된장에서 분리된 Bacillus licheniformis의 β-galactosidase 생산성과 효소특성
진현경, 윤기홍*
우송대학교바이오식품과학전공
Received: October 15, 2014 / Revised: November 25, 2014 / Accepted: November 25, 2014
서 론
β-galactosidase는미생물을비롯하여동식물에두루존 재하며 lactose를 glucose와 galactose로분해하는효소이고 식품산업에 활용성이 높다. 우유, 유제품과 유장으로부터 lactose를제거하거나[2] 장내유용미생물의성장을증진시키 는갈락토올리고당[26]을비롯한 lactulose [6], lactosucrose [16]의생산에사용되며또한대장균유래의β-galactosidase 가작용하지못하는재조합숙주균에서표지효소로이용되 고있다[27].
효소의생산균주에따라서β-galactosidase의 lactose 가 수분해능과당전이효율을비롯하여반응특성이다르므로 산업적활용에적합한효소특성과생산성을확보하기위해 내열성이나 저온성 균주를 비롯한 여러 균주로부터 β-
galactosidase가연구되었다[22]. Bacillus stearothermophilus [8], B. circulans [26]와 B. coagulans [1]는 3종이상의 β- galactosidase isozymes를생산하는데, 온천에서분리된 B.
coagulans RCS3의 isozyme III은다른 isozymes와는달리
균체외로분비되며 lactose 분해능이우수한것으로확인되
었다. 또한 B. circulans의효소는 isozymes간에면역학적으
로동일하나 lactose 분해능과당전이에의한갈락토올리고
당 생산능에는 차이가 큰 것으로 보고되었다. 한편 B.
licheniformis의 β-galactosidase는 당 전이효율은 낮으나 lactose 분해에적합한효소를생산하는것으로알려졌다[12].
Bacillus속균주중 B. megaterium [23], B. licheniformis [12], B. stearothermophilus [5, 8]와 B. coagulans [1]로부 터β-galactosidase 효소와유전자의특성이보고되었으며, 북극에서분리된 Alkalilactibacillus ikkense로부터저온에 서활성이높은 β-galactosidase의유전자도클로닝되었다 [22]. β-galactosidase의활용성을증대시키기위해서단백질
의구조를기반으로하여 lactose 가수분해능이향상된변이체
가개발되었으며[5], 담체에고정화된 B. stearothermophilus와 B. circulans 유래의효소가당전이반응과 lactose 분해반응 Production and Characterization of β-galactosidase from Bacillus licheniformis Isolated from Doenjang
Hyun Kyung Jin and Ki-Hong Yoon*
Food Science & Biotechnology Major, Woosong University, Daejeon 300-718, Republic of Korea
A bacterial strain was isolated from homemade doenjang (Korean fermented soybean paste) as a producer of the extracellular β-galactosidase, capable of hydrolyzing lactose to liberate galactose and glucose residues. The isolate YB-1414 has been identified as Bacillus licheniformis on the basis of its 16S rDNA sequence, morphology and biochemical properties. The pro- duction of β-galactosidase by B. licheniformis YB-1414 reached maximum levels of 6.2 U/ml in culture medium containing wheat bran (1%) and yeast extract (2.5%) as carbon and nitrogen sources, respectively. Particularly, the insoluble fraction was more effective for β-galactosidase production than the soluble extract of wheat bran. The enzyme exhibited maximum activity for hydrolysis of para-nitrophenyl-β-D-galactopyranoside (pNP-βGal) under reaction conditions of pH 6.0 and 55-60oC. Its hydrolyzing activity for pNP-βGal was drastically decreased by the addition of low concentrations of galactose, but only slightly decreased by glucose, with 85% of maximal activity in the presence of 400 mM glucose.
Keywords: Bacillus licheniformis, β-galactosidase, productivity, property
*Corresponding author
Tel: +82-42-630-9742, Fax: +82-42-636-2676 E-mail: [email protected]
© 2014, The Korean Society for Microbiology and Biotechnology
에사용되었다[3, 7]. β-galactosidase의생산성을증가시키 기 위해서는 B. stearothermophilus의 효소 유전자를 B.
subtilis [9] 또는 B. amyloliquefaciens [10]에서발현시켰다. 또한 B. subtilis [13]와 B. licheniformis [18]의β-galactosidase 생산성에영향을미치는배지성분에대한연구도수행되었다. 최근들어청국장의기능성에관심이높아지면서생청국 장이나건조된청국장의분말또는환의형태로유유와혼 합하여건강식으로이용되고있다. β-galactosidase 활성이 높은청국장은우유와함께섭취할경우유당불내증을방 지하는효과가예상되므로β-galactosidase를생산하는미생 물을청국장발효균으로활용하기위해청국장으로부터 B.
licheniformis가분리된바있으며[28], 본연구에서는된장 으로부터β-galactosidase를생산하는 Bacillus속균주중효 소생산성이우수한미생물을분리하여반응특성을조사하 였다.
