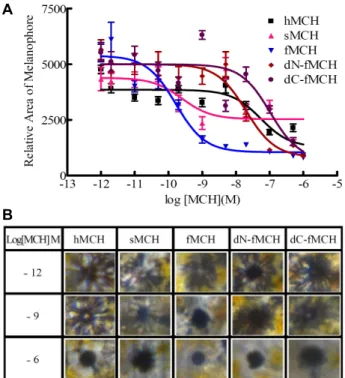
Characterization of Melanin-concentrating Hormone from Olive Flounder (Paralichthys olivaceus)
In Young Chung
1, Jeong Min Jeon
2and Young Hwan Song
1*
1
Department of Microbiology, Pukyong National University, Busan 46041, Korea
2
전체 글
1
2
수치


관련 문서
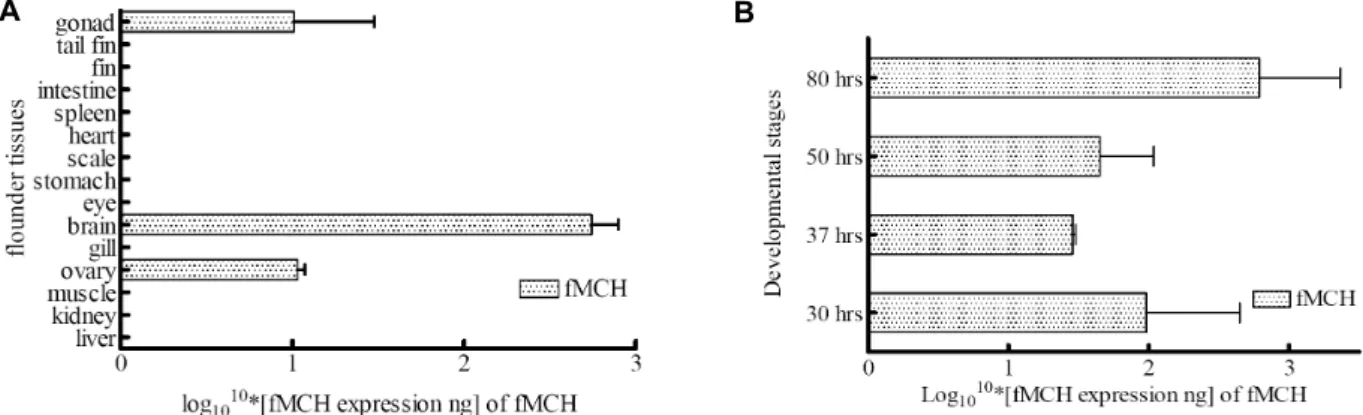
in internal organs (kidney, intestine, spleen, brain and liver) from non-emaciated (farm-A) or emaciated (farm-B and farm-C) olive flounder Paralichthys olivaceus
These results suggest that circulating miR-122 might be used as a sensitive indicator as well as GOT and GOT levels in evaluat- ing liver injury in olive
Detection of RSIV (red sea bream iridovirus), VHS (viral heamorrhagic septiceamia) virus, and VNN (viral nervous necrosis) virus in infected olive flounder tissues by
In the present study, polyclonal antiserum against olive flounder (Paralichthys olivaceus) IgM heavy chain was generated by intramuscular immuniza- tion of rabbit with
Hispopathologic characterization of viral pathogens in cultured olive flounder, Paralichthys olivaceus, using in-situ hybridization methods, Journal of Fish Pathology
In the present study, isoeugenol as new anesthetic in marine fish, especially olive flounder, was examined to know the efficacy and proper concentration.. In acute toxicity test,
In the present study, to evaluate the effect of immunosuppression on MABV bur- den in fish, fingerlings of olive flounder naturally infected with MABV were injected
Optimum feed- ing rate for growing olive flounder (317 g) Paralichthys olivaceus fed practical extruded pellets at optimum water temperature (21-24℃). Evaluation
