39(2) : 110 117 (2008)
110
Salidroside 의 RAW 264.7 세포에서 NF-κB 불활성화를 통한 LPS에 의해 유도되는 iNOS, COX-2 및 cytokine들의 발현 저해효과
원소정
1· 박희준
2· 이경태
1*
1
경희대학교 약학대학 ,
2상지대학교 자원식물학과
Inhibition of LPS induced iNOS, COX-2 and cytokines expression by salidroside through the NF- κB inactivation in RAW 264.7 cells
So-Jung Won
1, Hee-Juhn Park
2, Kyung-Tae Lee
1*
1
Department of pharmaceutical Biochemistry, College of Pharmacy, Kyung-Hee University, Seoul 130-701, South Korea
2
Department of Botanical Resources, Sangji University, Woosan-Dong, Wonju 220-702, South Korea
Abstract − In this study, we investigated the anti-inflammatory effects of salidroside (SAL) isolated from the MeOH extract of Acer tegmentosum Maxim heartwood in RAW 264.7 macrophage cells. SAL pretreatment significantly inhibited nitric oxide (NO) and prostaglandin E
2(PGE
2) productions in the lipopolysaccharide (LPS)-induced RAW 264.7 cells. Western blot and RT-PCR analyses revealed that SAL inhibited the LPS-induced expressions of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) at the protein and mRNA levels in a concentration-dependent manner. In addition, SAL reduced the release and the mRNA expressions of tumor necrosis factor- α (TNF- α ) and interleukin-6 (IL-6). Furthermore, nuclear factor- kappa B (NF- κ B) luciferase reporter assay was performed to know the involvement of SAL in the production of pro-inflam- matory cytokines, we confirmed that LPS-induced transcription activity of NF- κ B was inhibited by SAL. Taken together, our data indicate that anti-inflammatory property of salidroside might be the result from the inhibition of iNOS, COX-2, TNF- α and IL-6 expressions via the down-regulation of NF- κ B activity.
Key words − salidroside, Acer tegmentosum Maxim, LPS, NF- κ B, Anti-inflammation
선천적 면역은 박테리아나 바이러스 감염 , 스트레스에 대
해 방어할 수 있도록 다양한 작용기전을 통해 빠르게 활성화되는 고전적 숙주 방어 형태이다 . 선천적 면역
반응이 결여되면 활성산소 (reactive oxygen species),
보체 (complement) 그리고 항미생물펩티드 (anti-
microbial peptides) 의 생성이 저해되어 감염에 민감해
지고 방어능력이 떨어진다 . 그러나 과도한 선천적 면
역반응은 오히려 조직 손상을 촉진하여 패혈증이나 만 성 염증과 같은 질환을 유발한다 .
1)염증과정 중에 많은 양의 염증유도 사이토카인 (pro- inflammatory cytokines), nitric oxide (NO) 그리고 prostaglandin E
2(PGE
2) 가 inducible nitric oxide synthase (iNOS) 와 cyclooxygenase-2 (COX-2) 에 의해 생성된다 .
2)포
유 세포에서 NO 는 neuronal NOS (nNOS), endothelial
NOS (eNOS) 그리고 iNOS 세가지 형태의 NOS 에 의해 합
성된다 . nNOS 와 eNOS 는 세포 내에 항상 존재하지만 , iNOS
는 interferon- γ , lipopolysaccharide (LPS) 그리고 다양한 염
증유도 사이토카인에 노출되는 경우에만 발현된다 .
3-4)이들 NOS 중 iNOS 에 의한 NO 생성이 절대적으로 많으며 이는
병리적으로 중요한 작용을 한다 . 일반적인 NO 형성은 박테
리아를 죽이거나 종양을 제거하는 중요한 역할을 하지만 , 병
리적인 원인에 의한 과도한 NO 형성은 염증을 유발시켜 조
직의 손상 , 유전자 변이 및 신경손상 등을 일으킨다 .
5-7)PGE
2는 cyclooxygenase (COX) 에 의해 arachidonic acid 로부터 생
성되는데 ,
8)이때 작용하는 효소인 COX 는 1990 년대 초에 두
가지 형태로 존재함이 밝혀졌다 . COX-1 은 거의 모든 조직
에서 항상 발현되며 prostaglandins (PGs) 을 형성하여 위세
포 보호와 신혈류 조절 등의 생리적인 기능을 수행한다 .
8-9)반면에 COX-2 는 대식세포와 활막세포 (synoviocyte) 같은 염
증세포에서 감염 , 상처 또는 다른 스트레스에 의해 유도되
*교신저자(E-mail):[email protected] (FAX):02-966-3885
며 많은 양의 PGs 를 생성하여 침해수용체 (nociceptor) 를 활
성화시키고 염증상태로 유도한다 .
10-11)또한 PGs 는 새로운
혈관 생성을 유도하고 세포사멸을 억제함으로써 종양생성 에 기여한다 .