재료 및 방법
β-galactosidase 생산균의 탐색
Spizizen 최소배지[25]의 탄소원인 포도당과 sodium citrate 및질소원인 ammonium sulfate 대신에대두분 (1%) 이첨가된변형배지(K2HPO4, 7 g; KH2PO4, 1.5 g; MgSO4· 7H2O 0.5 g; MnCl2· 4H2O, 0.04 g; CaCl2· 2H2O, 0.003 g;
FeSO4·7H2O, 0.0025 g; (NH4)6Mo7O24·4H2O, 0.002 g; water, 1 L)에가정에서수집된된장을접종하여 37oC에서약 3일간 진탕배양하고동일한배지에 3-4일간격으로 3회계대배 양을실시하였다. 최종적으로계대배양액을동일한성분의 평판배지에도말하고 37oC에서 4일간배양한후형성된콜 로니중에서서로다른모양을보이는콜로니를채취하였다. 이들분리균을 LB 액체배지에서 24시간동안진탕배양한 후 배양상등액을 회수하여 1 mM para-nitrophenyl-β-D- galactopyranoside (pNP-βGal; Sigma, USA)을 포함한 20 mM sodium phosphate 완충액(pH 6.0)에 첨가하고
45oC에서 2시간동안방치한후반응액의색깔을관찰함으
로써β-galactosidase 활성을갖는균을탐색하였다. 분리균주의 동정
그람염색과포자염색을통해분리균의형태를조사하였으 며, 균체현탁액을 API 20E와 API 50CHB (Biomereux사, France) kits에제조사의지침을따라접종하고 37oC에서배 양하면서 1일과 2일째각각관찰하여탄수화물이용능과생 화학적특성을판별하였다. 분리균의총염색체 DNA를주 형으로하고, 세균의 16S rRNA 유전자의보존적지역의염 기서열 5'-AGAGTTTGATCCTGGCTCAG-3' (E. coli 16S rRNA 유전자염기서열 8-27), 5'-GGTTACCTTGTTACGACTT-
3' (E. coli 16S rRNA 유전자염기서열 1492-1510)을 primer로
사용하여중합효소연쇄반응(PCR)을실시한후증폭된 PCR
산물을정제하여분리균의 16S rDNA 염기서열을분석하였다. β-galactosidase 조효소액 제조
분리균 B. licheniformis YB-1414를 LB 배지에서 16시간
동안배양한배양액을 1% (v/v)가되도록동일성분의본배
양액에접종하고 37oC에서 36시간동안진탕배양하였다. 원 심분리하여얻은배양상등액을 ammonium sulfate (25-75%) 로처리하고침전된단백질을 10 mM sodium phosphate 완충
액(pH 6.0)에현탁하여동일완충액으로투석한후조효소
액으로사용하였다.
β-galactosidase 반응특성 분석
β-galactosidase 활성을결정하기위해서는 1 mM pNP- βGal과 50 mM sodium phosphate 완충액(pH 6.0)을포함 한반응액에효소를첨가하여 50oC에서 10분간반응시킨후 반응액의 2배부피의 1 M Na2CO3 용액을첨가하여반응을 종결시키고 405 nm에서 흡광도를 측정하였다. para- nitrophenol (pNP)을표준물질로하여얻은검량곡선을사 용하여효소반응에의해생성된 pNP의양을결정하였다. 효소의활성도 1 unit는 1분동안 1 μmol의 pNP를유리시 키는효소량으로정의하였다. 효소활성에미치는반응온도 와 pH의영향을조사하기위하여 30-70oC와 pH 5.0-8.0의 범위에서 각각 β-galactosidase 활성을 측정하였다. β-
galactosidase의열안정성을조사하기위하여조효소액을
서로다른온도에서일정시간방치한후잔존활성을측정하 였다. β-galactosidase에의한최종가수분해산물을조사하 기위해 lactose (1%)와과량의조효소액을포함한반응액을 40oC에서 5시간반응시킨후반응액을열처리하여단백질을 제거하고상등액을취해 chloroform, acetic acid와증류수 (4.3 : 5 : 0.7, (v/v)) 혼합용액을 전개용액으로하여 silica gel-precoated thin layer plate (Merck, Gemany)에서박층 크로마토그래피를수행하였다. 전개된물질을발색시키기위 해서는 9 ml ethanol, 0.5 ml p-anisaldehyde, 0.5 ml sulfuric acid와 glacial acetic acid 몇방울을혼합한발색제용액을 뿌린후, 120oC에서 10분간방치하였다. 당이효소활성에
미치는영향을분석하기위하여기질로 pNP-βGal을포함한
반응액에종류와농도가다르게당을첨가한상태에서반응 을수행한후β-galactosidase 활성을측정하였다.
