대한소화기학회지 2002;39:324 - 334
서 론
2)위점막을 둘러싸고 있는 상피세포는 Helicobacter pylori 와 인체가 제일 처음 접촉하는 부위이다. 즉, 위상피세포는
접수: 2002년 1월 7일, 승인: 2002년 3월 13일 연락처: 정현채, 110-744, 서울시 종로구 연건동 28번지
서울대학교 의과대학 내과학교실 Tel: (02) 740-8120, Fax: (02) 743-6701 E-mail: [email protected]
※본 연구는 한국야쿠르트(주)의 연구비 지원으로 이루어졌음.
H. pylori와 반응하여 interleukin (IL)-8과 같은 chemokine과 cyclooxygenase-2 (COX-2)와 같은 일련의 친염증성 매개체 를 활성화시키고, 최종적으로 apoptosis 과정을 밟게 된 다.1-3 또한 사람 호중구와 림프구에서도 H. pylori 감염에 의해 친염증성 cytokine이 발현될 수 있음이 증명된 바 있 다.4-7 이와 같은 사실은 침윤된 염증세포에서 생성되는 신 호(signal)가 H. pylori 감염에 의한 위상피세포의 염증반응 에 영향을 미칠 수 있음을 시사해 준다.
산화질소(nitric oxide, NO)는 NO 합성효소(NO synthase, NOS)에 의해 L-arginine이 L-citrulline으로 변환되는 과정
Helicobacter pylori 감염에 의한 인체 위상피세포로부터의 Inducible Nitric Oxide Synthase 발현 및 Nitric Oxide 생성
한양대학교 의과대학 미생물학교실 및 의과학연구소, 서울대학교 의과대학 내과학교실 및 간연구소*
김정목·김주성*·정현채*·송인성*·김정룡*
Expression of Inducible Nitric Oxide Synthase and Nitric Oxide Production in Human Gastric Epithelial Cells Infected with Helicobacter pylori
Jung Mogg Kim, M.D., Joo Sung Kim, M.D.*, Hyun Chae Jung, M.D.*, In Sung Song, M.D.*, and Chung Yong Kim, M.D.*
Department of Microbiology and Institute of Biomedical Science, Hanyang University College of Medicine, Seoul;
Department of Internal Medicine and Liver Research Institute, Seoul National University College of Medicine*, Seoul, Korea
Background/Aims: Nitric oxide (NO) is known to be an important regulator of inflammatory response. Our aim was to study the role of inducible nitric oxide synthase (iNOS) expression and NO production in Helicobacter pylori (H. pylori)-infected gastric epithelial cells. Methods: Human gastric epithelial Hs746T cells were infected with H. pylori and iNOS mRNA expression was assessed by quantitative RT-PCR. NO production was assayed by determining nitrite/nitrate levels in culture supernatants. To determine the polarity of NO secretion, Caco-2 cells were cultured in transwell chambers and NO production was measured. Results: iNOS mRNA and NO production were significantly increased in the cells infected with H. pylori. The specific MAP kinase inhibitors decreased H.
pylori-induced iNOS and NO up-regulation. After polarized Caco-2 epithelial cells were infected with H. pylori, NO was released predominantly into the apical compartment, and IL-8 was released predominantly into basolateral compartment. The addition of IFN- to H. pylori-infected Caco-2 cells showed a synergistically apical and basolateral NO release. Conclusions: These results suggest that apical NO production in H. pylori-infected gastric epithelial cells may influence the bacteria and basolateral production of NO may play a role in the tissue inflammation. (Korean J Gastroenterol 2002;39:324-334)
Key Words: Gastric epithelial cells, Helicobacter pylori, Inducible nitric oxide synthase
김정목 외 4인. H. pylori에 의한 iNOS 발현
에서 생성된다. 이 과정을 매개하는 NOS는 3가지 종류 가 알려져 있다.8,9 이중에서 유도성 NOS (inducible NOS, iNOS)는 지질다당체(lipopolysaccharide, LPS)로 자극한 각 종 세포에서 발현될 뿐만 아니라8,9 침습성 세균에 감염된 상피세포10 또는 IL-1과 같은 cytokine의 자극에 의해서도 발현이 증가한다.11,12 또한 신경 NOS (neuronal NOS, nNOS) 와 내피세포 NOS (endothelial NOS, eNOS)도 알려져 있 다.8,9 비록 H. pylori에 감염된 위점막에서 iNOS의 발현이 증가한다는 사실이 보고되고 있지만,13-15아직까지 H. pylori 가 위상피세포에서 iNOS를 어떻게 유도하는지에 대해서는 확실하게 규명되어 있지 않다. 뿐만 아니라 iNOS 발현과 NO 생성이라는 관점에서 위상피세포와 염증세포 사이의 상호관계에 대해서도 확실하게 알려진 바가 없다.
위점막에 존재하는 림프구는 H. pylori 감염에 의해 IL-1 과 interferon (IFN)- 와 같은 친염증성 cytokine을 발현한 다.6,7 또한 위점막에 존재하는 T 세포는 H. pylori 감염과 관 계없이 많은 양의 IFN- 를 생성해낼 수 있다고 알려져 있 다.16 이와 같은 IL-1과 IFN- 는 몇몇 종류의 상피세포에서 iNOS 발현을 증가시킬 수 있다.11,12,17위의 보고들을 토대로 저자들은 H. pylori 감염에 의해 유도되는 iNOS 발현과 NO 생성이 위점막에 존재하는 염증세포로부터 생성되는 면역매개체(immune mediator)와 연관되어 있을 것이라는 가 설을 설정하였다. 이와 같은 가설을 증명하기 위하여 본 연 구에서는 H. pylori 감염에 의해 위상피세포로부터 iNOS의 발현과 NO 생성이 증가할 수 있는지의 여부를 확인하였다.
또한 염증세포로부터 생성되는 면역매개체가 위상피세포에 의한 NO 생성을 조절할 수 있는지를 관찰하였다.
대상 및 방법
1. H. pylori 분리 및 배양
본 연구에 사용된 H. pylori 균주는 기존의 방법을 이용 하여 분리 배양하였다.18 계대배양 횟수가 10회 미만의 균 을 이용하기 위하여 H. pylori 균주들은 15% glycerol이 포 함된 Brucella broth (Difco Laboratories, Detroit, MI, USA) 에 넣어 -70℃에 보관하였고, 실험 직전 녹인 뒤 증식시켜 사용하였다. H. pylori 균주의 독성인자인 cagA 및 공포성 세포독소 (vacuolating cytotoxin)검사는 기존의 방법대로 시 행하였다.18
H. pylori 수용성 단백질(water-soluble protein, HPWP)은 기 존의 방법을 이용하여 추출하였다.5,19 즉, 세균을 멸균된 증 류수에 모은 후(1×1010/mL) 실온에 10분간 방치시켰다.
