대한소화기학회지 1999;33:765 - 775
5)
접수: 1999년 12월 10일, 승인: 1999년 2월 20일
연락처: 정현채, 110-744, 서울시 종로구 연건동 28, 서울대학교 의과대학 내과학교실 Tel: (02) 740-8120, Fax: (02) 743-6701
H e lic o b ac t e r p y lo ri 감염 후 C a s p a s e - 3 활성을 통한 인체 위상피세포의 A popt os is
한양대학교 의과대학 미생물학교실 및 의과학연구소, 서울대학교 의과대학 내과학교실 및 간연구소*
김정목・김주성*・정현채*・송인성*・김정룡*
Ca s p a s e -3 Ac t i v a t i o n Le a d s t o Ap o p t o s i s o f H u m a n Ga s t ri c E p i t h e li a l Ce l ls In fe c t e d w i t h H el i c ob a c t er p yl or i
J u n g Mog g Ki m , M.D., J o o S u n g Ki m , M.D.*, H y u n Ch ae J u n g , M.D.*, In S u n g S o n g , M.D.* a n d Ch u n g Yo n g Ki m , M.D.*,
Department of Microbiology and Institute of Biomedical Science, Hanyang University College of Medicine, Seoul; Department of Internal Medicine and Liver Research Institute*,
Seoul National University College of Medicine, Seoul, Korea
Background/Aims: Infection with Helicobacter pylori activates a proinflammatory gene program in
human gastric epithelial cells and is associated with significant epithelial cell damage. We evaluated whether H. pylori infection could increase apoptosis of gastric epithelial cells via caspase-3 activationMethods: After human gastric epithelial cells were infected with H. pylori, apoptosis was assessed
by Hoechst staining, flow cytometric analysis, and cell death detection enzyme linked immunosorben assay. Caspase-3 activation was determined by the detection of the chromophore p-nitroanilide (pNA) after cleavage from the substrate DEVD-pNA. Results: Activation of caspase-3 was first apparent 12 hours and the phenotypic expression of apoptosis was first apparent 18 hours after H. pylori infection The addition of DEVD-fmk inhibited apoptosis of H. pylori-infected epithelial cells. The addition o TNFα significantly increased caspase-3 activation and apoptosis of Hs746T gastric epithelial cells infected with H. pylori. The extent of apoptosis was similar in cases of cagA+cytotoxin+, cagA+cytotoxin- or cagA-cytotoxin- H. pylori-infected gastric epithelial cell cultures. Conclusions: These results suggest that H. pylori can induce gastric epithelial cell apoptosis by activation of caspase-3 Furthermore, this apoptotic process can be induced directly by H. pylori and regulated by immune mediators such as TNFα. (Kor J Gastroenterol 1999;33:765 - 775)
Key Words: Apoptosis, Caspase-3, Gastric epithelial cells, Helicobacter pylori
766 대한소화기학회지 : 제 33 권 제 6 호 1999
서 론
Helicobacter pylori는 만성 위염과 재발하는 소화 성 궤양의 중요한 원인균으로 지목받고 있다.1-3 이 균의 초기 염증반응은 숙주 위상피세포층과의 상호 작용을 통하여 나타난다. 즉, 감염 한 시간 이내에 interleukin (IL)-8과 같은 chemokine 유전자뿐만 아 니라 IL-1, tumor necrosis factor (TNF)α와 같은 일 련의 친염증성 cytokine 유전자 program이 활성화된 다.4,5 또한 상기의 친염증성 cytokine 등에 의하여 위 점막 내로 유입된 호중구에서도 IL-8과 같은 chemo- kine이 생성되어 염증반응을 더욱 촉진시키게 된다.6 따라서 chemokine에 의한 염증세포의 위점막 내 유 입, 위점막의 위축, 위상피세포 증식의 변화 등은 H.
pylori 감염증의 중요한 병리조직 소견이다.1,7,8 위점막 상피세포는 수명이 약 2-6일 정도이며, 위 선의 경부에 존재하는 세포로부터 분화/증식하여 위 점막 표면으로 이동하게 된다. 그리고 생체에서는 위상피세포의 수를 일정하게 유지하기 위하여 apop- tosis (programmed cell death)라는 기전을 통해 위상 피세포의 증식을 조절한다.9-12 이와 같은 apoptosis 는 세균 또는 바이러스에 감염된 세포를 능동적으로 제거한다는 점에서 감염증의 방어기전으로 작용한 다.13-16 그러나 apoptosis가 비정상적으로 증가할 경 우 오히려 과도한 세포의 손실을 초래하여 점막의 파괴를 가져올 수 있다. 이런 점에서 H. pylori의 감 염시 관찰되는 apoptosis17-19는 오히려 병리기전으로 작용할 가능성이 매우 높다. 또한 H. pylori가 위상 피세포에 부착되는 것과 감염된 위상피세포의 apop- tosis를 class II MHC를 인지할 수 있는 항체에 의해 억제시킬 수 있다는 보고20는, H. pylori 부착과 apop tosis가 밀접한 연관성이 있음을 시사해 줌과 동시에 H. pylori가 직접적으로 위상피세포의 apoptosis를 유도할 수 있다는 사실을 제시해 준다. 그러나 아직 까지 H. pylori의 감염이 어떤 경로를 통해 apoptosis 로 유도되는지에 대해서는 알려져 있지 않은 점이 많다.
