23(2) : 92-96 (2017)
https://doi.org/10.20307/nps.2017.23.2.92
92
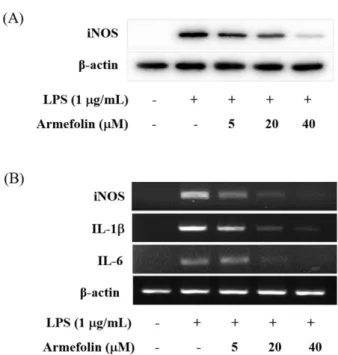
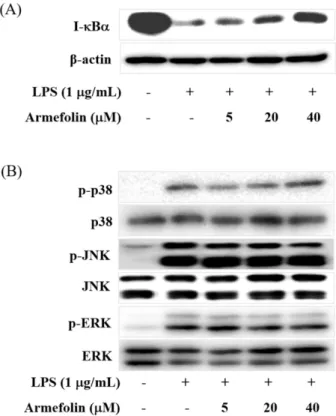
Inhibitory Effect of a Sesquiterpene from Artemisia iwayomogi on Expression of Inducible Nitric Oxide Synthase by Suppression of I- κBα Degradation
in LPS-stimulated RAW 264.7 Cells
Na Yeon Kim
1, Hye Jin Koh
1, Hua Li
1, Hwa Jin Lee
2,*, and Jae-Ha Ryu
1,*
1
College of Pharmacy, Sookmyung Women’s University, Seoul 04310, Republic of Korea
2