그 뒤, vortex를 이용하여 20초간 혼합한 뒤, 12,000 rpm (17,400 g)으로 15분간 원심하였다. 이렇게 얻어진 상청액을
-70℃에 보관하였다. 실험 직전 18,000 rpm (39,100 g)으로 20분간 원심한 뒤 0.2 µm filter로 여과한 것을 사용하였다.
총 단백질 양은 Bradford 검사법(Bio-Rad, Hercules, CA, USA)으로 측정하였다. 또한 기존의 방법대로5,19 urease 활성 및 내독소(endotoxin)의 양은 각각 결합효소법(coupled enzyme assay)과 Limulus amoebocyte lysate assay (Associates of Cape Cod, Inc., Falmouth, MA, USA)로 측정하였다.
대조균으로는 Campylobacter jejuni, Campylobacter fetus subsp. fetus, 비병원성 Escherichia coli DH5α(Promega, Madison, WI, USA)를 이용하였다.
2. 위상피세포주 배양 및 감염
실험에 이용한 기본 인체 위상피세포주로는 Hs746T 세 포(ATCC HTB 135)였다. 이 세포는 Dulbecco's modified Eagle's medium (DMEM, pH 7.4, Sigma, St. Louis, MO, USA) 에 10% 우태아혈청(fetal bovine serum, FBS, Hyclone Laboratories, Logan, UT., USA)과 항생제 (100 unit/mL의 penicillin, 100 µg/mL의 streptomycin)를 사용하여 37℃ 5%
CO2 배양기에서 배양하였다.
세균 감염은 항생제가 제거된 상태에서 세포를 6-well tissue culture plate에 24-48시간 동안 배양한 다음 균수:위 상피세포의 비율이 250:1이 되게 균을 첨가하였다. 균수 측정은 McFarland scale과 hemocytometer를 이용하였다.18
경우에 따라서는 H. pylori 감염과 함께 NOS 억제제인 nitro-L-arginine methyl ester (L-NAME, ICN Biomedical, Aurora, OH, USA)를 첨가하여 실험에 이용하였다. 또한 p38 mitogen activated protein (MAP) kinase 특이 억제제인 SB203580 (10 mM, Calbiochem, La Jolla, CA, USA) 또는 MAP kinase 억제제 PD98059 (25 mM, Calbiochem)를 H. pylori 감염 30분 전에 미리 전처치하여 실험에 이용하였다.20,21
극성(polarity) 실험을 위해 사용한 Caco-2 (5×105) 세포는 20% FBS가 포함된 RPMI-1640 배지를 이용하여 transwell chamber (Costar, Cambrige, MA, USA)의 collagen-coated microporous supports (0.4 µm pore size; 4.7 cm2 growth area) 상단부에 배양하였다. 그 뒤, 매 2일마다 배지를 교환 하고, 15일 동안 배양하여 세포가 완전히 분화되도록 하였 다.22이 때 apical compartment와 basal compartment의 배지 양을 각각 1.5 mL과 2.6 mL로 조정하였다. 15일간 배양 후 장상피세포의 분화와 세포간 tight junction이 완전히 이루 어진 상태에서 H. pylori를 감염시키고, 일정 시간 배양한 다음, apical compartment와 basal compartment로부터 배양 액을 회수하였다. Tight junction의 형성은 Millicell electrical resistance system (Millipore, Bedford, MA, USA)을 이용하 여 monolayer를 통한 전기저항(transepithelial resistance)의 측정으로 평가하였다.
325
The Korean Journal of Gastroenterology: Vol. 39, No. 5, 2002
3. 사람 말초혈액 림프구와 호중구 분리
건강한 지원자로부터 헤파린 처리 혈액을 얻은 후, Ficoll- Hypaque (Amersham Pharmacia Biotech, Uppsala, Sweden) density gradient centrifugation 방법을 이용하여 사람 말초 혈액 림프구(human peripheral blood lymphocytes, PBMC) 를 분리하였다. 분리한 PBMC는 Hanks' balanced salt solution (HBSS)으로 3회 세척한 뒤 10% FBS가 포함된 RPMI-1640 배지(Sigma)에 0.5×106 cells/mL로 재부유시 켰다.
사람 호중구는 기존의 방법대로4,19 건강한 지원자로부 터 얻은 혈액을 dextran sedimentation 및 Ficoll-Hypaque (Histopaque 1077, Sigma) density gradient separation법으로 분 리하였다. 세포의 생존도는 trypan blue exclusion검사로 시 행하였으며, 95% 이상의 생존을 나타내는 호중구를 실험 에 이용하였다. 한편 호중구의 순도는 cytospin을 이용한 Wright-Giemsa염색법으로 확인하였다. 이 때 호중구는 97%
이상이었고, 오염된 림프구는 0.2% 미만이었다. 한편 단핵 구 오염은 fluorescein isothiocyanate (FITC)-conjugated anti- CD14 항체로 염색한 뒤 flow cytometry (Becton Dickinson, San Jose, CA, USA)를 이용하여 측정하였다. 그 결과 단핵 구 오염도는 10,000개의 세포당 0.02% 이하였다. 이와 같이 분리한 호중구를 HBSS로 2회 세척한 뒤, 10 mM HEPES (Sigma)와 10% FBS가 포함된 RPMI-1640 배지에 5×106/mL 농도로 부유시켰다.