현재까지 잘 알려져 있는 apoptosis의 경로로는 TNF 수용체를 통한 것과 Fas 수용체를 통한 것을
들 수 있다.12,21 이 수용체에 TNFα 또는 Fas ligand 가 결합될 경우 caspase 경로를 통해 apoptosis가 유 도된다. 이 때, caspase-3 (apopain, CPP-32, YAMA) 은 이와 같은 경로에서 중심적인 역할을 담당한다.
즉, caspase-3은 세포질에 존재하는 DNase를 활성화 시켜 핵 내로 이동시킴으로써 염색체 DNA의 분절 화를 초래케 한다.22 본 연구자들은 이미 H. pylori에 감염된 위상피세포에서 TNFα가 발현될 수 있음을 보고한 바 있다.4 따라서 이와 같이 발현된 TNFα는 TNF 수용체를 통하여 위상피세포의 apoptosis를 매 개할 수 있다. 만일 이와 같은 경로로 apoptosis가 유 도된다면 결국 caspase-3의 활성으로 이어질 가능성 이 매우 높다. 그러나 현재까지 H. pylori에 감염된 위상피세포에서의 caspase-3 활성에 대해서는 보고 된 바가 전혀 없다. 상기의 가능성을 평가하기 위하 여 본 연구에서는 먼저 H. pylori에 감염된 위상피세 포의 apoptosis 측정을 확립한 뒤, 이같은 apoptosis 가 caspase-3의 활성을 통하여 유도되는지를 검사하 였다.
대상 및 방법
1. 세균 배양
본 연구에 사용된 H. pylori 균주는 기존의 방법을 이용하여 분리 배양하였다.4,5,23 계대배양 횟수가 10 회 미만의 균을 이용하기 위하여 H. pylori 균주들은 15% glycerol이 포함된 Brucella broth에 넣어 -70℃
에 보관하였고, 실험 직전 녹인 뒤 증식시켜 사용하 였다. H. pylori 균주의 독성인자인 cagA 및 공포성 세포독소(vacuolating cytotoxin)검사는 기존의 방법 대로 시행하였다.23
2. 위상피세포주 배양 및 감염
실험에 이용한 인체 위상피세포주로는 AGS (AT- CC CRL 1739), Hs746T (ATCC HTB 135), KATO- III (ATCC HTB 103)과 SNU-5 (KCLB 0005)였다.
이들 세포들은 Dulbecco' s modified Eagle' s medium (DMEM, pH 7.4, Sigma, St. Louis, Missouri, USA), RPMI-1640 (Sigma) 또는 Ham' s F12 (Sigma)에 10%
우태아 혈청(Hyclone Laboratories, Logan, Utah,
김정목 외 4인. H. pylori 감염에 의한 Apoptosis 767
USA)과 항생제(100 unit/ml의 penicillin, 100 μg/ml 의 streptomycin)를 사용하여 37℃ 5% CO2 배양기에 서 배양하였다.
세균 감염은 항생제가 제거된 상태에서 각 세포 를 6-well tissue culture plate에 24-48시간 동안 배양 한 다음 균수: 위상피세포의 비율이 250:1이 되게 균 을 첨가하였다. 균수 측정은 McFarland scale 또는 hemocytometer를 이용하였다. 경우에 따라서는 H.
pylori 감염과 동시에 인체 재조합 TNFα(R&D Sys tems, Minneapolis, Minnesota, USA) 또는 goat anti- human TNFα 단세포군 항체(R&D Systems)를 첨가 한 뒤 일정 시간 배양 후 apoptosis를 측정하였다.