12)Tumor necrosis factor- α (TNF- α ) 는 활성화된 대식세포 ,
섬유모세포 (fibroblast) 및 다른 여러 세포에서 생성되는데
이는 종양세포에 영향을 미치는 숙주 방어인자 및 염증 매 개물질로 알려져 있다 . Interleukin-6 (IL-6) 의 생성은 TNF- α
나 IL-1 β 같은 요인 외에도 LPS 에 의해 유도된다 .
13)또한 , IL-6 는 염증유도 사이토카인으로서 내인성 발열인자 (pyrogen)
로 작용하며 면역 체계와 조혈 등에 다양한 영향을 미친다 .
14)Nuclear transcription factor-kappa B (NF- κ B) 는 세포 분
화 , 염증반응 , 세포 부착 등에 관련된 여러 유전자들의 발
현에 가장 중요한 역할을 하는 전사인자이다 . 활성화된 NF-
κ B 는 iNOS, COX-2, TNF- α 그리고 IL-6 등 여러 염증 매
개물질들의 전사를 촉진한다 .
15)산겨릅나무 ( Acer tegmentosum Maxim) 는 단풍나무과에
속하는 식물로서 고산지대의 계곡에서 자라는 낙엽소교목 이다 .
16)중국에서는 줄기를 칭해척이라 하여 소종과 외상출
혈 치료에 사용하고 , 한국에서는 최근에 벌나무 또는 산청
목이라 하여 주로 간암 , 간염 , 간경화 등의 치료제로 민간
에서 널리 유통되고 있다 .
17)이 나무에 주된 성분을 이루고
있는 salidroside (SAL) 는 amyloid- β에 의한 산화적 스트레
스에 대한 신경세포보호작용 ,
18)지질 과산화 억제 ,
19)bone marrow cell cycle 에 미치는 영향 ,
20)carbohydrate 대사에 미
치는 영향
21)등 다양한 약리학적 활성을 나타낸다는 것이 보고되어있다 . 그러나 SAL 에 대한 항염효과와 그 기전에
대한 보고는 거의 없는 실정이다 .
본 연구진은 여러 천연물의 추출물 및 분리된 단일화합물 들의 항염증 효과를 검색하고 그 기전을 연구함으로써 새 로운 항염증 약물의 개발을 시도하고 있다 . 본 논문에서는
이러한 연구의 일환으로 산겨릅나무로부터 분리한 salidroside
가 LPS 에 의해 활성화된 RAW 264.7 세포에서 항염효과를
나타내는 기전을 연구하였으며 염증에 관련된 다양한 단백
질 , mRNA 그리고 사이토카인 등의 발현을 측정하였다 .
재료 및 방법
재료 − Dulbecco’s modified Eagle’s minimum essential medium (DMEM), fetal bovine serum (FBS), penicillin, streptomycin 은 life Technologies Inc. (Grand Island, NY)
에서 구입하였다 . 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl- tetrazolium bromide (MTT), dimethyl sulfoxide (DMSO), sulfanilamide, aprotinin, leupeptin, phenylmethylsulfonylfluride (PMSF), dithiothreitol (DTT), L-N
6-(1-iminoethyl)lysine (L- NIL), NS-398, Escherichia coli lipopolysaccharide (LPS) 는
Sigma Chemical Co. (CA, U.S.A.) 에서 구입하였으며 , COX-2 와 iNOS monoclonal antibodies 및 peroxidase conjugated secondary antibody 는 Santa Cruz Biotechnology (CA, U.S.A.) 에서 구입하였다 . iNOS, COX-2, TNF- α , IL-6
그리고 β -actin oligonucleotide primers 는 Bioneer (Seoul, Korea) 에서 구입하였다 . 그리고 TNF- α , IL-6, PGE
2측정을 위한 kit 는 R&D systems (MN, U.S.A.) 에서 구입하였다 .
시료의 추출 및 분리
산겨릅나무 목부 2.0 kg 을 MeOH 로 환류 하에 5 시간씩 3
회 추출하였다 . 추출물을 여과하고 얻어진 여액을 모두 합
쳐 감압농축하여 MeOH 추출물을 얻었다 . MeOH 추출물을
증류수 1L 에 현탁시키고 CHCl
3로 3 회 분획한 다음 농축하
여 CHCl
3분획물을 얻었다 . 남은 수층에 대하여 BuOH 로 3 회 분획한 다음 농축하여 BuOH 분획물을 얻었다 . 남은 수
층을 농축하여 물 분획물을 얻었다 .
BuOH 분획물 10 g 을 silica gel column chromatography
에서 CHCl
3-MeOH 전개용매를 용출용매로 사용하여 9 : 1
로 1 L 용출하다가 6 : 4 로 변경하여 2.5 L 전개하여 소분
획 Ⅰ부터 Ⅶ 까지 얻었다 . 이 중 소분획 Ⅲ 을 acetone 에서
재결정하여 무색분말의 p -hydroxyphenylethyl- O - β -D- glucopyranoside (2.8 g) 을 얻었다 . 이 화합물의 소분획 Ⅵ 과
Ⅶ 을 MeOH 에서 재결정하여 무색결정을 얻었으며 이 화
합물은 TLC,
1H-NMR,
13C-NMR 의 분석에 의하여 sucrose
로 동정되었다 .