4. RNA 추출 및 RT-PCR
위상피세포주에 H. pylori를 일정 시간 동안 감염시킨 뒤, guanidinium thiocyanate-phenol-chloroform법으로 세포 RNA
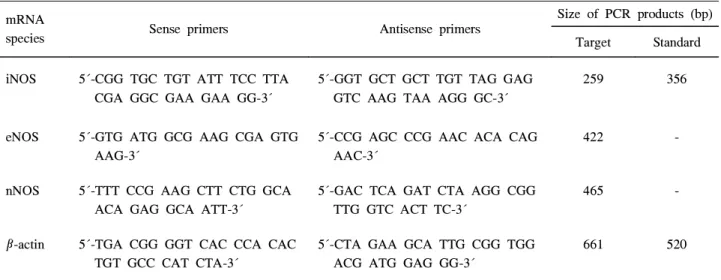
Table 1. Sequences of Oligonucleotide Primer for PCR Analysis of Human iNOS, eNOS, and nNOS mRNA
species Sense primers Antisense primers
Size of PCR products (bp) Target Standard iNOS 5´-CGG TGC TGT ATT TCC TTA
CGA GGC GAA GAA GG-3´
5´-GGT GCT GCT TGT TAG GAG GTC AAG TAA AGG GC-3´
259 356
eNOS 5´-GTG ATG GCG AAG CGA GTG AAG-3´
5´-CCG AGC CCG AAC ACA CAG AAC-3´
422 -
nNOS 5´-TTT CCG AAG CTT CTG GCA ACA GAG GCA ATT-3´
5´-GAC TCA GAT CTA AGG CGG TTG GTC ACT TC-3´
465 -
β-actin 5´-TGA CGG GGT CAC CCA CAC TGT GCC CAT CTA-3´
5´-CTA GAA GCA TTG CGG TGG ACG ATG GAG GG-3´
661 520
를 분리하였다. iNOS 및 β-actin에 대한 정량적 RT-PCR은 표준 합성 RNA를 이용하여 기존의 방법대로 시행하였 다.1,10 즉, 일정 수의 표준합성 RNA와 추출한 세포 RNA를 함께 역전사를 시행한 후, 얻어진 cDNA를 특이한 primer를 이용하여 PCR을 시행하였다. 실험에 사용한 iNOS, eNOS 및 nNOS primer의 염기서열은 Table 1과 같다.10,23,24 이와 같은 primer를 이용하여 얻은 iNOS의 PCR 산물의 크기는 표준합성 RNA의 경우 356 bp였고, 표적 RNA는 259 bp였 다. PCR 증폭은 thermal cycler (GenAmp PCR System 9600, Perkin Elmer Cetus, Norwalk, CT, USA)를 이용하여 iNOS와 eNOS의 경우 94°C에서 45초, 60°C에서 45초 그리 고 72°C에서 2분간 시행하였다. nNOS의 경우에는 95°C에 서 35초, 60°C에서 2분간 시행하였다. 이들의 시행 횟수는 총 35회였다. β-actin에 대한 정량적 RT-PCR은 기존의 방 법과 동일하게 시행하였다.1 한편 iNOS에 대한 양성 대조 군으로는 PBMC, eNOS에 대한 양성 대조군으로는 사람 제 대혈관 내피세포, nNOS에 대한 양성 대조군으로는 사람 astrocytoma 세포로부터 분리한 RNA를 이용하였다.10 모든 실험은 3회 이상 반복 시행하였다.
PCR 생성물은 2% NuSieve agarose gel (FMC Bioprod- ucts, Rockland, ME, USA)을 이용한 전기영동으로 분리하고 폴라로이드 필름에 영상을 기록하였다. 이 영상을 imaging densitometer (GS-670, BioRad, Hurcules, CA, USA)로 처리 하여 각 PCR 밴드의 peak에 해당하는 면적을 구하였다.
그 뒤 세포 RNA로부터 얻은 PCR 면적과 표준 RNA로부터 얻은 PCR 면적비를 이중 로그 스케일을 이용하여 표준 RNA 분자수와 동일한 등전점(equimolar point)을 구하여 세포 RNA 내에 존재하는 iNOS mRNA 분자수를 계산하 였다.
326
Kim, et al. H. pylori-induced iNOS expression
5. NO 및 IL-8 단백질 측정
안정화된 NO 최종 산물인 nitrite와 nitrate의 양을 측정 하기 위하여 먼저 nitrate를 nitrite로 환원시켰다. 즉, 배양 상청액 50 µL를 39 µL의 3[N-morpholino]propanesulfonic acid (MOPS, 50 mM, pH 7.0, Sigma), 10 µL의 nicotinamide adenine dinucleotide phosphate (NADH, 2 mM, Sigma), 1 µL의 nitrate reductase (5 U/mL, Oxford Biomedical, Oxford, MI, USA)와 혼합한 뒤, 실온에서 60분 동안 반응 시켰다. 그 뒤, nitrite 농도는 Griess 반응으로 측정하였다.
즉, 배양상청액 100 µL를 동일 용량의 Griess 반응액 (0.5% sulfanilamide, 2.5% H3PO4, 0.05% naphthylethylene diamine)과 혼합하였다. 그 뒤 실온에서 10분간 반응시킨 다음, 550 nm에서 흡광도를 측정하여 sodium nitrite로 얻 은 표준곡선과 비교하여 nitrite의 농도를 구하였다.10
IL-8 단백질은 배양상청액을 대상으로 효소면역측정법 (ELISA)으로 측정하였다. ELISA 방법은 R&D System 사 (Quantikine, Minneapolis, MN, USA)의 kit를 이용하였고, 최소 측정 단위는 20 pg/mL이었다.
6. iNOS 단백질 확인을 위한 Western blot
6-well plate에 배양한 Hs746T 세포를 H. pylori에 감염 시킨 24시간 뒤, 얼음 속에서 냉각시킨 phosphate buffer saline (PBS, pH 7.4)으로 3회 세척하였다. 그 뒤, well당 0.5 mL의 lysis buffer (150 mM NaCl, 20 mM Tris, pH 7.5, 0.1% Triton X-100, 1 mM PMSF, 10 µg/mL aprotinin) 를 넣어 세포를 파쇄하였다. 이 세포액을 12,000 g (4℃) 에서 20분간 원심하여 상청액을 회수하였다. 단백질 농도 측정은 Bradford법을 이용하였다. Western blot에 사용된 단백질 양은 well당 15 µg이었다. SDS-PAGE 후 단백질을 nitrocellulose membrane (0.1 µm pore size, Hybond ECL, Amersham, Buckinghamshire, England)으로 전기 이동시킨 다음, 마우스 항-사람 iNOS 또는 마우스 항-사람 eNOS 단세포군 항체 (Transduction Laboratories, Lexington, KY, USA)와 반응시켰다. 그 뒤 2차항체(per- oxidase가 결합된 항-마우스 IgG, Transduction Labo- ratories)와 enhanced chemiluminescence detection kit (ECL, Amersham)를 이용하여 각각의 NOS 단백질 발현을 검 사하였다.
7. 통계 처리
자료 분석은 SAS System software release 6.12 (SAS Institute Inc., Cary, NC, USA)를 이용하여 Wilcoxon rank sum test를 이용하여 유의성을 검증하였으며, p 값이 0.05 이하인 경우를 유의한 값으로 인정하였다.
