analysis. The peptides were analyzed by BurkerAutoflex MALDI-TOF (BurkerDaltonics) and spectra interpreted using Burk- arBiotools Version 2.2 (BurkerDaltonik). Homologous proteins were searched in the NCBI database using BLAST program (http://www.ncbi.nlm.nih.gov/BLAST/) [23].
N-terminal sequencing of the purified protease was performed at The Institute of Microbial Technology (IMTECH), Chandigarh. The sequences obtained were subjected to pBLAST, sequence alignment was done by ClustalW and a phylogenetic tree was constructed by neighbour-joining method using MEGA5.
Characterization of the purified protease
Optimization of assay conditions for protease activity:
Firstly, buffer conditions for optimal activity of the puri- fied protease were determined. Enzyme assays were per- formed with 0.1 M concentration of different buffers:
citrate (pH 4−6), potassium phosphate (pH 6−8), acetate (pH 4−6) and Tris-HCl (pH 7−9). The buffer in which highest protease activity was recorded was then selected for optimization of molarity (25 mM to 200 mM).
The optimum reaction temperature was selected by performing assays at different temperatures (25℃ to 50℃) and recording the protease activity. Similarly, thermostability studies were done by incubating the enzyme at temperatures varying from 30℃ to 60℃ for 6 h and noting the effect on protease activity after every hour. The half life of an enzyme is the time taken for its activity to reduce to half of the original activity. Half life of the purified protease was calculated by storing it at 4℃ and recording its activity every day.
Reaction time (2.5 min to 60 min) and substrate con- centration (0.1% to 0.6% of casein) were optimized in a similar manner. At each optimization step only one of the parameters was varied while keeping all others con- stant.
Effect of metal ions on the protease activity: The study of effect of different metal ions was undertaken to find which ones act as stimulators or inhibitors of the prote- ase activity. Enzyme assays were performed with the purified protease in the presence of 2 mM concentration of different metal ions (CoCl2, FeCl2, MnCl2· 4H2O, CuSO4· 5H2O, FeSO4· 7H2O, MnSO4· H2O, ZnSO4· 7H2O, MgSO4· 7H2O and CaCl2· 2H2O). Taking the enzyme activity under normal assay conditions as control, the effect of each metal ion on the protease activity was recorded.
Effect of oxidizing and bleaching agents on protease activity: For this study the purified protease of S. marc- escens PPB-26 was incubated with different concentra- tions (5 to 30% v/v) of hydrogen peroxide and sodium hypochlorite for 1 h at 30℃. Assays were then performed in each case and residual activity calculated, taking activity in the control as 100%.
Effect of detergents and surfactants on protease activ- ity: The effect of different non-ionic (Tween-20, Tween- 60, Triton X-100) and commercial detergents (Ariel and Tide) on S. marcescens PPB-26 protease activity was studied. The enzyme was pre-incubated with different detergents (5% v/v of 10 mg/ml) at 30℃ and its residual
activity in each case recorded after 30 min and 60 min intervals taking the activity in control as 100%. The detergent with which highest protease activity was recorded was then selected for the wash performance analysis.
Wash performance analysis: For this 1cm x 1cm pieces of cotton fabric were soaked in blood and dried. These stained fabrics were then subjected to three separate wash treatments at 30℃ for 5 min. Treatment 1: puri- fied protease (10 U/ml), Treatment 2: selected detergent (5% v/v of 10 mg/ml), Treatment 3: equal volumes of both protease and the detergent. Effects of the different treat- ments were analysed visually, taking unstained and stained but untreated fabric pieces as control.
Results
Purification of S. marcescens PPB-26 protease
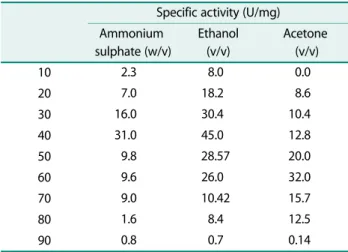
Of the three different fractionation methods used, highest protease activity in the precipitates was observed with 40% ethanol saturation (45 U/mg protein), followed by 60% acetone saturation (32 U/mg protein) and 40% ammonium sulphate saturation (31 U/mg pro-
tein) (Table 1). Large scale precipitation was thus car- ried out using ethanol fractionation and subjected to column chromatography. The active fractions showing protease activity were obtained with 0.6 M NaCl elution.