Salidroside ( p -hydroxyphenylethyl- O - β -D-glucopyranoside, Fig. 1) : Colorless powder, mp 220
o;
1H-NMR(500 MHz, DMSO-d
6) δ : 4.17 (1H, d, J=7.5 Hz, H-1'), 6.67 (2H, d, J=8.5 Hz, H-2.6), 7.04 (2H, d, J=3,5 Hz);
13C-NMR(125.5 MHz, DMSO-d
6) δ : 35.3 (C-7), 61.5(C-6'), 70.3 (C-8), 70.5(C-4'), 73.8 (C-2'), 77.1 (C-3'), 77.3 (C-5', 103.3 (C-1'), 115.4 (C-2,5), 129.2 (C-1), 130.2 (C-3,5), 155.9 (C-4);
FAB-MS m/z 323.2 [M + Na]
+.
세포의 배양 − RAW 264.7 세포는 10% FBS 및 penicillin (100 µg/mL), streptomycin (100 U/mL) 이 포함된 DMEM 배지에서 37
oC, 5% CO
2incubator 에서 배양했다 . RAW 264.7 세포에 시료용액의 여러 농도 (25, 50, 100 µM)
또는 양성 대조군을 1 시간 전처리한 후 LPS (1 µg/mL) 를
처리하고 24 시간 배양하였다 .
Fig. 1. Chemical structure of salidroside.
세포독성 시험 − 96 well plate 에 1×10
5cells/well 로 세포
를 동일하게 분주하고 24 시간 동안 배양한 후 여러 농도의
시료용액을 두 군으로 나누어 배지에 희석하여 첨가 하였 다 . 1 시간 후 한 군에만 LPS (1 µ g/mL) 를 처리하였다 . 24 시
간이 지난 후 MTT 시약을 넣고 4 시간 동안 방치한 후 상등
액을 제거하고 형성된 formazan 을 DMSO 100 µL 를 첨가하
여 녹였다 . 30 분 후 540 nm 에서 흡광도를 측정하였다 .
Nitrite 양의 측정 − RAW 264.7 세포로부터 생성된 NO
의 양은 Griess 시약을 이용하여 세포 배양액 중에 존재하
는 NO
2−의 형태로서 측정하였다 . 즉 세포배양 상등액 100 µL
와 Griess 시약 [1% (w/v) sulfanilamide in 5% (v/v) phosphoric acid 와 0.1% (w/v) naphtylethylenediamine- HCl] 100 µL 를 혼합하여 96 well plates 에서 10 분 동안 반
응시킨 후 540 nm 에서 흡광도를 측정하였다 .
Western blot 시험 − SAL 을 처리한 세포 및 대조군을 phosphate buffered saline (PBS) 로 씻어낸 후 lysis buffer 인 PRO-PREP (Intron Biotechnology) 으로 단백질을 추출한 후
원심분리하여 상등액을 취하였다 . 상등액을 Bradford 시약
을 사용해 단백질 농도를 정량하여 40 µg 의 단백질을 취했
다 . 추출된 단백질은 10% 의 SDS-polyacrylamide gel 에 전
기영동시킨 후 nitrocellulose membrane 으로 gel 의 단백질을 blot 시켰다 . 5% skim milk 로 하루 밤 동안 blocking 한 후 1:500 의 비율로 iNOS 와 COX-2 antibody 를 4 시간 동안 상
온에서 방치한 후 TTBS 로 15 분 간격으로 2 회 세척하였다 . 1:1000 의 비율로 희석한 secondary antibody 를 1 시간 동안
상온에서 방치 시켰다 . 다시 TTBS 로 15 분 간격으로 3 회 세
척한 후 chemiluminescence 로 현상하였다 .
PGE
2, TNF- α 및 IL-6 양의 측정 − 세포배양액을 취해 각각 R&D systems (MN, U.S.A.) kit 의 지시에 따라 PGE
2, TNF- α 및 IL-6 를 정량하였다 .