결 과
1. H. pylori 감염에 의한 위상피세포에서의 iNOS 발현 및 NO 생성
사람 위상피세포주인 Hs746T 세포에 H. pylori를 감염시 킨 뒤 RT-PCR로 3종류의 NOS mRNA 발현을 검사하였다.
그 결과 H. pylori 감염에 의해 iNOS mRNA 발현이 증가 함을 확인할 수 있었다. 그러나 eNOS mRNA와 nNOS mRNA는 H. pylori 감염에 의해 별다른 차이를 나타내지 못했다(Fig. 1). 발현된 mRNA를 정량한 결과, H. pylori 감염 12시간에 최대의 iNOS mRNA 발현 분자수를 나타내었다.
반면 β-actin mRNA는 전 감염 시간에 걸쳐 유의한 변화를 나타내지 못했다(Fig. 2A). 또한 배양상청액을 대상으로 측 정한 NO 농도는 iNOS mRNA의 증가와 일치하고 있었다 (Fig. 2B). 한편 이와 같은 실험 system에서 IL-8의 생성량 은 감염군 5,836±456 pg/mL, 대조군 435±67 pg/mL (mean±SEM, n=7)을 나타내어 기존 실험 보고와 동일한 결과를 나타냄을 알 수 있었다.25 한편 H. pylori 배양액을 Hs746T 세포에 24시간 동안 처리했을 때는 NO의 유의한 증가를 관찰할 수 없었다(대조군, 10.3±2.5 µM; H. pylori 감염군, 35.7±4.9 µM; H. pylori 배양상청액을 처리한 군,
Fig. 1. Several NOS mRNA expressions in gastric epithelial cells infected with H. pylori. Confluent Hs746T monolayers were grown in 6 well plates and were infected with cagA+cytotoxin+H. pylori for the indicated period. RNA was extracted from infected and uninfected cells. mRNA expression for iNOS, eNOS, nNOS, and β-actin was assessed by RT-PCR using specific primers.
C, uninfected control; H, H. pylori-infected cultures.
327
대한소화기학회지: 제39권 제5호, 2002
Fig. 2. Time course of increased iNOS mRNA and NO levels. (A) Quantification of iNOS and β-actin mRNA. Confluent Hs746T monolayers in 6-well plates were infected with cagA+cytotoxin+ H. pylori for the indicated period. To quantify the iNOS and β-actin transcripts, the total RNA was reverse-transcribed using an oligo (dT) primer and a synthetic internal RNA standard, and amplified by PCR. The data are expressed as mean±SD from five separate experiments. Black circle, H. pylori-infected;
Open circle, uninfected control. (B) NO levels. At the indicated times after H. pylori infection in the same culture and infection conditions as in (A), the culture supernatants were taken and the levels of NO were determined using a Griess reaction and nitrate reductase. The values shown are expressed as mean±SEM of seven separate experiments. Asterisks indicate values significantly different from those in the respective uninfected controls (p<0.05).
Black circle, H. pylori-infected; Open circle, uninfected control.
10.7±1.5 µM; mean±SEM, n=3). 이와 같은 결과는 NO의 증가를 위해서는 위상피세포와 H. pylori와의 상호 접촉이 필요하다는 점을 시사해 준다.
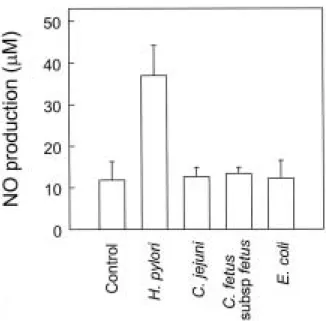
H. pylori 감염에 비하여 C. jejuni, C. fetus subsp fetus 및 비병원성 E. coli를 Hs746T 세포에 감염시킬 경우 NO 증가 는 관찰할 수 없었다(Fig. 3). 이와 같은 결과로 미루어 위 상피세포에서의 NO 생성은 H. pylori에 특이적인 현상이라 는 점을 시사해 준다.
Fig. 3. NO production by Hs746T cells after infection with bacteria. Confluent Hs746T monolayers in 6-well plates were infected with H. pylori or other bacteria for 24 hr. Then, culture supernatants were taken and the levels of NO were determined using a Griess reaction and nitrate reductase. The data shown are expressed as mean ±SEM of five separate experiments.
Fig. 4. Western blot for iNOS and eNOS. At eighteen hr of infection with cagA+cytotoxin+H. pylori, the Hs746T cell lysates were prepared, size-fractionated, and blotted onto a nitrocellulose membrane. iNOS and eNOS were detected with specific antibodies and enhanced chemiluminescence. Representative examples of X-ray films are shown. Major bands representing iNOS and eNOS are present at 130 kD and 140kD, respectively. C, uninfected control;
H, H. pylori-infected cultures.
328
김정목 외 4인. H. pylori에 의한 iNOS 발현
iNOS mRNA 증가가 세포질 내의 iNOS 단백질 발현으 로 이어지는지의 여부를 확인하기 위하여 Western blot을 시행하였다. 그 결과 H. pylori 감염에 의해 Hs746T 세포로 부터 iNOS 단백질의 발현이 증가하는 반면, eNOS 단백질 band는 별다른 변화를 관찰할 수 없었다(Fig. 4). 이 결과는 H. pylori 감염에 의해 증가하는 NO는 iNOS 발현 증가에 기인한다는 점을 제시해 준다. 이와 같은 사실을 재확인하 기 위하여 NOS 억제제인 L-NAME을 처리한 Hs746T 세포 에 H. pylori를 감염시킨 뒤, NO 농도를 측정하였다. 그 결 과 L-NAME 농도에 비례하여 NO 생성이 억제됨을 관찰할 수 있었다(Fig. 5).
Fig. 5. The effects of L-NAME on the NO production in H. pylori- infected Hs746T cells. Confluent monolayers of Hs746T cells in 6-well plates were infected with cagA+cytotoxin+ H. pylori in the presence of iNOS inhibitor L-NAME. The supernatants were harvested at 24 hr post-infection, and the levels of NO were determined using a Griess reaction and nitrate reductase. The data are expressed as the percentage of NO production compared with the cultures not treated with L-NAME. The values are expressed as mean±SEM of five separate experiments. The mean NO levels in the H. pylori-infected cultures not treated with L-NAME are 35.3±4.6 µM.