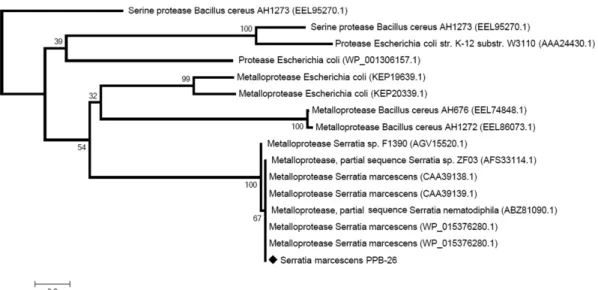
These fractions were pooled and subjected to SDS-PAGE and Native-PAGE analysis which revealed that the purified protease of S.marcescens PPB-26 is a 50 kDa monomeric protein (Fig. 1). Results of the protease puri- fication are summarised in Table 2. MALDI-TOF analy- sis and N-terminal sequencing of the protease showed that it had similarities with numerous previously reported metalloproteases of Serratia marcescens. By aligning its homologous sequences a phylogenetic tree (Fig. 2) and a jalview of the S. marcescens PPB-26 puri- fied protease were generated (Fig. 3).
Optimization of assay conditions for protease activity Buffer optimization showed that the purified protease was active between pH 5 (24.5 U/mg protein) and pH 9 (73 U/mg protein) showing maximum activity (76.4 U/
mg protein) at pH 7.5 with 0.15 M Tris-HCl buffer (Fig.
Table 1. Fractionation of protese of S. marcescens PPB-26.
Specific activity (U/mg) Ammonium
sulphate (w/v)
Ethanol (v/v)
Acetone (v/v)
10 2.3 8.0 0.0
20 7.0 18.2 8.6
30 16.0 30.4 10.4
40 31.0 45.0 12.8
50 9.8 28.57 20.0
60 9.6 26.0 32.0
70 9.0 10.42 15.7
80 1.6 8.4 12.5
90 0.8 0.7 0.14
Fig. 1. SDS-PAGE of DEAE-Cellulose chromatography fraction having protease activity loaded in increasing concentra- tions and run against the marker. Lane 1-Marker, Lane 2 to 6- increasing concentrations of purified protease of S. marcescens PPB-26.
Table 2. Purification summary of S. marcescens PPB-26 protease.
Purification step
Volume (ml)
Protein (mg/ml)
Protein (mg)
Activity (U)
Specific activity
(U/mg) % Yield Fold
purification
Crude 500 0.24 120 2400 20.0 100 1.0
30-40% ethanol 10.0 1.2 12.0 840 45.0 28.0 2.25
Pure 5.0 1.1 5.5 419.1 76.2 20.0 3.81
4A, 4B). Optimal temperature for its activity was found to be 30℃, though it retained 94% of its activity at 50℃.
Thermostability studies further showed that the prote- ase can tolerate 30℃ and 40℃ temperatures for over 6 h while at higher temperatures (50℃ and 60℃) it loses approximately 75% of its activity after 1 h incubation fol- lowed by a steady decline in activity, approaching zero at the end of 6 h incubation (Fig. 4C). The protease had a half life of 15 days at 4℃.
Optimum reaction time of the purified metalloprote- ase was 10 min while the optimum substrate concentra- tion was found to be 0.25% casein (76.5 U/mg protein).
The enzyme followed a normal Lineweaver-Burk plot up to 0.3% casein (Fig. 4D). Analysis of the plot showed that the purified protease had Km = 0.28% and Vmax = 111.11 µmoles mg-1 min-1 protein.