결과 및 고찰
β-galactosidase 생산균의 분리와 특성
된장으로부터고분자물질의가수분해효소를생산하는 B.
subtilis, B. licheniformis 및 B. amyloliquefaciens가분리 된바있으며[11, 29], 여러종류의 Bacillus속및이와관련 된균이발효장류에다수존재하는것으로보고되었다[19].
된장을직접복합평판배지에도말하여균주를분리할경우 우점균이나한정된균종만빠르게성장할수있으므로[17]
탄소원과질소원대신대두분을첨가한최소배지를사용하 여계대배양과분리과정을실시하고평판배지에서형성된 콜로니의모양이서로다른균을분리하였다. 가정에서제조 된된장 60점시료로부터균을분리한결과된장시료당약 1-4개의균주가분리되었다. 총 150개분리균을 LB 액체배 지에배양하여얻은배양상등액의β-galactosidase 활성을조 사하였으며, 그결과 5개분리균주를제외하고는대부분이 β-galactosidase 활성이관찰되지않거나미약한것으로나 타났다. 따라서효소생산성이우수한 5개분리균의배양상 등액내효소활성을비교하여최종적으로β-galactosidase 활 성이높은분리균 YB-1414를확보하였다.
분리균 YB-1414는포자를형성하는그람양성간균이며,
API 50CHB와 20E kit를사용하여조사한생화학적특성을 Biomeriux의 API web (https://apiweb.biomerieux.com/
jsp/login.jsp)에서다른균주와비교한결과 B. licheniformis
와유사도가 99.9%로가장높게 나타났고당이용성도 B.
licheniformis와동일하였다. 분리균은 arginine dihydrolase, gelatinase와 oxidase 활성을 보였고, nitrate 환원능과 citrate 이용능이있으며 acetoin을생성하였다. 또한 lysine decarboxylase, ornithine decarboxylase, urease와 tryptophan deaminase의활성은없고, indole과황화수소를생성하지못 하였다. 분리균의 16S rRNA 유전자를 PCR로 증폭한 후 1,430 bp 크기의염기서열을결정하였으며(Genbank accession No. KM660627) 이를미국 NCBI의 BLAST 검색방법을사 용하여세균들의상응하는염기서열과비교하였다. 그결과, B. licheniformis DSM 13 (NR_118996)과 BCRC 11702 (NR_116025)의 16S rDNA 서열과가장상동성이높았으며, 2개의염기를제외하고는동일하였다. 이상의형태적, 생화 학적특성및 16S rDNA 서열의결과로볼때분리균 YB- 1414는 B. licheniformis인 것으로 판단된다. 분리균 YB-
1414의고분자 물질의 분해능을조사하기위해 1% skim
milk, 0.2% potato starch, 0.5% xylan과 0.5% carboxymethyl
cellulose를각각첨가한평판배지에서하룻밤배양하여분
해환을관찰하였으며그결과분리균은 xylan을분해하지못 하였고나머지물질을모두분해하는것으로나타났다. 한편 청국장에서도β-galactosidase를생산하는 B. licheniformis 가분리된바있는데[28], 분리균 YB-1414는청국장에서분 리된β-galactosidase 생산균과는달리 acetoin을생성하고, citrate, rhamnose, sorbitol, melibiose, inuline과 β-gentiobiose 를탄소원으로이용하는차이점을보였다.
B. licheniformis YB-1414의 β-galactosidase 생산성 β-galactosidase는균체내효소로존재하는경우가많으며 B. stearothermophilus [4, 8], B. megaterium ATCC 14581 [23], B. subtilis KL88 [20]에서균체내효소로보고 된바있다. 또한청국장에서 분리된 B. licheniformis YB- 1105도배양상등액보다균체내에β-galactosidase 활성이
높은 것으로 알려졌다[28]. 한편 면양유에서 분리된 B.
subtilis [13]와온천에서분리된 Bacillus sp. [21]는균체외 로β-galactosidase를분비생산하며, B. coagulans RCS3의 경우 5개의β-galactosidase isozymes 중한개가배양상등 액에서관찰된바있다[1]. 분리균 B. licheniformis YB-1414 도배양상등액에서β-galactosidase가높은활성으로관찰되 므로균의성장과효소생산성과의관계를조사하기위해 LB 액체배지에서진탕배양하면서일정시간마다배양액을채취
하여 600 nm에서 흡광도와 배양상등액에 존재하는 β-
galactosidase의활성을측정하였다.
B. licheniformis YB-1414는배양시간이 12시간되었을때 최대성장도에이르렀고이때부터 β-galactosidase 생산이시 작되어정지기를지나사멸기에도달한후까지효소생산성 이지속적으로증가하였으며 30시간이되었을때 2.8 U/ml 로최대에이르렀고이후에도배양상등액에존재하는효소 활성이감소하지않고그대로유지되었다(Fig. 1). 균체외 β- galactosidase를생산하는 Bacillus sp.의경우도효소생산 성이정지기이후에지속적으로증가되는현상을보여분리 균과유사하였으며, 이러한현상은균체내에서합성된효소 가균체외로확산되는데걸리는시간의차이때문인것으로
Fig. 1. Growth and β-galactosidase production of B. licheni- formis YB-1414. B. licheniformis YB-1414 was grown at 37oC in LB medium. The cell growth (-●-) was determined by measuring absorbance of the cell culture at wavelength of 600 nm. β-galac- tosidase activities (-○-) were determined with the culture filtrate.