2.H. pylori 에 감염된 위상피세포에서의 iNOS 발현 및 NO 생성에 있어서의 MAP Kinase의 역할
H. pylori 감염은 위상피세포로부터 MAP kinase를 활성 화시키고, 이 활성은 IL-8 생성과 관련이 있다는 보고가 발표된 바 있다.26 따라서 본 연구에서 MAP kinase 활성 이 H. pylori 감염에 의해 유도되는 iNOS 발현과 NO 생 성에 연관성이 있는지를 관찰하였다. 이 실험을 위하여 p38 kinase 활성을 억제하는 MAP kinase 특이 억제제 SB203580과 MEK1 kinase 활성을 억제하여 ERK1/2 인산
화 과정(phosphorylation)을 차단하는 PD98059를 사용하였 다. 실험 결과 p38 억제제 SB203580 (10 µM)은 NO 생성을 약 43% 감소시켰으며 MEK1 억제제인 PD98059 (25 µM)는 NO 생성을 약 45% 감소시켰다(Fig. 6A). 한편 이 실험 system에서 p38 억제제와 MEK1 억제제는 H. pylori 감염 에 의해 유도되는 IL-8 생성을 각각 약 62%와 약 58% 감 소시켰다(Fig. 6B). 이와 같은 결과는 MAP kinase 활성 경 로가 H. pylori 감염에 의한 위상피세포로부터의 IL-8 생 성뿐만 아니라 iNOS 및 NO 발현에도 관여한다는 점을 제시해 준다.
Fig. 6. The effects of MAP kinase inhibition on the NO production in H. pylori-infected Hs746T gastric epithelial cells. Confluent monolayers of Hs746T cells in 6-well plates were treated with the specific MEK1 MAP kinase inhibitor, PD98059 (25 µM), and p38 MAP kinase inhibitor, SB203580 (10 µM), for 30 min before exposure to H. pylori. The supernatants were harvested at 24 hr post-infection with cagA+cytotoxin+ H. pylori. The levels of NO were determined using a Griess reaction and nitrate reductase (A), and IL-8 was assayed by ELISA (B). The data are expressed as mean±SEM of five separate experiments. Asterisks indicate values significantly different from those in H. pylori-infected cells without MAP kinase inhibitors (p<0.05).
329
The Korean Journal of Gastroenterology: Vol. 39, No. 5, 2002
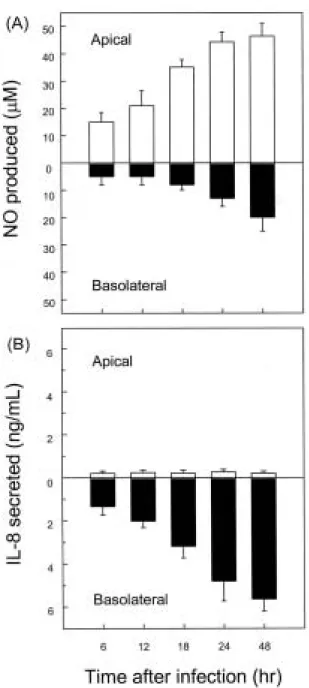
Fig. 7. Increased apical NO release after H. pylori infection of polarized Caco-2 monolayers. Polarized monolayers of the Caco-2 cells were grown in transwell chambers. cagA+cytotoxin+H. pylori was added to the apical compartment. Supernatants were removed from the apical and basolateral compartments at the indicated times. The levels of NO were determined using a Griess reaction and nitrate reductase (A), and IL-8 was assayed by ELISA (B).
Open bars and solid bars represent NO or IL-8 concentrations in the apical and basolateral compartments, respectively. The data are expressed as mean±SEM of five separate experiments.
3. H. pylori 감염에 의한 극성 Caco-2 세포로부터의 NO 분비 방향
생체 내에서 위장관 상피세포는 구조적으로뿐만 아니라 기능적으로도 극성을 나타낸다.27 이와 같은 극성 환경을
실험실적으로 조성하기 위하여 본 연구에서는 transwell chamber에 키운 Caco-2 세포주를 이용하였다.22즉, Caco-2 세포에 H. pylori를 감염시킨 6-48시간 후 NO 농도를 측정 해본 결과, H. pylori에 의해 상피세포로부터 생성되는 NO 는 주로 apical 방향으로 분비됨을 알 수 있었다(Fig. 7A).
반면 호중구 유인 chemokine인 IL-8은 주로 basolateral 방 향으로 분비되고 있었다(Fig. 7B).
4. 극성 Caco-2 세포로부터의 NO 분비 방향에 대한 IFN-γ의 영향
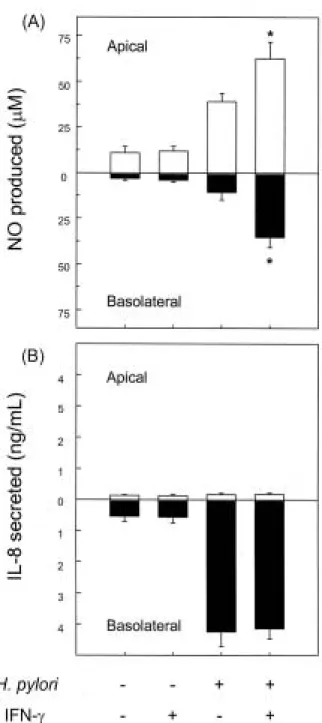
IFN- 는 상피세포의 NO 생성을 조절할 능력을 지닌다고 알려져 있다.11,12 이와 같은 IFN- 가 사람 면역세포에서 어 느 정도 발현되는지를 검사하였다. 실험 결과 H. pylori 수 용성 단백질(200 µg/mL)을 9시간 동안 처리한 사람 말초혈 액 단핵세포에서는 대조군에 비해 IFN- mRNA 발현이 약 6배 증가하였다(Fig. 8). 그러나 호중구에서는 IFN- mRNA 발현을 관찰할 수 없었다. 그런데 H. pylori 감염에 의해 유 도되는 위염에서 단핵세포 림프구는 주로 상피세포 표면 근처에 분포한다고 보고되고 있다.28 따라서 본 연구에서는 림프구에서 생성될 수 있는 IFN- 가 H. pylori 감염에 의해 나타나는 위상피세포에서의 NO 분비 방향에 어떤 영향을 미치는지 관찰하였다. 그 결과 basolateral compartment에 IFN- (50 ng/mL)를 첨가해 줄 경우 NO 생성이 apical 방 향뿐만 아니라 basolateral 방향으로도 증가함을 관찰할 수 있었다(Fig. 9A). 반면 IL-8 분비는 유의한 변화를 찾아볼 수 없었다(Fig. 9B).