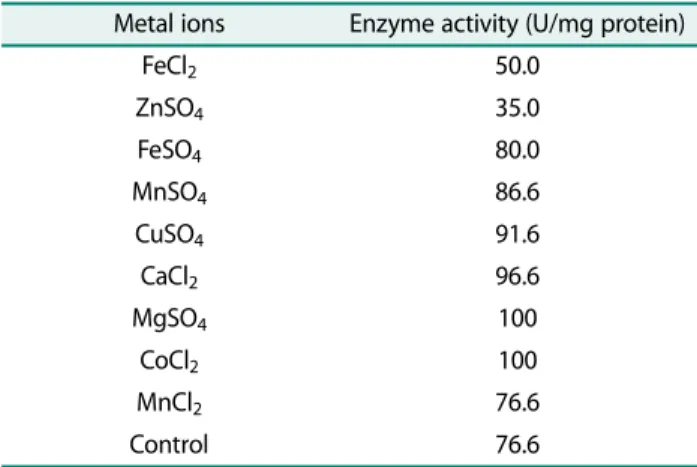
Effect of metal ions on protease activity
The effects of various metals were examined in order to find which ions are stimulators and which are inhibi- tors of the catalytic process. The protease was stable in most metal ions showing increased activity with MgSO4 (30.54% increase), CoCl2 (30.54% increase), CaCl2 (26%
increase), CuSO4 (19.6% increase) and MnSO4 (13%
increase). MnCl2 had no effect on its activity while ZnSO4 and FeCl2 reduced the activity by 34.7% and 54.3% respectively (Table 3).
Effect of oxidizing and bleaching agents on protease activity
The protease of S. marcescens PPB-26 showed toler- ance towards high concentrations of oxidizing and bleaching agents over a period of 1 h. At 5% concentra- Fig. 2. Phylogenetic tree from the protein sequence of S. marcescens PPB-26.
Fig. 3. Jalview alignment of N-terminal sequence of protease of S. marcescens PPB-26 with Serratia metalloprotease (gi 157833779, gi 3114529, gi 1431808, gi 418205677, gi 167487383, gi 672090854, gi 157833854, gi 218198433, gi 47591).
tion of hydrogen peroxide and sodium hypochloride, the protease retained 15% and 20% of its original activity respectively. While at 15% concentration the activity was reduced to 10% and 12% respectively. The activity was lost completely when incubated with concentrations higher than 15% (Table 4).
Compatibility of the protease with detergents and surfac- tants
The purified protease of S. marcescens PPB-26 was Table 3. Effect of metal ions on S. marcescens PPB-26 prote-
ase activity.
Metal ions Enzyme activity (U/mg protein)
FeCl2 50.0
ZnSO4 35.0
FeSO4 80.0
MnSO4 86.6
CuSO4 91.6
CaCl2 96.6
MgSO4 100
CoCl2 100
MnCl2 76.6
Control 76.6
Table 4. Effect of detergents/surfactants and oxidizing/
bleaching agents on S. marcescens PPB-26 protease activity.
Detergents/surfactants (5% v/v of 10 mg/ml)
Relative activity %
30 min 60 min
Tween 20 96 93
Tween 80 94 93
Triton X-100 100 89
Ariel 106 100
Tide 104 102
Oxidizing/bleaching agent concentration (% v/v)
Relative activity %
H2O2 Sodium hypochloride
5 15 20
10 14 18
15 10 12
20 0 10
25 0 0
30 0 0
Fig. 4. (A) Effect of pH on activity of purified protease of S. marcescens PPB-26, (B) Effect of buffer molarity on purified pro- tease of S. marcescens PPB-26, (C) Thermostability of protease of S. marcescens PPB-26, (D) Lineweaver-Burk plot for purified protease of S. marcescens PPB-26.
compatible with different detergents and surfactants retaining almost all of its activity even after 1 h incuba- tion (Table 4). The protease showed greater stability in the commercial detergents (100 to 106% activity) as com- pared to non-ionic detergents (89 to 93% activity). With Ariel however, the protease activity was the highest (106%) after 30 min incubation, it was therefore selected for the further wash performance studies.
Wash performance analysis of the protease
The wash performance analysis showed that S.marc- escens PPB-26 protease has the ability of removing pro- tein stains from fabrics (Fig. 5). Stain removal using the protease treatment gave better results than the Ariel treatment. However, when treated with equal volumes of both the purified protease and the detergent Ariel, the stain removal was more efficient and the cleaning time was reduced to half (2 min) the initial time.
Discussion
The largest application of proteases is in the detergent industry as detergent additives [24]. A majority of these proteases come from the genus Bacillus [25] and there is an increasing demand for new alternative sources of pro- teases with novel properties. Further, with the increas- ing environment and energy constrains there is a growing need for enzymes that act at normal tempera- tures and neutral pH. In this respect the present research work was undertaken with a focus on exploring the biotechnological potential of S. marcescens PPB-26 protease in the detergent industry. The enzyme was purified to homogeneity and characterized for novel properties that support its application as a detergent
additive. Protein precipitation was best achieved with ethanol fractionation which is similar to the behaviour of Bacillus sp. APR-4 protease [26] and Streptomyces sp.