RT-PCR 시험 − Easy Blue
®kits (Intron Biotechnology)
를 이용하여 Kit 의 protocol 에 따라 전체 cellular RNA 를 추
출하였다 . 각각의 시료에서 MuLV reverse transcriptase, 1 mM dNTP 그리고 oligo (dT
12-18) 0.5 µg/µL 를 이용하여 1 µg 의 RNA 를 역전사 하여 cDNA 를 얻었다 . cDNA 에 Taq
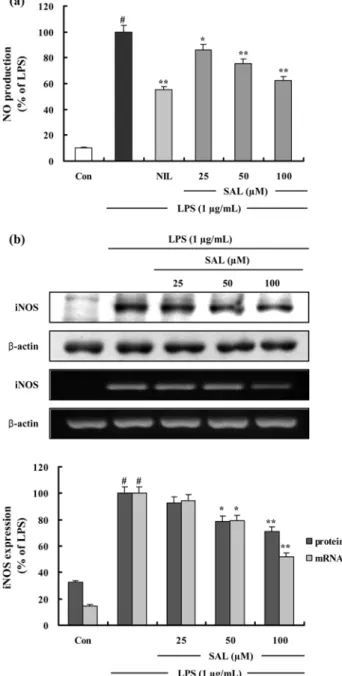
Fig. 2. The effects of SAL on LPS-Induced NO production and iNOS protein and mRNA expression in RAW 264.7 cells. (a) Cells were pretreated with different concentrations (25, 50, 100 µM) of SAL for 1 h, LPS (1 µg/mL) was then added, and the cells were incubated for 24 h. Control (Con) values were obtained in the absence of LPS or tested samples. L-N
6-(1- iminoethyl) lysine (L-NIL) was used as an assay positive control at a concentration of 10 µM. (b) Cells were treated as described above. Total cellular proteins (40 µg) were resolved by SDS-PAGE, transferred to nitrocellulose membranes, and detected with specific antibodies, as described in methods. A representative immunoblot of three separate experiments is shown. Cells were pretreated with different concentrations (25, 50, 100 µM) of SAL for 1 h, LPS (1 µg/mL) was then added, and the cells were incubated for 4 h. Control (Con) values were obtained in the absence of LPS or tested samples. Total
RNA was prepared for the RT-PCR analysis of iNOS gene expression. iNOS-specific sequences (807 bp) was detected by agarose gel electrophoresis, as described in methods. PCR of
β -actin was performed to verify that the initial cDNA contents
of the samples were similar. The experiment was repeated
three times and similar results were obtained. The Western blot
results and the RT-PCR results are shown by relative ratio
graphs. The values are the mean ± S.D. of three independent
experiments.
#p <0.05 vs. the control group; * p <0.05, ** p <0.01
vs. the LPS-treated group; the significances of the difference
between the treated groups was evaluated using the Student’s
t -test.
DNA polymerase 1 unit, 0.2 mM dNTP, ×10 reaction buffer 그리고 5' 와 3' primers 100 pmol 을 포함한 전체 부피 25 µL 의 시료를 thermal cycler (Perkin Elmer Cetus, Foster City, CA, USA) 를 이용하여 PCR 분석을 하였다 . PCR 반
응은 95
oC 에서 2 분간 initial denaturation 시킨 후 iNOS (95
oC 1 분 denaturation, 60
oC 1 분 annealing 그리고 72
oC 1.5 분 extension), COX-2 (94
oC 1 분 denaturaion, 60
oC 1 분 annealing 그리고 72
oC 1 분 extension), TNF- α (95
oC 1 분 denaturation, 55
oC 1 분 annealing 그리고 72
oC 1 분 extension) 그리고 IL-6 (94
oC 1 분 denaturation, 56
oC 1 분 annealing 그리고 72
oC 1 분 extension) 를 30 회 amplification 하였다 . 이번 연구에서 아래의 목록과 같은 PCR primers 가 사용되었다 : sense strand iNOS, 5'-ATT GGC AAC ATC AGG- TCG GCC ATC ACT-3', anti- sense strand iNOS,5'-GCT GTG TGT CAC AGA AGT CTC GAA- CTC-3'; sense strand COX-2, 5'-GGA GAG ACT ATC AAG ATA GT-3' anti-sense strand COX-2, 5'- ATG GTC AGT AGA CTT TTA CA-3'; sense strand TNF- α , 5'-ATG AGC ACA GAA AGC ATG- ATC-3', anti-sense strand TNF- α , 5'-TAC AGG CTT GTC ACT CGA ATT-3'; sense strand IL-6, 5'-GAG GAT ACC ACT CCC AAC AGA CC-3', anti-sense strand IL-6, 5'-AAG TGC ATC ATC GTT GTT CAT ACA-3';5'-GTG CTG CCT- AAT GTC CCC TTG AAT C-3'; sense strand β - actin, 5'-TCA TGA AGT GTG ACG TTG ACA- TCC GT-3', anti-sense strand β -actin, 5'-CCT AGA AGC ATT TGC GGT GCA CGA TG-3'. Amplification 후에 PCR 반
응 시킨 시료를 2% agarose gel 에서 전기영동하고 ethidium bromide 염색과 UV 조사를 통해 확인하였다 .