Fig. 8. IFN-γ mRNA expression in human peripheral blood mononuclear cells treated with H. pylori water extract. Human peripheral blood mononuclear cells were stimulated with H. pylori water-soluble protein (HPWS, 200 µg/mL) for 9 hr. To quantify the IFN-γ transcripts, the total RNA was reverse-transcribed using an oligo (dT) primer and a synthetic internal RNA standard, and amplified by PCR. The data are expressed as mean±SD from five separate experiments.
330
Kim, et al. H. pylori-induced iNOS expression
Fig. 9. Synergistic increased apical and basolateral NO release by H. pylori-infected polarized Caco-2 monolayers after the addition of IFN-γ. The polarized monolayers of Caco-2 cells were grown in transwell chambers. Bacteria were added to the apical compartment and IFN-γ (50 ng/mL) was added to the basolateral compartment.
Supernatants were removed from apical and basolateral com- partments at 24 hr post-infection. The levels of NO were determined using a Griess reaction and nitrate reductase (A), and IL-8 was assayed by ELISA (B). Open bars and solid bars represent NO or IL-8 concentrations in the apical and basolateral compartments, respectively. The data are expressed as mean±SEM of five separate experiments. Asterisks indicate values significantly different from the H. pylori-infected cells without added IFN-γ (p<0.05).
고 찰
본 연구에서 H. pylori 감염에 의해 위상피세포로부터 iNOS가 발현되어 NO가 생성된다는 사실을 입증하였다. 또 한 H. pylori 감염에 의한 NO는 주로 apical 방향으로 분비 되고 있었다. 그렇지만 점막 내부의 염증세포로부터 분비 되는 IFN-γ에 의해 apical 방향뿐만 아니라 basolateral 방향 으로도 분비가 증가한다는 점은 매우 흥미로운 결과라고 할 수 있다. 이와 같은 결과들은 H. pylori에 감염된 위상피 세포와 염증세포 사이의 상호작용에 의해 유도되는 현상을 반영하는 것이라고 추정된다.
H. pylori 배양액을 위상피세포에 처리했을 경우에는 iNOS 발현과 NO 생성을 관찰할 수 없었다. 이 같은 현상 은 사람의 위조직을 면역화학염색한 결과 iNOS는 주로 H. pylori와 접촉하는 부위에서 관찰되었다는 보고13와 일치 하는 현상이라고 할 수 있다. 즉, iNOS 발현과 NO의 증가 를 위해서는 위상피세포와 H. pylori와의 상호 접촉이 필요 하다는 점을 시사해 준다.
MAP kinase는 세포 신호전달 분자를 구성하는 중요한 물질이다.29-31 MAP kinase는 하부 경로에 있는 각종 kinase 와 전사물질(transcription factor)과 같은 매개체를 인산화시 킨다. 즉, MAP kinase는 세포 외부에서 들어오는 각종 자극 에 의해 활성화되어 그 신호를 핵으로 전달하여 유전자 발 현을 조절하는 기능을 수행한다. 현재 MAP kinase는 크게 나누어 3종류로 구분될 수 있다. 즉, extracellular signal- regulated kinase (ERK), c-Jun N-terminal kinase (JNK)와 p38 MAP kinase로 구분된다.29-31 ERK MAP kinase는 주로 성장인자(growth factor)와 phorbol ester에 의해 활성화되지 만 염증 자극에 의한 발현은 낮다. 반면 JNK와 p38 MAP kinase는 염증성 cytokine과 스트레스 자극에 의해 강하게 활성화되는 반면 성장인자에 의한 활성은 미약하다.29-31 최 근 H. pylori가 위상피세포에서 MAP kinase를 활성화시킬 수 있다는 보고들이 발표되고 있다.26,32 또한 p38 MAP kinase 경로는 대식세포에서 LPS 자극에 의해 유도되는 iNOS 발현에 관여한다는 보고도 발표되고 있다.33 이 결과 들은 H. pylori 감염에 의해 위상피세포 내에서 활성화되는 MAP kinase 신호전달경로가 iNOS 유전자 발현에 관여할 가 능성이 있음을 시사해 준다. 이와 같은 가정하에 본 연구에 서는 p38 kinase 활성을 차단하는 SB203580과20ERK 인산화 과정을 매개하는 MEK1을 차단하는 PD98059를21 이용하여 MAP kinase 경로를 차단시켰다. 그 결과 SB203580과 PD98059 모두 H. pylori 감염에 의한 위상피세포로부터의 IL-8 및 NO 생성을 감소시켰다. 따라서 p38 및 ERK 활성 화 경로는 H. pylori 감염에 의해 나타나는 iNOS 발현과 NO 생성에 필요하다는 점을 제시해 준다.
331
대한소화기학회지: 제39권 제5호, 2002
최근 MAP kinase와 NF-κB 활성화 경로 사이의 연관성 을 제시한 연구들이 많이 발표되고 있다. 예를 들면 MAP kinase kinase kinase (MEKK1) 또는 NF-κB-inducing kinase (NIK)와 같은 MAP kinase 구성원이 IκB kinase (IKK)를 직접 인산화시켜 NF-κB를 활성화시킨다.34-36 반면 MAP kinase와 NF-κB 활성화 경로는 H. pylori 감염에 의한 IL-8 생성에 있어서 독립적으로 작용한다는 결과가 제시된 바도 있다.26본 연구에서도 MAP kinase 억제제가 NO 생성을 완 전히 차단시키지는 못했다(PD98059에 의해 약 45% 억제, SB203580에 의해 약 43% 억제; Fig. 6). 따라서 H. pylori 감염에 의한 NO 생성에는 NF-κB와 같은 부가적인 경로 가 작용할 것으로 추정되어 이에 대한 연구가 요망된다.