3. 위상피세포의 a popt os is 측정 1) A popt os is 의 형태학적 검사
Apoptosis가 일어난 세포는 정상적인 세포와는 다 른 형태학적 특징을 나타낸다.21 이를 검사하기 위하 여 위상피세포를 phosphate-buffered saline (PBS, pH 7.4)로 1회 세척한 뒤 기존의 방법15대로 Hoechst 33258 [5 μg/ml; 2' -(4-hydroxyphenyl)-5-(4-methyl- piperazinyl)-2,5-bi-1H-benzimidazole; CalBiochem, La Jolla, California, USA]을 이용하여 37℃에서 10분 동안 염색한 뒤, epifluorescence microscopy (Axio- phot, Carl Zeiss, Oberkochen, Germany)로 apoptotic body 또는 DNA 분절화를 관찰하였다. 한편 부유세 포들은 3% paraformaldehyede로 고정시킨 뒤, Hoe- chst 33258 (5 μg/ml)로 37℃에서 10분 동안 염색한 다음 500 rpm으로 5분 동안 cytospin을 시행하였다.
그 뒤, 앞서와 같이 epifluorescence microscopy로 apoptosis가 일어난 세포를 관찰하였다.
2) A popt os is 의 생화학적 검사
형태학적 검사로는 객관적인 apoptosis의 정량이 불가능하므로 생화학적 검사를 이용하여 apoptosis 의 증감 정도를 검사하였다.
(1) Annexin V와 propidium iodide를 이용한 검사 Apoptosis의 초기에는 원래 세포막 내부에 위치하 는 phosphatidylserine이 세포 표면으로 노출하게 된 다. 이와 같이 노출된 phosphatidylserine은 annexin
V와 쉽게 결합된다.24,25 이를 이용하여 햇빛이 차단 된 상태에서 FITC-conjugated annexin V (R&D Sys- tems)로 위상피세포를 실온에서 15분 동안 염색하 였다. 그 뒤, 동일한 조건에서 다시 propidium iodide (R&D Systems)로 이 세포를 15분 동안 더 염색한 다음 1시간 이내에 flow cytometry로 염색된 세포를 관찰하였다. 이와 같이 이중 염색(double stain)을 하 게 되면 apoptosis가 진행 중인 세포는 annexin V로 만 염색되고 propidium iodide로는 염색되지 않는 반면, necrosis가 진행 중인 세포 또는 late stage에 있는 apoptosis 세포들은 annexin V와 propidium io- dide로 동시에 염색된다. 이 결과는 Fig. 1에 제시되 어 있다.
(2) DiOC6(3)을 이용한 검사
Apoptosis가 일어난 세포는 mitochondria로부터 cytochrome c의 분비에 이어 mitochondrial trans- membrane potential (Δψm)이 감소된다.26 이를 이용 하여 Δψm에 의하여 세포 내로 유입될 수 있는 DiOC6(3) (3' 3-dihexyloxacarbocyanine iodide; Mole- cular Probes, Eugene, Oregon, USA) (40 nM)로 위상 피세포를 37℃에서 15분 동안 염색한 뒤, flow cytometry (Becton Dickinson, Mountain View, Cali- fornia, USA)로 Δψm이 감소되어 있는 apoptosis 세 포들을 관찰하였다.15
(3) 세포질 내로 유리된 nucleosome검사
Apoptosis가 진행 중인 세포의 염색체는 DNase에 의해 분절화가 일어나 결국 oligonucleosome 형태로 세포질 내로 유출된다.21 이와 같은 oligonucleosome 은 histone과 DNA로 구성되어 있다. 이를 이용하여 H. pylori를 감염시킨 위상피세포(1×105)의 lysate를 만든 뒤, 미리 streptavidin이 coating되어 있는 well에 가하였다. 그 후 biotin이 결합된 마우스 항- histone 단세포군 항체를 첨가하여 실온에서 1시간 동안 방 치한 다음, peroxidase가 결합된 마우스 항- DNA 단 세포군 항체를 1시간 동안 가하였다. 최종적으로 2,2' -azino-di(3-ethylbenzthiazolin-sulfonat)로 10분 동 안 발색시킨 다음, 405 nm에서 흡광도를 측정하였 다. 이 과정에서 사용한 시약들은 cell death detec-
768 The Korean Journal of Gastroenterology : Vol. 33, No. 6, 1999
tion ELISAplus kit (Boehringer Mannheim GmbH, Ger- many)를 이용하였다. 한편 세포질 내로 유출된 oli- gonucleosome의 상대적 증가율은 제조회사의 설명 서대로 감염시키지 않은 대조군에 대한 감염군에서 나타나는 상대적인 흡광도의 비율(enrichment factor) 로 표기하였다.