NF- κ B Luciferase activity 측정 − RAW 264.7 세포를 dish 에 각각 2×10
5cells/dish 농도로 분주한 후 , Invitrogen (CA, U.S.A.) 에서 구입한 Lipofectamine 2000 Reagent 를 이
용하여 NF- κ B luciferase reporter plasmid DNA 를 형질감염 (transfection) 시켰다 . 형질감염 48 시간이 경과한 후 3~4×10
5cell/well 로 12 well plate 에 세포를 분주하고 SAL 를 1 시간
동안 전처리 한 후 LPS (1 µg/mL) 를 처리하였다 . 24 시간 후
세포를 수집하여 luciferase assay system (Promega, U.S.A.)
와 luminometer (Perkin Elmer Cetus, U.S.A) 를 이용하여
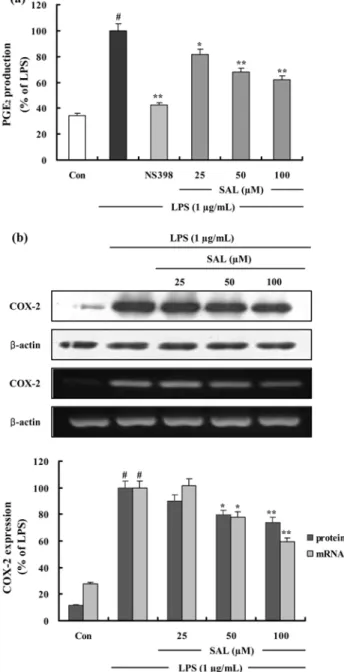
Fig. 3. The effects of SAL on LPS-Induced PGE
2production and COX-2 protein and mRNA expression in RAW 264.7 cells. (a) Cells were treated as described in the Figure Legend of 2a. Control (Con) values were obtained in the absence of LPS or tested samples. 10µM of NS-398 was as a positive control in the assay. (b) Cells were treated as described in the Figure Legend of 2a. Total cellular proteins (40 µg) were resolved by SDS-PAGE, transferred to nitrocellulose membranes, and detected with specific antibodies, as described in methods. A representative immunoblot of three separate experiments is shown. Cells were treated as described in the Figure Legend of 2b. Control (Con) values were obtained in the absence of LPS or tested samples. Total RNA was prepared for the RT-PCR analysis of COX-2 gene expression. COX-2-specific sequences (721 bp) was detected by agarose gel electrophoresis, as described in methods. PCR of β -actin was performed to verify
that the initial cDNA contents of the samples were similar.
The experiment was repeated three times and similar results were obtained. The Western blot results and the RT-PCR results are shown by relative ratio graphs. The values are the mean ± S.D. of three independent experiments.
#p <0.05 vs. the control group; * p <0.05, ** p <0.01 vs. the LPS-treated group;
the significances of the difference between the treated groups
was evaluated using the Student’s t -test.
luciferase 활성을 측정하였다 .
통계학적 분석 − 실험치의 값은 mean±S.D. 로 나타냈으
며 분석은 Student’s t -test 로 그 유의성을 나타내었다 .
결 과
SAL 의 Nitrite 생성 저해 − iNOS 에 의한 NO 생성은 LPS 에 의해 활성화된 RAW 264.7 세포의 배양액 중에 존
재하는 nitrite (NO
2−) 의 양을 Griess 시약을 사용하여 측정
함으로써 조사하였으며 , 양성 대조군으로는 L-arginine 과의
기질경쟁에 의하여 iNOS 를 저해하는 것으로 알려진 L-NIL (10 µM) 을 사용하였다 . SAL 은 농도 의존적으로 NO 생성
을 저해하였으며 100 µ M 에서 NO 생성을 38.0% 저해하였
다 (Fig. 2a). 이러한 NO 생성 저해가 세포독성에 기인하는
것을 확인하기 위하여 MTT 법을 이용하여 세포독성을 관찰
하였으나 최대 200 µ M 에서 RAW 264.7 세포에 독성을 나
타내지 않는 것을 확인하였다 (data not shown).
SAL 의 iNOS 단백질 생성과 mRNA 발현 저해 − SAL
에 의한 NO 생성 저해기전을 확인하기 위해 Western blot
과 RT-PCR 로 iNOS 의 단백질 생성과 mRNA 발현을 조사
하였다 . LPS 에 의해 증가된 iNOS 단백질 생성과 mRNA
발현이 SAL 을 처리하였을 때 농도 의존적으로 감소하였으
며 50 µM 과 100 µM 에서 유의성있게 감소되었다 (Fig. 2b).
β -actin 의 band density 비율에 따라 iNOS 단백질 생성과 mRNA 발현 정도를 보정하였을 때 SAL 은 100 µM 농도에
서 iNOS 단백질 생성을 29.0%, mRNA 발현을 47.9% 저
해함을 확인하였다 . 이를 통해 SAL 은 iNOS 의 생성과 발현
을 저해함으로써 NO 의 생성을 저해함을 알 수 있었다 .