Caco-2 또는 T84와 같은 상피세포주는 위장 내의 점막 표면과 유사한 환경을 제공해주는 모델로 이용되고 있
다.37,38 본 연구에서 H. pylori 감염에 의한 NO는 주로 상피
세포의 apical 방향으로 분비되고 있었다. 반면 호중구를 유 인하는 chemokine인 IL-8은 basolateral 방향으로 분비됨을 확인할 수 있었다. 한편 NO는 H. pylori의 나선형(spiral) 형 태를 구형(coccoid)으로 변화시킨다는 보고가 발표된 바 있 다.39 또한 H. pylori의 나선형 형태만이 위상피세포에 부착 되어 IL-8을 생성할 수 있다는 보고가40 있음에 비추어 apical 방향으로 분비되는 NO는 H. pylori의 기능에 영향을 미칠 것으로 추측된다.
iNOS는 위염에서 상피세포 근처에서 발견되는 호중구와 림프구에서도 관찰된다.28 이와 같은 염증세포는 H. pylori 에 감염된 위상피세포로부터 분비되는 IL-8과 monocyte chemotactic protein (MCP)-1과 같은 chemokine에 의해 점 막하부로 이동한다.41특히 본 연구에서 림프구는 H. pylori 수용성 단백질의 자극에 의해 친염증성 cytokine의 일종인 IFN-γ의 발현을 증가시킬 수 있음을 관찰하였다. 더구나 이 와 같은 IFN-γ는 H. pylori 감염에 의한 NO 생성을 상피세 포의 apical 방향뿐만 아니라 basolateral 방향으로도 증가시 켰다. 이와 같은 결과는 H. pylori에 의해 위상피세포로부터 생성되는 NO가 세균뿐만 아니라 점막 염증반응에도 관여 할 수 있음을 시사해 준다.
결론적으로 H. pylori 감염에 의해 위상피세포로부터 분 비되는 NO는 세균의 활성에 영향을 미칠 뿐만 아니라, 점 막 내부에 존재하는 면역세포와의 상호작용을 통해 염증 반응을 조절할 것으로 생각된다.
요 약
목적: 산화질소(nitric oxide, NO)는 염증반응을 조절할 수 있는 인자 중의 하나로 알려져 있다. 본 연구에서는 H. pylori에 감염된 위상피세포에서 유도성 산화효소 합성
효소(inducible nitric oxide synthase, iNOS)의 발현과 NO 생성의 역할에 대해 연구하였다. 대상 및 방법: 사람 위상 피세포주인 Hs746T 세포에 H. pylori를 감염시킨 뒤 iNOS mRNA 발현을 정량적 RT-PCR로 검사하였다. NO 생성은 배양상청액에 포함된 nitrite 및 nitrate를 측정하여 결정하였 다. NO 분비 방향을 알아보기 위하여 극성을 나타내는 Caco-2 세포를 transwell chamber에 배양한 뒤, 각각의 chamber에 분비된 NO의 양을 측정하였다. 결과: iNOS mRNA와 NO 생성은 H. pylori를 감염시킨 세포에서 유의 하게 증가하였다. MAP kinase 억제제는 H. pylori에 의한 iNOS 발현과 NO 생성을 억제시켰다. 극성을 띠는 Caco-2 세포에 H. pylori를 감염시켰을 때 NO는 주로 apical 방향 으로 분비되었고, IL-8은 주로 basolateral 방향으로 분비되 었다. 한편 H. pylori에 감염된 Caco-2 세포에 IFN-γ를 첨가 했을 경우, NO 생성은 apical 방향뿐만 아니라 basolateral 방향으로도 증가하였다. 결론: H. pylori 감염에 의해 위상 피세포의 apical 방향으로 분비되는 NO는 세균에 영향을 미칠 것으로 보이며, basolateral 방향으로 분비되는 NO는 점막 내부의 염증반응에 관여할 것으로 생각된다.
색인단어: 위상피세포, 헬리코박터 파이로리, 유도성 산화 질소 합성효소
참 고 문 헌
1. Jung HC, Kim JM, Song IS, Kim CY. Helicobacter pylori induces an array of proinflammatory cytokines in human gastric epithelial cells: quantification of mRNA for interleukin-8, -1α/β, granulocyte-macrophage colony-stimulating factor, monocyte chemoattractant protein-1 and tumor necrosis factor-α. J Gastroenterol Hepatol 1997;12:473-480.
2. Kim JM, Kim, JS, Jung HC, Song IS, Kim CY. Upregulated cyclooxygenase-2 inhibits apoptosis of human gastric epithelial cells infected with Helicobacter pylori. Dig Dis Sci 2000;
45:2436-2443.
3. Kim JM, Kim JS, Jung HC, Song IS, Kim CY. Apoptosis of human gastric epithelial cells via caspase-3 activation in response to Helicobacter pylori infection: possible involvement of neutrophils through tumor necrosis factor-alpha and soluble Fas ligands. Scand J Gastroenterol 2000;35:40-48.
4. Kim JS, Jung HC, Kim JM, Song IS, Kim CY. Interleukin-8 expression by human neutrophils activated by Helicobacter pylori soluble proteins. Scand J Gastroenterol 1998:33;1249- 1255.
5. Kim JS, Jung HC, Kim JM, Song IS, Kim CY. Helicobacter pylori water-soluble surface proteins activate human neutrophils 332
김정목 외 4인. H. pylori에 의한 iNOS 발현
and upregulate the expression of CXC chemokines. Dig Dis Sci 2000;45:83-92.
6. Ihan A, Tepez B, Gubina M, Malovrh T, Kopitar A.
Diminished interferon-γ production in gastric mucosa T lymphocytes after Helicobacter pylori eradication in duodenal ulcer patients. Hepatogastroenterology 1999;46:1740-1745.
7. Bamford KB, Fan X, Crowe SE, et al. Lymphocytes in the human gastric mucosa during Helicobacter pylori have a T helper cell 1 phenotype. Gastroenterology 1998;114:482-492.
8. Nathan C, Xie QW. Nitric oxide synthases: roles, tolls, and controls. Cell 1994;78:915-918.
9. Nathan C, Xie QW. Regulation of biosynthesis of nitric oxide.
J Biol Chem 1994;269:13725-13728.
10. Witthoft T, Eckmann L, Kim JM, Kagnoff MF. Enteroinvasive bacteria directly activate expression of iNOS and NO production in human colon epithelial cells. Am J Physiol 1998;275:G564-G571.
11. Kolios G, Brown Z, Robson RL, Robertson DA, Westwick J.
Inducible nitric oxide synthase activity and expression in a human colonic epithelial cell line, HT-29. Br J Pharmacol 1995;116:2866-2872.
12. Linn SC, Morelli PJ, Edry I, Cottongim SE, Szabo C, Salzman AL. Transcriptional regulation of human inducible nitric oxide synthase gene in an intestinal epithelial cell line. Am J Physiol 1997;272:G1499-G1508.
13. Fu S, Ramanujam KS, Wong A, et al. Increased expression and cellular localization of inducible nitric oxide synthase and cyclooxygenase 2 in Helicobacter pylori gastritis. Gastroent- erology 1999;116:1319-1329.
14. Sakaguchi AA, Miura S, Takeuchi T, et al. Increased expression of inducible nitric oxide synthase and peroxynitrite in Helicobacter pylori gastric ulcer. Free Radic Biol Med 1999;27:781-789.