The curve for β-galactosidase production represents the average of three independent experiments within standard errors of 1.5%
between them.
추정된 바 있다[21]. 또한 B. stearothermophilus의 β- galactosidase 유전자를함유한 B. subtilis 재조합균주는균 체내β-galactosidase를생산하므로효소생산성이균의성 장과연계된형태로정지기에이를때까지지속적으로증가 하여최대생산성이약 6.3 U/ml로나타났다[9].
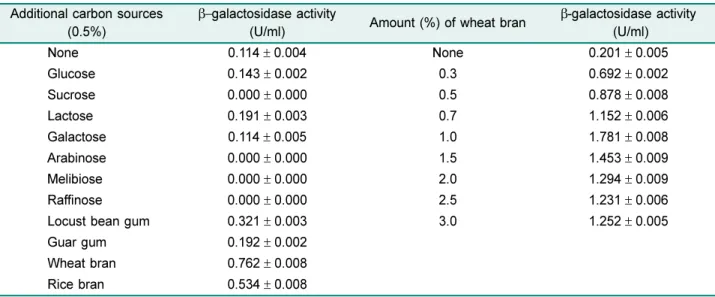
β-galactosidase 생산성은배지내탄소원과질소원성분에 따라영향을받는다고알려져있으므로 potassium phosphate (0.01%)와 magnesium sulfate (0.1%)를기본성분으로하고
탄소원과질소원을달리하여분리균 YB-1414의효소생산
성을조사하였다. 질소원으로 peptone (0.5%)을함유하는배
지에서로다른탄소원을 0.5%가되도록각각첨가한상태
에서분리균을접종하여 37oC에서 36시간동안진탕배양한 후배양상등액의효소활성을측정한결과 Table 1에보인 바와같이밀기울을비롯하여쌀기울, locust bean gum 또
는 guar gum을첨가한배지에서효소생산성이증가되었으
며 glucose와 lactose를비롯한단당류나이당류가첨가된배 지에서는탄소원을첨가하지않은배지보다효소생산성이
감소하였거나큰변화가없는것으로나타났다. 이는쉽게 대사되는 glucose와 lactose보다귀리가루, 밀기울과쌀기울 이함유된배지에서 B. subtilis의β-galactosidase 생산성이증 가되거나, 왕겨를첨가한고체배지에서 B. licheniformis의β- galactosidase 생산성이증가한현상과유사하지만[13, 18], B. megaterium [23]과 B. stearothermophilus [9] 및 B.
stearothermophilus의β-galactosidase 유전자가 함유된재 조합 B. amyloliquefaciens [10]에서효소생산이 lactose에 의해유도된다는결과와는달랐다. 또한밀기울을첨가한배 지에서효소생산성이가장높았으므로밀기울의농도를달 리한배지에서동일조건으로배양하였을때밀기울첨가량 이 1%가될때까지는효소생산성이증가되었으나 1.5% 이 상의밀기울이첨가되었을때는효소생산성이약간감소하 는현상을보였다(Table 1).
탄소원으로밀기울첨가량을 1%로고정하고유기질소원
(0.5%)의종류를달리한배지에서효소생산성을조사하였
을때 yeast extract 또는 tryptone이첨가된배지에서효소
Table 1. Effects of additional carbon sources on the β-galactosidase production from B. licheniformis YB-1414 Additional carbon sources
(0.5%)
β−galactosidase activity
(U/ml) Amount (%) of wheat bran β-galactosidase activity (U/ml)
None 0.114± 0.004 None 0.201± 0.005
Glucose 0.143± 0.002 0.3 0.692± 0.002
Sucrose 0.000± 0.000 0.5 0.878± 0.008
Lactose 0.191± 0.003 0.7 1.152± 0.006
Galactose 0.114± 0.005 1.0 1.781± 0.008
Arabinose 0.000± 0.000 1.5 1.453± 0.009
Melibiose 0.000± 0.000 2.0 1.294± 0.009
Raffinose 0.000± 0.000 2.5 1.231± 0.006
Locust bean gum 0.321± 0.003 3.0 1.252± 0.005
Guar gum 0.192± 0.002
Wheat bran 0.762± 0.008
Rice bran 0.534± 0.008
Table 2. Effects of additional nitrogen sources on the β-galactosidase production from B. licheniformis YB-1414 Additional nitrogen sources
(0.5%)
β-galactosidase activity
(U/ml) Amount (%) of yeast extract β-galactosidase activity (U/ml)
None 0.083± 0.005 0.0 0.042± 0.003
Corn steep powder 0.003± 0.002 0.5 1.485± 0.009
Soytone 0.851± 0.001 0.8 2.385± 0.010
Yeast extract 1.312± 0.004 1.0 3.092± 0.010
Tryptone 1.153± 0.009 1.5 4.793± 0.017
Peptone 0.884± 0.009 2.0 6.152± 0.001
Casein hydrolyzate 0.742± 0.005 2.5 6.205± 0.014
3.0 5.523± 0.011
생산성이높았으며, soytone, peptone 또는 casein 가수분해 물이첨가된배지에서는서로유사한수준의효소생산성을 보였고특이하게도 corn steep powder가함유된배지에서는
효소가거의생산되지않았다(Table 2). 효소생산성을가장
크게 증가시킨 yeast extract의 첨가량을 달리하였을 때
yeast extract의첨가량이많아질수록β-galactosidase 생산
성이증가하여 2.5%에서최대생산성을보이고 3.0%에서는
약간감소하였다. B. subtilis도분리균 YB-1414와유사하게 yeast extract를질소원으로사용하였을때효소생산성이가 장높았으나, 분리균과는달리 peptone이 tryptone 보다는 효소생산능을더증가시키는것으로보고되었다[13].