SAL 의 PGE
2형성 및 COX-2 단백질 생성과 mRNA
발현 저해 효과 − RAW 264.7 세포에서 염증인자인 PGE
2의 SAL 에 의한 억제효과를 ELISA kit 를 이용하여 정량하
였다 . LPS 에 의한 PGE
2생성이 SAL 을 처리하였을 때 농
도 의존적으로 유의성있게 감소되었으며 (Fig. 3a), 100 µM
농도의 SAL 에 의해 PGE
2형성이 38.1% 저해되었다 . 선택
적인 COX-2 저해제로 알려진 NS398 (5 µM) 을 양성대조군
으로 사용하여 PGE
2의 형성이 뚜렷하게 저해되는 것을 확 인하였다 . SAL 에 의한 PGE
2형성 저해기전을 확인하기 위 해 Western blot 과 RT-PCR 로 COX-2 단백질 생성과 mRNA 발현을 조사하였다 . LPS 에 의해 증가된 COX-2 단
백질 생성과 mRNA 발현이 SAL 을 처리하였을 때 농도 의
존적으로 유의성있게 감소되었다 (Fig. 3b). β -actin 의 band density 비율에 따라 iNOS 단백질 생성과 mRNA 발현 정
도를 보정하였을 때 SAL 은 100 µM 농도에서 COX-2 단백
질 생성을 25.9%, mRNA 발현을 40.6% 저해함을 확인하
였다 . 이를 통해 SAL 은 iNOS 의 mRNA 발현을 억제하여
단백질 생성을 저해함으로써 NO 의 생성을 저해함을 알 수
있었다 .
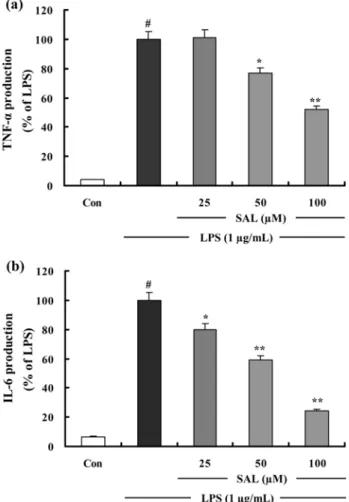
SAL 의 TNF- α 와 IL-6 의 형성 및 mRNA 발현 저해
효과 − SAL 이 RAW 264.7 세포에서 염증유도 사이토카인
인 TNF- α와 IL-6 을 억제하는지 알아보기 위해 ELISA kit
와 RT-PCR 을 이용하여 TNF- α와 IL-6 의 형성 및 mRNA
발현을 조사하였다 . LPS 처리에 의해 생성된 TNF- α는 고
농도 (100 µM) 의 SAL 에 의해 48.1% 저해되었다 (Fig. 4a).
LPS 에 의한 IL-6 형성은 SAL 에 의해 농도 의존적으로 감
소되었으며 , 100 µM 농도의 SAL 에서 75.6% 저해됨을 확인
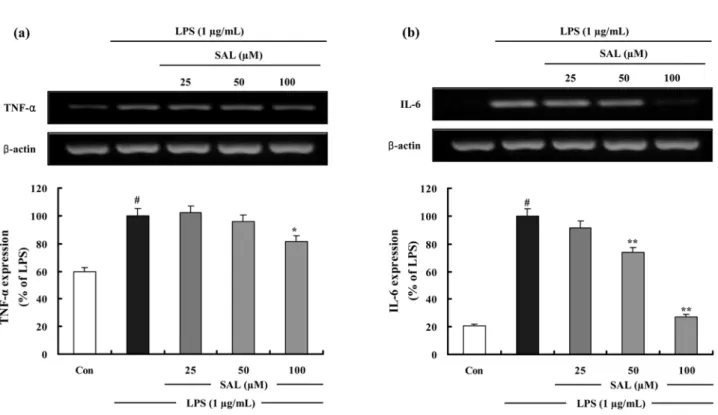
하였다 (Fig. 4b). 또한 SAL 은 LPS 에 의한 TNF- α와 IL-6 mRNA 의 발현을 유의성있게 저해하며 TNF- α와 IL-6 의 형
성 억제효과와 상관성이 있음을 확인하였다 (Fig. 5a, Fig. 5b).
Fig. 4. The effects of SAL on LPS-induced TNF- α and IL-6 production in RAW 264.7 cells. (a) Effect of the SAL on TNF- α production by LPS-induced RAW 264.7 macrophages for 24 h. (b) Effect of the SAL on IL-6 production by LPS- induced RAW 264.7 macrophages for 24 h. TNF- α and IL-6 production results are shown by relative ratio graphs. The values are the mean±S.D. of three independent experiments.
#
p <0.05 vs. the control group; * p <0.05, ** p <0.01 vs. the LPS-
treated group; the significances of the difference between the
treated groups was evaluated using the Student’s t -test.