XE-1 protease [27]. Organic solvent fractionation is advantageous over salt precipitation, since a change in density of the solution upon addition of organic solvent causes the precipitates to settle down more readily thus giving a product of higher purity and activity [28]. Pre- cipitates from ethanol fractionation were dissolved in Tris-HCl buffer (pH 9) and subjected to column chroma- tography for further purification and active fractions were obtained with 0.6 M NaCl elution. This suggests that the enzyme has a high negative charge over its sur- face at pH 9. The final purification of the protease was 3.8 fold with 20% yield which is much higher than the previously reported 2.1 fold purification and 4.6% yield of B. brevis MWB-01 serine metalloprotease [29] and 1.7 fold purification and 7.4% yield of Planomicrobium sp.
protease [30]. The protease of S. marcescens PPB-26 is a neutral protease showing optimal activity at neutral pH 7.5 of 0.15 M Tris-HCl buffer, a trend similar to the pro- tease of S. marcescens S3-R1 [31] and the S. marcescens subsp. sakuensis TKU019 protease [32]. The neutral pro- teases are highly preferred in food industries since they cause less bitterness in food protein hydrolysates by hydrolyzing hydrophobic amino acid bonds at neutral pH [33]. Neutral and alkaline proteases are also pre- ferred in detergent and leather tanning industries due to an increasing trend of developing environment friendly technologies [8]. The optimal temperature for protease activity was found to be 30℃ which is similar to that of S. rubidaea protease [34] and S. marcescens S3-R1 pro- tease [31]. Since the protease is hydrophobic, as observed during the elution profile in DEAE chromatog- Fig. 5. Wash performance analysis of S. marcescens PPB-26 protease. (A) Unstained fabric, (B) Blood stained fabric, (C) Ariel treated stained fabric after 5 min, (D) Protease treated stained fabric after 5 min, (E) Stained fabric treated with protease and Ariel after 2 min.
raphy, an increase in temperature beyond 40℃ causes breaking of hydrogen bonds and non-polar hydrophobic interactions which results in loss of enzyme activity.
Analysis of the Lineweaver-Burk plot showed that the protease had Km = 0.28% (1.36 mM) and Vmax = 111.11 µmoles mg-1 min-1 protein. A fibrinolytic protease from Bacillus megaterium with Km and Vmax as 0.61 mM and 10.2 µmoles mg-1min-1 respectively has been reported previously [35] whereas an acidic protease from Aspergillus oryzae has the Km and Vmax of 0.12 mM and 14µmoles min-1 respectively [36]. The protease of S.mar- cescens PPB-26 is stable in the presence of metal ions (2 mM). It showed increased activity with MgSO4, CaCl2, CoCl2, CuSO4 and MnSO4 whereas MnCl2 had no effect and ZnSO4 and FeCl2 slightly inhibited its activ- ity. This is an importance property for a detergent addi- tive since a detergent should be equally capable of removing stains in hard or heavy metal rich water as it does in soft water. Ca2+ is a known activator of the prote- ases. It acts by stabilising the enzyme in its active con- formation therefore having a positive effect on its activity [13, 37, 38]. However, S. marcescens subsp.
sakuensis protease is reported to be inhibited by all diva- lent metal ions (Mg2+, Cu2+, Fe2+, Ca2+, Zn2+, Mn2+) [32].
The study of effect of metal ions on protease activity has been done in several other previously reported works [12, 13, 36, 39, 40]. Stability and compatibility studies of the protease with different detergents showed that the protease retained 93% of its activity in non ionic deter- gents which is much higher than 60% to 70% activity in case of B. cereus TKU006 metalloprotease [10] and 20%
to 60% activity in Aeribacillus. pallidus C10 protease [41]. Besides this, 100% of its activity was retained with commercial detergents (Ariel and Tide). These results are different from reports of B. brevis MWB-01 protease whose activity was inhibited by Ariel [29] and from a Bacillus sp. SSR1 protease which retained 37% of its ini- tial activity after 1 h incubation with 5 mg/ml Ariel [42].