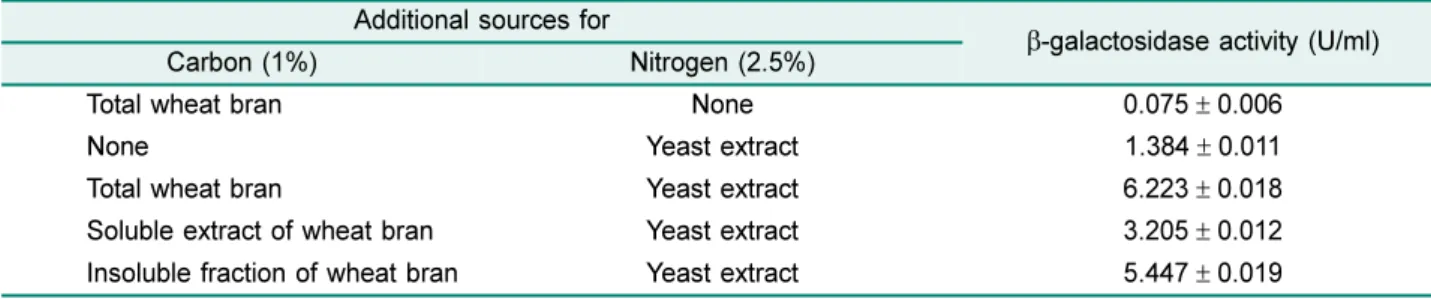
한편밀기울의불용성물질과수용성물질중어느것이 효소생산성을증가시키는지확인하기위해서밀기울 1%를 증류수에현탁하고원심분리후불용성의침전물과수용성 의상등액부분으로나누고 yeast extract (2.5%)와기본성 분을각각첨가하여제조된배지에서효소생산성을비교하
였다. Table 3에보인바와같이밀기울의불용성물질이첨
가된배지에서효소생산성은밀기울이그대로첨가되었을 때보다약간낮은것으로나타났지만수용성물질이첨가된 배지에서보다더높았다. 또한밀기울의수용성물질이함 유된 배지에서의 효소 생산성은밀기울이 첨가되지않고 yeast extract만첨가된배지에서보다 2배이상높은것으 로나타났다. 이로보아β-galactosidase의생산성을증가시 키는물질이밀기울의불용성물질과수용성물질에모두있 으며불용성물질에좀더많이존재하는것으로판단된다. 한편 yeast extract를첨가하지않고밀기울만을첨가한배 지에서는효소생산성이매우낮았는데이는질소원이거의 없어균의성장이낮아진때문으로여겨진다.
B. licheniformis의 β-galactosidase의 반응특성 배양상등액을 ammonium sulfate (25-75%)로처리하여제 조된조효소액을사용하여반응조건에따른β-galactosidase 활성을측정하였다. 그결과 pH 6.0과 55-60oC에서최대활 성을나타냈으며, pH 5.5-6.5 범위에서는최대활성의 90%
이상의활성을보였다(Fig. 2). 분리균효소의최적반응온
도는 50oC에서최대 효소활성을 보이는 B. licheniformis
DSM 13 [12]과 B. licheniformis YB-1105 [28]의 효소에 비해 높았으며, B. coagulans L4 [14]와 B. megaterium 2- 37-4-1 [15]의효소와는유사하였고, 최적반응온도가 65oC 와 70oC로보고된 B. coagulans RCS3 [2], B. subtilis [13]
와 B. stearothermophilus [4]의효소보다는낮았다. 한편북
극에서 분리된 A. ikkense가 생산하는저온활성 효소는
20-30oC에서최대 활성을보이는것으로보고되었다[22].