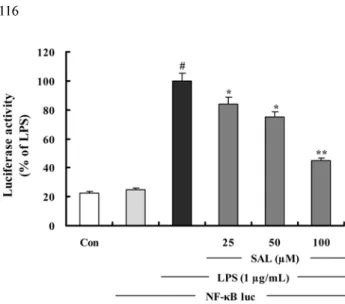
SAL 의 NF- κ B 활성 저해 효과 – LPS 에 의해 유도되는 iNOS, COX-2, TNF- α , 그리고 IL-6 등의 유전자 프로모터
에 존재하는 NF- κ B 요소 (element) 가 이들의 유전자 발현
에 중요한 역할을 한다는 보고를 통해 ,
22)SAL 이 LPS 에 의
한 NF- κ B 의 활성화를 억제하는지 알아보기 위해 luciferase assay 를 수행하였다 . RAW 264.7 세포에 일시적으로 pNF-
κ B-luc plasmid 를 형질감염 시키고 SAL 을 처리한 군과 처
리하지 않은 대조군에 LPS (1 µg/mL) 로 자극을 가한다 . SAL 이 LPS 에 의해 유도된 NF- κ B 의존적인 luciferase 효
소의 발현을 농도 의존적으로 유의성있게 감소시키는 것을 확인하였다 (Fig.6).
고 찰
본 연구진은 여러 천연물의 성분 중에서 LPS 에 의해 활
성화된 RAW 264.7 세포로부터 유도되는 NO 생성을 저해
하는 물질을 검색하는 과정에서 , 산겨릅나무 ( Acer tegmentosum Maxim) 의 목부에서 분리한 salidroside (SAL)
이 RAW 264.7 세포에서 항염증 효과를 나타냄을 발견하였
다 . SAL 은 이전의 연구에서 COX 와 5-lipoxygenase 를 억제
하여 PGE
2와 thromboxane B
2를 감소시키는 antieicosanoid
작용이 보고되어 있으나
23)염증인자 억제효과와 그 기전은 보고된 바 없다 . 따라서 본 연구에서 SAL 이 어떤 기전을
통해 항염증 효과를 나타내는지 연구하였다 .
대식세포는 LPS 와 같은 염증유발물질에 의해 활성화되어 toll-like receptor-4 (TLR-4) 의 발현을 자극한다 .
24)TLR-4 는 LPS/CD-14 복합체에 대한 막투과성 수용체로서 NF- κ B 를
통해 염증유도의 단계적 과정 (cascade) 을 활성화한다 .
25-27)이러한 기전을 통해 NO, PGs, 염증유도 사이토카인 등의
다양한 염증매개물질이 유도된다 .
28)본 실험에서는 먼저 SAL 이 RAW 264.7 세포에서 LPS 에
의해 유도된 NO 생성을 농도 의존적으로 뚜렷하게 감소시
키는 것을 확인하였으며 위의 사실에 기초하여 NO 생성저
해의 기전을 알아보기 위하여 Western blot 과 RT-PCR 로 iNOS 의 발현을 분석한 결과 SAL 에 의한 iNOS 의 발현억제
가 NO 생성억제와 유사한 경향을 나타냄으로써 NO 생성
억제는 iNOS 의 발현저해를 경유한 것임을 알 수 있었다 . 또
한 다수 항염증 약물들의 작용기전이 되는 COX-2 의 생성
Fig. 5. The effects of SAL on LPS-induced TNF- α and IL-6 mRNA expressions in RAW 264.7 cells. Cells were treated as
described in the Figure Legend of 2b. Control (Con) values were obtained in the absence of LPS or tested samples. (a) Total RNA
was prepared for the RT-PCR analysis of TNF- α gene expression. TNF- α -specific sequences (351 bp) was detected by agarose gel
electrophoresis, as described in methods. (b) Total RNA was prepared for the RT-PCR analysis of IL-6 gene expression. IL-6-
specific sequences (142 bp) was detected by agarose gel electrophoresis, as described in methods. PCR of β -actin was performed to
verify that the initial cDNA contents of the samples were similar. TNF- α and IL-6 RT-PCR results are shown by relative ratio
graphs. The values are the mean±S.D. of three independent experiments.
#p <0.05 vs. the control group; * p <0.05, ** p <0.01 vs. the
LPS-treated group; the significances of the difference between the treated groups was evaluated using the Student’s t -test.
억제와 그에 따른 PGE
2형성억제가 SAL 에 의해서 나타나
는지 조사하였다 . SAL 은 LPS 에 의해 형성되는 PGE
2를 유 의성있게 감소시키며 이는 COX-2 단백질 생성과 mRNA 의
발현저해에 의한 것임을 확인할 수 있었다 .
염증매개물질인 TNF- α와 IL-6 는 in vivo 및 in vitro 모 두에서 염증반응을 조절하는 물질로 알려져 있다 . 이러한
사이토카인들은 상호작용이 있는 것으로 알려져 있으며 LPS
와 같은 염증자극물질에 의해 형성이 유도된다 .