The purified protease of S. marcescens PPB-26 was also more effective in blood stain removal from fabric than the commercial detergent Ariel. Further, supplementa- tion of Ariel with the protease enhanced its efficiency and also reduced the cleaning time to half. Several stud- ies on synergistic effects of protease and detergent have been reported in the past [4, 43]. Greene [44] had reported that enzyme supplemented detergent was con-
siderably more effective at removing the soiling agents (blood, milk and carbon black) than the control. Oxidiz- ing and bleaching agents are the common components of the modern day laundry detergents. The protease of S.
marcescens PPB-26 was tolerant to the oxidizing and bleaching agents over a period of 60 min, retaining 15%
and 20% of its activity respectively at 5% concentration.
The protease from Bacillus mojavensis A21 has been reported to retain 70% and 38% activity on 60 min incu- bation with 1% concentration of hydrogen peroxide and sodium perborate respectively [13]. The cleansing for- mulations are often supplemented with oxidizing and bleaching agents to enhance the brightness or to steril- ize the washed material [22, 44, 45], therefore stability of the purified enzyme in oxidizing and bleaching agents is important for eventual use in laundry formulations.
In conclusion, the purified metalloprotease of S.
marcescens PPB-26 has several novel properties. It is optimally active at a neutral pH and at room tempera- ture which makes it an environment friendly option for application in industries. The positive effect of most metal ions on its activity is a rare and unique finding.
The protease is stable in surfactants and is also tolerant to high concentrations of bleaching and oxidizing agents.
It is capable of removing stains from fabrics and it greatly increases the efficiency of commercial detergents when added to them. All these findings make the prote- ase of S. marcescens PPB-26 an excellent candidate for application in the detergent industry.
Acknowledgments
We acknowledge Council of Scientific and Industrial Research (CSIR) New Delhi, India, for financial support in the form of SRF to Shikha Thakur. We are also thankful to computational facility at Sub Distrib- uted Information Centre, Himachal Pradesh University, Shimla- 171005, India. Special thanks to School of Life Sciences, JNU, New Delhi, India and IMTECH, Chandigarh, India for their help.
Conflict of Interest
The authors have no financial conflicts of interest to declare.
References
1. Shankar S, Rao M, Laxman RS. 2011. Purification and characteri- zation of an alkaline protease by a new strain of Beauveria sp.
Process Biochem. 46: 579-585.
2. Joanitti GA, Freitas SM, Silva LP. 2006. Proteinaceous protease inhibitors: structural features and multiple functional faces. Curr.
Enzym. Inhib. 2: 199-217.
3. Sarmiento F, Peralta R, Blamey JM. 2015. Cold and hot extremo- zymes: industrial relevance and current trends. Front. Bioeng. Bio- technol. 3: 148.
4. Bouacem K, Bouanane-Darenfed A, Laribi-Habchi H, Elhoul MB, Hmida-Sayari A, Hacene H, et al. 2015. Biochemical characteriza- tion of a detergent-stable serine alkaline protease from Caldico- probacter guelmensis. Int. J. Biol. Macromol. 81: 299-307.
5. Bhalla T, Kumar D, Gajju H, Agrawal H. 1999. Thermophilic bacte- rial proteases. J. Punjab Acad. Sci. 1: 77-91.
6. Salem S, Shabeb M, Amara A. 2009. Optimization of thermophilic protease production in Bacillus mixed cultures under mesophilic conditions. World J. Agric. Sci. 5: 375-383.
7. Padmapriya B, Rajeswari T, Nandita R, Raj F. 2012. Production and purification of alkaline serine protease from marine Bacillus spe- cies and its application in detergent industry. Eur. J. Appl. Sci. 4:
21-26.
8. Rao MB, Tanksale AM, Ghatge MS, Deshpande VV. 1998. Molecu- lar and biotechnological aspects of microbial proteases. Micro- biol. Mol. Biol. Rev. 62: 597-635.
9. Jisha VN, Smitha RB, Pradeep S, Sreedevi S, Unni KN, Sajith S, et al.
2013. Versatility of microbial proteases. Adv. Enzyme Res. 1: 39-51.
10. Wang SL, Chao CH, Liang TW, Chen CC. 2009. Purification and characterization of protease and chitinase from Bacillus cereus TKU006 and conversion of marine wastes by these enzymes.