또한 B. licheniformis DSM 13, B. coagulans [1]와 B.
stearothermophilus [4] 유래효소의최적 pH는 6.0-7.0 범위 로알려져분리균의효소와유사하였으며, B. megaterium 2-37-4-1과 A. ikkense의효소 및 B. stearothermophilus의 변이체효소[5]는 pH 7.5-8의범위에서최대활성을보이는 것으로보고되었다.
30-60oC 범위의온도에서조효소액을 30분과 60분동안 Table 3. β-galactosidase production from B. licheniformis YB-1414 according to soluble and insoluble fraction of wheat bran
Additional sources for
β-galactosidase activity (U/ml)
Carbon (1%) Nitrogen (2.5%)
Total wheat bran None 0.075± 0.006
None Yeast extract 1.384± 0.011
Total wheat bran Yeast extract 6.223± 0.018
Soluble extract of wheat bran Yeast extract 3.205± 0.012
Insoluble fraction of wheat bran Yeast extract 5.447± 0.019
Fig. 2. Effects of reaction temperature and pH on the β-galac- tosidase activity. Temperature profile (triangles) was obtained by measuring the β-galactosidase activities at different temperatures and pH 6.0. The reactions were done at 50oC and various pHs for determining the pH profile (circles). Buffers used were as follows:
sodium citrate (-●-), sodium phosphate (-○-). Each curve rep- resents the average of three independent experiments within stan- dard errors of 2% between them.
각각열처리한후잔존활성을조사한결과 Fig. 3에보인바 와같이 40oC 이하에서는 60분동안방치하여도안정하였 으나, 45oC 이상의 온도에서는 1시간 방치하였을 때 β- galactosidase는서서히실활되기시작하였으며 60oC에서는 30분방치하였을때도잔존활성이거의관찰되지않는것으 로보아 60oC 이상에서는매우불안정한것으로판단된다. 그러므로분리균의β-galactosidase는 55oC에서반감기가 94 시간인 Bacillus sp.의효소[21], 65oC에서반감기가 2시간인 B. coagulans RCS3의 효소와 반감기가 50시간인 B.
stearothermophilus ATCC 8005의효소에비해열안정성이 낮으며[2, 4], 55oC에서 30분만에 70%가실활된 B. coagulans L4의 효소와 45oC에서 1시간만에 70%가 실활된 B.
licheniformis YB-1105의균체외효소보다는열안정성이높 았다[14, 28].
pNP-βGal의가수분해능이있는β-galactosidase 중에서 실제 lactose를거의분해하지못하는경우도있으므로[24]
분리균이생산하는 β-galactosidase의 lactose 가수분해산물 을 TLC로 분석하였다. 그 결과 Fig. 4에 보인 바와 같이 lactose가완전히분해되지는않았지만, 분해산물로 galactose
와 glucose의이동도가동일한반응산물이생성된것으로보
아 lactose를분해하는것을알수있다. 실제β-galactosidase 는 lactose의분해산물인 galactose 또는 glucose에의해가 수분해활성이저해를받는것으로알려져있는데, 반응산
물에상당량의 lactose가분해되지못한상태인것으로보아
YB-1414의β-galactosidase도 lactose 가수분해과정에서반 응산물에의해크게저해를받은것으로여겨진다.
β-galactosidase의 가수분해 활성에 미치는 당의 영향 β-galactosidase, β-glucosidase, β-xylosidase와 α- galactosidase 등을포함하는여러종류의 glycosidase는가 수분해반응시반응산물에의해효소활성이저해되는것으 로알려져있다. 따라서 YB-1414의β-galactosidase 활성에 미치는당의영향을분석하기위해 1 mM pNP-βGal을기질 로하고 glucose, galactose, xylose, mannose의첨가농도를 달리하여가수분해활성을측정하였다. 그결과 galactose에 의한가수분해활성의저해도가가장큰것으로나타났으며,
20 mM 이상이존재할경우효소활성이 50% 이상저해되었
다(Fig. 5). B. coagulans RCS3 [2], B. stearothermophilus [9], B. licheniformis DSM 13 [12]과 B. licheniformis YB- 1105 [28]의효소도 galactose에의해활성이강력하게저해 를받는것으로알려져있으나, B. subtilis KL88의효소는 galactose가 20% 존재할때도 50% 이상의β-galactosidase 활성을유지하는것으로보고되었다[16].
Glucose는 galactose에비해분리균의효소활성을약하게 저해하였으며, 400 mM의존재하에서도약 85%의활성을 보였다. Glucose와유사한 mannose도효소활성을약하게저 해하였으며, 400 mM 존재하에서약 91% 이상의활성을보 였다. β-galactosidase에의한 lactose 분해산물로 glucose와 galactose가동시에생성되는데 glucose 보다 galactose에의 한효소활성저해도가큰것은 galactose는효소의활성부위
에경쟁적저해제로작용하는반면에 glucose는비경쟁적저
해제로작용하기때문인것으로알려졌다[16]. 그러므로대
부분의β-galactosidase는분리균의효소와유사하게 glucose Fig. 3. Thermostability of the β-galactosidase in culrue filtrate.