15. Slomiany BL, Piotrowski J, Slomiany A. Induction of caspase-3 and nitric oxide synthase-2 during gastric mucosal inflammatory reaction to Helicobacter pylori lipopolysaccharide.
Biochem Mol Biol Int 1998;46:1063-1070.
16. Itoh T, Wakatsuki Y, Yoshida M, et al. The vast majority of gastric T cells are polarized to produce T helper 1 type cytokines upon antigenic stimulation despite the absence of Helicobacter pylori infection. J Gastroenterol 1999;34:560-570.
17. Salzman A, Denenberg AG, Ueta I, OConnor M, Linn SC, Szabo C. Induction and activity of nitric oxide synthase in cultured human intestinal epithelial monolayers. Am J Physiol 1996;270:G565-G573.
18. Kim JM, Kim, JS, Jung HC, Song IS, Kim CY. Virulence factors of Helicobacter pylori in Korean isolates do not
influence on proinflammatory cytokine gene expression and apoptosis of human gastric epithelial cells, nor do these factors influence the clinical outcome. J Gastroenterol 2000;35:
898-906.
19. Kim JS, Kim JM, Jung HC, Song IS. Caspase-3 activity and expression of Bcl-2 family in human neutrophils by Helicobacter pylori water-soluble proteins. Helicobacter 2001;6:207-215.
20. Badger AM, Bradbeer JN, Votta B, Lee JC, Adams JL, Griswold DE. Pharmacological profile of SB 203580, a selective inhibitor of cytokine suppressive binding protein/p38 kinase, in animal models of arthritis, bone resorption, endotoxin shock and immune function. J Pharmacol Exp Ther 1996;279:
1453-1461.
21. Dudley DT, Pang L, Decker SJ, Bridges AJ, Saltiel AR. A synthetic inhibitor of the mitogen-activated protein kinase cascade. Proc Natl Acad Sci USA 1995;92:7686-7689.
22. 김정목, 김주성, 정현채, 송인성, 김정룡. Clostridium difficile toxin A에 의한 장상피세포에서의 CXC 및 CC chemokine의 발현. 대한소화기학회지 2001;37:345-355.
23. Tse WY, Williams J, Pall A, Wilkes M, Savage CO, Adu D.
Antineutrophil cytoplasm antibody-induced neutrophil nitric oxide production is nitric oxide synthase independent. Kidney Int 2001;59:593-600.
24. Gradini R, Realacci M, Ginepri A, et al. Nitric oxide synthases in normal and benign hyperplastic human prostate: immuno- histochemistry and molecular biology. J Pathol 1999;189:224-229.
25. 김정목, 김주성, 정현채, 고은주, 송인성, 김정룡. Helico- bacter pylori 병독인자에 의한 인체 위상피세포의 친염증성 cytokine 발현 및 apoptosis 유도와 임상 질환과의 관계. 대 한소화기학회지 2000;36:583-596.
26. Keates S, Keates AC, Warny M, Peek RM Jr, Murray PG, Kelly CP. Differential activation of mitogen-activated protein kinases in AGS gastric epithelial cells by cag+ and cag- Helicobacter pylori. J Immunol 1999;163:5552-5559.
27. Hofer D, Jons T, Kraemer J, Drenckhahn D. From cytoskeleton to polarity and chemoreception in the gut epithelium. Ann N Y Acad Sci 1998;859:75-84.
28. Mannick EE, Bravo LE, Zarama G, et al. Inducible nitric oxide synthase, nitrotyrosine, and apoptosis in Helicobacter pylori gastritis: effect of antibiotics and antioxidants. Cancer Res 1996;56:3238-3243.
29. Han J, Lee JD, Bibbs L, Ulevitch RJ. A MAP kinase targeted by endotoxin and hyperosmolarity in mammalian cells. Science 1994;265:808-811.
30. Kyriakis JM, Banerjee P, Nikolakaki E, et al. The stress- activated protein kinase subfamily of c-Jun kinases. Nature 1994;369:156-160.
333
The Korean Journal of Gastroenterology: Vol. 39, No. 5, 2002
31. Lee JC, Laydon JT, McDonnell PC, et al. A protein kinase involved in the regulation of inflammatory cytokine biosyn- thesis. Nature 1994;372:739-246.
32. Meyer-ter-Vehn T, Covacci A, Kist M, Pahl HL. Helicobacter pylori activates mitogen-activated protein kinase cascades and induces expression of the protooncogenes c-fos and c-jun. J Biol Chem 2000;275:16064-16072.
33. Jeon YJ, Kim YK, Lee M, Park SM, Han SB, Kim HM.
Radicicol suppresses expression of inducible nitric-oxide synthase by blocking p38 kinase and nuclear factor-kappa B/Rel in lipopolysaccharide-stimulated macrophages. J Pharmacol Exp Ther 2000;294:548-554.
34. Nemoto S, DiDonato JA, Lin A. Coordinate regulation of IkB kinases by mitogen- activated protein kinase kinase kinase 1 and NF-κB-inducing kinase. Mol Cell Biol 1998;18:
7336-7343.
35. Lee FS, Peters RT, Dang LC, Maniatis T. MEKK1 activates both IκB kinase alpha and IκB kinase beta. Proc Natl Acad Sci USA 1998;95:9319-9324.
36. Mercurio F, Zhu H, Murray BW, et al. IKK-1 and IKK-2:
cytokine-activated IκB kinases essential for NF-κB activation. Science 1997;278:860-866.
37. Papini E, Satin B, Norais N, et al. Selective increase of the permeability of polarized epithelial cell monolayers by Helico- bacter pylori vacuolating toxin. J Clin Invest 1998;102:
813-820.
38. Hofman V, Ricci V, Galmiche A, et al. Effect of Helicobacter pylori on polymorphonuclear leukocyte migration across polarized T84 epithelial cell monolayers: role of vacuolating toxin VacA and cag pathogenicity island. Infect Immun 2000;68:5225-5233.
39. Cole SP, Cirillo D, Kagnoff MF, Guiney DG, Eckmann L.
Coccoid and spiral Helicobacter pylori differ in their abilities to adhere to gastric epithelial cells and induce interleukin-8 secretion. Infect Immun 1997;65:843-846.
40. Cole SP, Kharitonov VF, Guiney DG. Effect of nitric oxide on Helicobacter pylori morphology. J Infect Dis 1999;180:
1713-1717.
41. Bodger K, Crabtree JE. Helicobacter pylori and gastric inflammation. Br Med Bull 1998;54:139-150.
334