Mar. Biotechnol. 11: 334-344.
11. Bajpai D. 2007. Laundry detergents: an overview. J. Oleo Sci. 56:
327-340.
12. Nadeem M, Qazi JI, Syed Q, Gulsher M. 2013. Purification and characterization of an alkaline protease from Bacillus licheni- formis UV-9 for detergent formulations. Songklanakarin J. Sci.
Technol. 35: 187-195.
13. Haddar A, Agrebi R, Bougatef A, Hmidet N, Sellami-Kamoun A, Nasri M. 2009. Two detergent stable alkaline serine-proteases from Bacillus mojavensis A21: purification, characterization and potential application as a laundry detergent additive. Bioresour.
Technol. 100: 3366-3373.
14. Hasan F, Shah AA, Javed S, Hameed A. 2010. Enzymes used in detergents: lipases. Afr. J. Biotechnol. 9: 4836-4844.
15. Grbavčić S, Bezbradica D, Izrael-Živković L, Avramović N, Milosavić N, Karadžić I, et al. 2011. Production of lipase and prote- ase from an indigenous Pseudomonas aeruginosa strain and their evaluation as detergent additives: compatibility study with detergent ingredients and washing performance. Bioresour.
Technol. 102: 11226-11233.
16. Hmidet N, Ali NE-H, Haddar A, Kanoun S, Alya S-K, Nasri M. 2009.
Alkaline proteases and thermostable α-amylase co-produced by Bacillus licheniformis NH1: Characterization and potential appli- cation as detergent additive. Biochem. Eng. J. 47: 71-79.
17. Gupta R, Beg Q, Lorenz P. 2002. Bacterial alkaline proteases:
molecular approaches and industrial applications. Appl. Micro-
biol. Biotechnol. 59: 15-32.
18. Kumar CG, Takagi H. 1999. Microbial alkaline proteases: from a bioindustrial viewpoint. Biotechnol. Adv. 17: 561-594.
19. Mei C, Jiang X. 2005. A novel surfactant-and oxidation-stable alkaline protease from Vibrio metschnikovii DL 33-51. Process Bio- chem. 40: 2167-2172.
20. Bhaskar N, Sudeepa E, Rashmi H, Selvi AT. 2007. Partial purifica- tion and characterization of protease of Bacillus proteolyticus CFR3001 isolated from fish processing waste and its antibacterial activities. Bioresour. Technol. 98: 2758-2764.
21. Thakur S, Sharma NK, Thakur N, Savitri, Bhalla TC. 2016. Organic solvent tolerant metallo protease of novel isolate Serratia marc- escens PPB-26: production and characterization. 3 Biotech. 6: 180.
22. Manachini PL, Fortina MG, Parini C. 1988. Thermostable alkaline protease produced by Bacillus thermoruber—a new species of Bacillus. Appl. Microbiol. Biotechnol. 28: 409-413.
23. Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990. Basic local alignment search tool. J. Mol. Biol. 215: 403-410.
24. Kumar D, Savitri TN, Verma R, Bhalla TC. 2008. Microbial prote- ases and application as laundry detergent additive. Res. J. Micro- biol. 3: 661-672.
25. Samal BB, Karan B, Stabinsky Y. 1990. Stability of two novel serine proteinases in commercial laundry detergent formulations. Bio- technol. Bioeng. 35: 650-652.
26. Kumar D, Bhalla T. 2004. Purification and characterization of a small size protease from Bacillus sp. APR-4. Indian J. Exp. Biol. 42:
515-521.
27. Xu Y, Lu Y, Xing Y, Xi T. 2015. Characterization of solvent, deter- gent and oxidizing agent stable protease from isolated Antarctic marine Streptomyces sp. XE-1. Wei Sheng Wu Xue Bao. 55: 311- 320.
28. Moon SH, Parulekar SJ. 1991. A parametric study ot protease pro- duction in batch and fed‐batch cultures of Bacillus firmus. Bio- technol. Bioeng. 37: 467-483.
29. Olajuyigbe FM, Falade AM. 2014. Purification and partial charac- terization of serine alkaline metalloprotease from Bacillus brevis MWB-01. Bioresour. Bioprocess 1: 8.