Thermostability was determined by measuring the residual activi- ties of β-galactosidase after pre-incubations for 30 min (-●-) and 1 h (-○-) at the different temperatures. Each curve represents the average of three independent experiments within standard errors of 2% between them.
Fig. 4. Thin-layer chromatogram of hydrolysis product of lactose. Reactions were done using lactose as a substrate by culture filtrate (lanes 1 and 2) at 40oC for 5 h . Reaction products were analyzed from reaction mixtures before (lane 1) and after reaction (lane 2). Authentic sugar abbreviations are as follows:
Gal, galactose; Glc, glucose.
에의한활성의저해도가 galactose에비해훨씬낮으나[20, 28], B. licheniformis DSM 13의효소경우는 glucose에의 해서도효소활성이크게저해되는것으로보고되었다[12].
한편 xylose에의해서는효소활성이저해를받지않았을뿐
아니라, xylose 농도가 300 mM 이하에서는β-galactosidase 활성이미약하게증가되는현상을보였는데이와같은현상 은 B. licheniformis YB-1105에서생산된효소에서도보고 된바있다.
요 약
가정에서 제조된 된장으로부터 lactose를 glucose와 galactose로가수분해하는균체외β-galactosidase의생산균 이분리되었다. 분리균 YB-1414는형태적특성, 생화학적성 질 및 16S rRNA 유전자 염기서열에 근거하여 Bacillus licheniformis로 확인되었다. 탄소원과 질소원으로 밀기울 (1%)과 yeast extract (2.5%)를사용하였을때 B. licheniformis YB-1414의β-galactosidase 생산성이최대 6.2 U/ml에이르 렀다. 특히 밀기울의 불용성 성분이 수용성 성분보다 β- galactosidase 생산성을더증가시키는것으로확인되었다. β-galactosidase의 para-nitrophenyl-β-D-galactopyranoside 가수분해활성은 pH 6.0과 55-60oC에서가장높았으며, 낮 은농도의 galactose에의해서도크게저해를받았다. 그러 나 glucose에의해서는β-galactosidase의가수분해활성이
약하게저해를받으며 400 mM glucose가존재하여도최대
활성의 85%에해당하는가수분해활성을보였다.
References
1. Batra N, Singh J, Joshi A, Bhatia S. 2011. Applications of β- gal-III isozyme from Bacillus coagulans RCS3, in lactose hydrolysis. Int. J. Biol. Macromol. 49: 879-884.
2. Batra N, Singh J, Banerjee UC, Patnaik PR, Sobti RC. 2002.
Production and characterization of a thermostable β-galacto- sidase from Bacillus coagulans RCS3. Biotechnol. Appl. Bio- chem. 36: 1-6.
3. Chen W, Chen H, Xia Y, Yang J, Zhao J, Tian F, et al. 2009.
Immobilization of recombinant thermostable β-galactosidase from Bacillus stearothermophilus for lactose hydrolysis in milk.
J. Dairy Sci. 92: 491-498.
4. Chen W, Chen H, Xia Y, Zhao J, Tian F, Zhang H. 2008. Pro- duction, purification, and characterization of a potential ther- mostable galactosidase for milk lactose hydrolysis from Bacillus stearothermophilus. J. Dairy Sci. 91: 1751-1758.
5. Dong Y-N, Liu X-M, Chen H-Q, Xia Y, Zhang H-P, Zhang H, et al. 2011. Enhancement of the hydrolysis activity of β-galacto- sidase from Geobacillus stearothermophilus by saturation mutagenesis. J. Dairy Sci. 94: 1176-1184.
6. Guerrero C, Vera C, Illanes A. 2013. Optimisation of synthesis of oligosaccharides derived from lactulose (fructosyl-galacto- oligosaccharides) with β-galactosidases of different origin.
Food Chem. 138: 2225-2232.
7. Hernaiz MJ, Crout DHG. 2000. A highly selective synthesis of N-acetyllactosamine catalyzed by immobilised β-galactosi- dase from Bacillus circulans. J. Mol. Catalysis B. 10: 403-408.
8. Hirata H, Negoro S, Okada H. 1984. Molecular basis of iso- zyme formation of β-galactosidases in Bacillus stearothermo- philus: isolation of two β-galactosidase genes, bgaA and bgaB. J. Bacteriol. 160: 9-14.
9. Hirata H, Negoro S, Okada H. 1985. High production of ther- mostable β-galactosidase of Bacillus stearothermophilus in Bacillus subtilis. Appl. Environ. Microbiol. 49: 1547-1549.
10. Iijima S, Lin KH, Kobayashi T. 1991. Increased production of cloned β-galactosidase in two-stage culture of Bacillus amylo- liquefaciens. J. Ferment. Bioeng. 71: 69-71.