29)LPS 에 의
해 유도된 TNF- α와 IL-6 의 형성을 SAL 이 농도 의존적으
로 뚜렷이 저해하였으며 RT-PCR 로 분석한 결과 TNF- α와
IL-6 의 mRNA 발현과 매우 유사한 경향을 나타냄으로써
TNF- α와 IL-6 의 형성억제는 그것의 전사단계를 저해함에
의한 것임을 알 수 있었다 .
이러한 COX-2, iNOS 그리고 염증유도 사이토카인들의
발현에 있어서 NF- κ B 가 promoter 에 작용하여 중요한 조절
인자로 작용한다 .
30)본 실험에서는 luciferase assay 를 이용
하여 NF- κ B 전사활성을 측정함으로써 SAL 이 NF- κ B 신호
전달의 조절에 의해 염증전구물질의 형성을 억제함을 확인 하였다 .
이상의 결과들을 요약하면 SAL 은 RAW 264.7 세포에서 NF- κ B 의 전사작용을 억제함으로써 LPS 에 의해 유도되는 iNOS, COX-2, TNF- α 그리고 IL-6 의 생성과 유전자 발현
을 효과적으로 저해한다 . 이러한 결과들은 salidroside 을 이
용하여 염증성 질환을 예방하거나 치료할 수 있는 약물로 서 개발될 수 있는 가능성을 제시하고 있다 .
사 사
이 논문은 경희대학교 대학원의 2007 학년도 2 차 우수연
구논문 장학금으로 지원을 받아 수행된 연구임 . 인용문헌
1. Brown, K. L., Cosseau, C., Gardy, J. L. and Hancock, R. E.
(2007) Complexities of targeting innate immunity to treat infection. Trends. Immunol. 28 : 260-266.
2. Posadas, I., Terencio, M. C., Guillén, I., Ferrándiz, M. L., Coloma, J., Payá, M., Alcaraz, M. J. (2000) Co-regulation between cyclo-oxygenase-2 and inducible nitric oxide syn- thase expression in the time-course of murine inflammation.
Naunyn Schmiedebergs Arch. Pharmacol. 361 : 98-106.
3. Moncada, S., Palmer, R. M., Higgs, E. A. (1991) Nitric oxide: physiology, pathophysiology, and pharmacology.
Pharmacol. Rev. 43 : 109-142.
4. Yun, H. Y., Dawson, V. L. and Dawson, T. M. (1996) Neu- robiology of nitric oxide. Crit. Rev. Neurobiol. 10 : 291-316.
5. Stuehr, H. H. J., Kwon, N. S., Weise, M. and Nathan, C.
(1991) Purification of the cytokine-induced macrophage nitric oxide synthase: an FAD- and FMN- containing fla- voprotein. Proc. Natl. Sci. USA. 88 : 7773-7777.
6. McCartney-Francis, N., Allen, J. B., Mizel, D. E., Albina, J.
E., Xie, Q. W., Nathan, C. F. and Wahl, S. M. (1993) Sup- pression of arthritis by an inhibitor of nitic oxide synthase. J.
Exp. Med. 178 : 749-754.
7. Weisz, A., Cicatiello, I. and Esumi, H. (1996) Regulation of the mouse inducible-type nitric oxide synthase gene pro- moter by interferon-gamma, bacterical lipopolysaccharide and NG-monomethyl-L-arginene. Biochem. J. 316 : 209-215.
8. Vane, J. R., Botting, R. M. (1998) Mechanism of action of nonsteroidal anti-inflammatory drugs. Am. J. Med. 104 : 2S-8S.
9. Vane, J. R., Bakhle, Y. S., Botting, R. M. (1998) Cycloox- ygenases 1 and 2. Annu. Rev. Pharmacol. Toxicol. 38 : 97- 10. Needleman, P. and Isakson, P. C. (1997) The discovery and 120.
function of COX-2. J. Rheumatol. Suppl. 49 : 6-8.
11. Seybold, V. S., Jia, Y. P. and Abrahams, L. G. (2003) Cyclo- oxygenase-2 contributes to central sensitization in rats with peripheral inflammation. Pain 105 : 47-55.
12. Bishop-Bailey, D., Calatayud, S., Warner, T. D., Hla, T. and Mitchell J. A. (2002) Prostaglandins and the regulation of tumor growth. J. Environ. Pathol. Toxicol. Oncol. 21 : 93- 13. Willeaume, V., Kruys, V., Mijatovic, T. and Huez, G. (1995) 101.
Tumor necrosis factor-alpha production induced by viruses
Fig. 6. The inhibition of NF- κ B-DNA binding by SAL. Cells were transiently cotransfected with pNF- κ B-luc reporter and then left untreated (Con) or were pretreated with different concentrations (25, 50, 100µM) of SAL. LPS (1µg/mL) was then added and the cells were further incubated for 3 h. The cells were then harvested and luciferase activities were determined using a Promega luciferase assay system and a luminometer. The values are the mean±S.D. of three independent experiments.
#