30. Liu Q, Sun S, Piao M, Yang JY. 2013. Purification and Characteriza- tion of a Protease Produced by a Planomicrobium sp. L-2 from Gut of Octopus vulgaris. Prev. Nutr. Food Sci. 18: 273-279.
31. Nam MS, Whang KS, Choi SH, Bae HC, Kim YK, Park YW. 2013.
Purification, characterization, and properties of an alkaline prote- ase produced by Serratia marcescens S3‐R1 inhabiting Korean ginseng rhizosphere. J. Sci. Food Agric. 93: 3876-3882.
32. Liang TW, Kuo YH, Wu PC, Wang CL, Dzung NA, Wang SL. 2010.
Purification and characterization of a chitosanase and a protease by conversion of shrimp shell wastes fermented by Serratia Marc- escens Subsp. Sakuensis TKU019. J. Chin. Chem. Soc. 57: 857-863.
33. Sandhya C, Sumantha A, Szakacs G, Pandey A. 2005. Compara- tive evaluation of neutral protease production by Aspergillus oryzae in submerged and solid-state fermentation. Process Bio- chem. 40: 2689-2694.
34. Doddapaneni KK, Tatineni R, Vellanki RN, Gandu B, Panyala NR,
Chakali B, et al. 2007. Purification and characterization of two novel extra cellular proteases from Serratia rubidaea. Process Bio- chem. 42: 1229-1236.
35. Kotb E. 2015. Purification and partial characterization of serine fibrinolytic enzyme from Bacillus megaterium KSK-07 isolated from kishk, a traditional Egyptian fermented food. Appl. Biochem.
Microbiol. 51: 34-43.
36. Yin L-J, Chou Y-H, Jiang S-T. 2013. Purification and characteriza- tion of acidic protease from Aspergillus oryzae BCRC 30118. J.
Marine Sci. Technol. 21: 105-110.
37. Hmidet N, Ali NE-H, Haddar A, Kanoun S, Alya SK, Nasri M. 2009.
Alkaline proteases and thermostable α-amylase co-produced by Bacillus licheniformis NH1: Characterization and potential appli- cation as detergent additive. Biochem. Eng. J. 47: 71-79.
38. Annamalai N, Rajeswari MV, Balasubramanian T. 2014. Extraction, purification and application of thermostable and halostable alkaline protease from Bacillus alveayuensis CAS 5 using marine wastes. Food Bioproducts Processing. 92: 335-342.
39. Doddapaneni KK, Tatineni R, Vellanki RN, Rachcha S, Anabrolu N, Narakuti V, et al. 2009. Purification and characterization of a sol- vent and detergent-stable novel protease from Bacillus cereus.
Microbiol. Res. 164: 383-390.
40. Chandrasekaran M, Chandrasekar R, Chun SC, Sathiyabama M.
2016. Isolation, characterization and molecular three-dimen- sional structural predictions of metalloprotease from a phyto- pathogenic fungus, Alternaria solani (Ell. and Mart.) Sor. J. Biosci.
Bioeng. 122: 131-139.
41. Yildirim V, Baltaci MO, Ozgencli I, Sisecioglu M, Adiguzel A, Adiguzel G. 2017. Purification and biochemical characterization of a novel thermostable serine alkaline protease from Aeribacillus pallidus C10: a potential additive for detergents. J. Enzyme Inhib.
Med. Chem. 32: 468-477.
42. Singh J, Batra N, Sobti R. 2001. Serine alkaline protease from a newly isolated Bacillus sp. SSR1. Process Biochem. 36: 781-785.
43. Jaouadi B, Ellouz-Chaabouni S, Ali MB, Messaoud EB, Naili B, Dhouib A, et al. 2009. Excellent laundry detergent compatibility and high dehairing ability of the Bacillus pumilus CBS alkaline proteinase (SAPB). Biotechnol. Bioprocess Eng. 14: 503-512.
44. Greene RV, Griffin HL, Cotta MA. 1996. Utility of alkaline protease from marine shipworm bacterium in industrial cleansing applica- tions. Biotechnol. Lett. 18: 759-764.
45. Sanatan PT, Lomate PR, Giri AP, Hivrale VK. 2013. Characterization of a chemostable serine alkaline protease from Periplaneta amer- icana. BMC Biochem. 14: 32.