11. Jo HD, Lee HA, Jeong SJ, Kim JH. 2011. Purification and characterization of a major fibrinolytic enzyme from Bacillus amyloliquefaciens MJ5-41 isolated from Meju. J. Microbiol.
Biotechnol. 21: 1166-1173.
12. Juajun O, Nguyen TH, Maischberger T, Iqbal S, Haltrich D, Yamabhai M. 2011. Cloning, purification, and characterization of β-galactosidase from Bacillus licheniformis DSM 13. Appl.
Microbiol. Biotechnol. 89: 645-654.
13. Konsoula Z, Liakopoulou-Kyriakides M. 2007. Co-production of α-amylase and β-galactosidase by Bacillus subtilis in com- plex organic substrates. Bioresour. Technol. 98: 150-157.
14. Levein RE, Mahoney RR. 1981. Purification and characteriza- tion of β-galactosidase from a strain of Bacillus coagulans.
Fig. 5. Effects of sugar on the β-galactosidase activities of the culture filtrate. The relative activity was determined by measuring β-galactosidase activity of the culture filtrate for pNP-βGal (1.0 mM) in the presence of various concentrations of each sugar including glucose (-○-), mannose (-▼-), xylose (-▽-), and galac- tose (-●-), respectively. Each curve represents the average of three independent experiments within standard errors of 2%
between them.
Antonie Van Leeuwenhoek. 47: 53-64.
15. Li Y, Wang H, Lu L, Li Z, Xu X, Xiao M. 2009. Purification and characterization of a novel β-galactosidase with transglyco- sylation activity from Bacillus megaterium 2-37-4-1. Appl. Bio- chem. Biotechnol. 158: 192-199.
16. Li W, Xiang X, Tang S, Hu B, Tian L, Sun Y, et al. 2009. Effec- tive enzymatic synthesis of lactosucrose and its analogues by β-D-galactosidase from Bacillus circulans. J. Agric. Food Chem. 57: 3927-3933.
17. Nam YD, Lee SY, Lim SI. 2012. Microbial community analysis of Korean soybean pastes by next-generation sequencing.
Int. J. Food Microbiol. 155: 36-42.
18. Ozic C, Akcan N. 2010. Molecular characterization of β-galac- tosidase enzyme from Bacillus licheniformis ATCC 12759. J.
Biotechnol. 150S: 537.
19. Phromraksa P, Nagano H, Boonmars T, Kamboonruang C.
2008. Identification of proteolytic bacteria from thai traditional fermented foods and their allergenic reducing potentials. J.
Food Sci. 73: 189-195.
20. Rahim KA, Lee BH. 1991. Specificity, inhibitory studies, and oligosaccharide formation by β-galactosidase from psychro- trophic Bacillus subtilis KL88. J. Dairy Sci. 74: 1773-1778.
21. Sani RK, Chakraborti S, Sobti RC, Patnaik PR, Banerjee UC.
1999. Characterization and some reaction-engineering aspects of thermostable extracellular β-galactosidase from a new Bacillus species. Folia Microbiol (Praha). 44: 367-371.
22. Schmidt M, Stougaard P. 2010. Identification, cloning and
expression of a cold-active β-galactosidase from a novel Arc- tic bacterium, Alkalilactibacillus ikkense. Environ. Technol. 31:
1107-1114.
23. Shaw GC, Kao HS, Chiou CY. 1998. Cloning, expression, and catabolite repression of a gene encoding β-galactosidase of Bacillus megaterium ATCC 14581. J. Bacteriol. 180: 4734- 4738.
24. Shipkowski S, Brenchley JE. 2006. Bioinformatic, genetic, and biochemical evidence that some glycoside hydrolase family 42 β-galactosidases are arabinogalactan type I oligo- mer hydrolases. Appl. Environ. Microbiol. 72: 7730-7738.
25. Spizizen J. 1958. Transformation of biochemically deficient strains of Bacillus subtilis by deoxyribonucleate. Proc. Natl.
Acad. Sci. USA 44: 407-408.
26. Warmerdam A, Paudel E, Jia W, Boom RM, Janssen AE.
2013. Characterization of β-galactosidase isoforms from Bacillus circulans and their contribution to GOS production.
Appl. Biochem. Biotechnol. 170: 340-358.
27. Welsch N, Homuth G, Schweder T. 2012. Suitability of differ- ent β-galactosidases as reporter enzymes in Bacillus subtilis.
Appl. Microbiol. Biotechnol. 93: 381-392.
28. Yoon K-H. 2012. Properties of β-galactosidase from Bacillus licheniformis isolated from cheongkookjang. Korean J. Micro- biol. Biotechnol. 40: 17-22.
29. Yoon K-H, Lim BL. 2007. Cloning and strong expression of a Bacillus subtilis WL-3 mannanase gene in B. subtilis. J. Micro- biol. Biotechnol. 17: 1688-1694.