4. H . p y lori에 감염시킨 위상피세포에서의 ca s pa s e - 3 활성도 측정
Caspase-3의 활성도는 p-nitroanilide (pNA)로 la- bel된 Asp-Glu-Val-Asp (DEVD)의 분해 정도로 측정 하였다. 즉, trypsin 처리하여 떨어뜨린 위상피세포를 PBS로 1회 세척한 다음 3×106개의 세포에 lysate buffer [10 mM Tris-HCl (pH 7.5), 10 mM NaH2PO4/
NaHPO4 (pH 7.5), 130 mM NaCl, 1% Triton-X100, 10 mM NaPPi]를 첨가하였다. 그 뒤 얼음 속에서 20 분 동안 방치한 뒤, 4℃에서 12,000 rpm으로 3분 원 침하여 상층액만을 회수하였다. 이 상층액에 1 mM HEPES buffer와 함께 50 μM의 DEVD-pNA (Clon- tech, Palo Alto, California, USA)를 첨가한 뒤, 37℃
에서 1시간 방치하였다. 그 후 ELISA reader를 이용 하여 405 nm에서 흡광도를 측정하여 대조군과 비교 한 상대적 수치로 caspase-3의 활성도를 평가하였다.
양성 대조군으로는 인체 T세포 림프종 세포주인 Jurkat 세포에 항-Fas 단세포군 항체(clone CH-11, IgM, Upstate Biotechnology, Lake Placid, New York, USA) (50 ng/ml)을 8시간 동안 가하여 얻은 세포 파 쇄액을 이용하였다. 한편 각 실험마다 DEVD-pNA 의 첨가에 앞서서 caspase-3 억제제인 Asp-Glu-Val- Asp-CH2F (DEVD-fmk; Clontech)를 사용하여 증가 된 caspase-3 활성도가 DEVD-fmk (50 μM)에 의하 여 억제되는지를 확인하여 활성도 증가가 특이적인 것인지를 평가하였다.
결 과
1. H . p y lori 감염에 의한 인체 위상피세포주의 a popt os is 평가
현재까지 H. pylori 감염에 의한 apoptosis의 시험 관적 관찰은 대부분 한 가지 방법으로 측정하였을
뿐, 다양한 방법으로 그 상대적인 비교가 이루어진 것은 없었다. 이와 같은 단점을 보완하고 H. pylori 감염에 의한 apoptosis의 기초 자료를 확립하고자 본 연구에서는 4가지의 다양한 방법을 이용하여 apo- ptosis를 평가하였다.
Hoechst 염색을 이용하여 apoptosis의 형태학적 특징을 관찰한 결과 대조군의 경우 1.2±0.2%, H.
pylori 감염군의 경우 2.3±0.5%의 세포에서 apop- tosis를 나타내었다(Table 1). 그런데 이와 같은 현미 경적 관찰은 주관적인 면이 많이 작용하므로 apop- tosis의 정도를 보다 객관적으로 정량하기 위하여 각 종 생화학적 검사법을 이용하였다. 즉, annexin V와 propidium iodide로 염색한 Hs746T 세포에서, an- nexin V로만 염색된 apoptosis가 진행 중인 세포는 (Fig. 1 A, B의 R1 지역) 대조군의 경우 1.5±0.7%인 데 비하여, H. pylori를 감염시킨 세포에서는 4.2±
0.3%로 대조군에 비하여 2.8배의 증가를 보여주고 있었다(Table 1). 한편 propidium iodide로는 염색되 지 않은 반면 annexin V와 propidium iodide로 동시 에 염색된 necrosis가 진행 중이거나 또는 late stage 에 있는 apoptosis 세포들은(Fig. 1 A, B의 R2 지역) 대조군에서 평균 0.1%, H. pylori를 감염시킨 군에서 는 평균 0.8%를 나타내고 있었다. 또한 mitochon- drial transmembrane potential이 감소한 apoptosis 세 포들은(Fig. 1 C, D의 M1 지역) 대조군의 경우 6.8±
0.5%, H. pylori 감염군의 경우 14.3±1.1%를 나타내 었다(Table 1). 그리고 cell death detection ELISA를 이용한 측정 결과 H. pylori 감염군에서의 oligonu- cleosome 유출(DNA 분절화) 정도는 대조군에 비하 여 평균 3.3배의 증가를 나타내었다(Table 1). 이와 같이 4가지의 서로 다른 방법을 이용한 apoptosis의 측정 결과 그 절대적인 수치는 차이를 나타내고 있 지만, 증가율은 거의 비슷한 경향을 보여주고 있었 다. 따라서 이후 실험에서는 cell death detection ELISA를 이용하여 apoptosis를 정량화하였다.
앞서의 결과는 Hs746T 세포를 이용한 결과였다.
그래서 다른 인체 위상피세포주도 동일한 결과를 나 타내는지를 확인하기 위하여 AGS, KATO-III 및 SNU-5 세포주에 H. pylori를 48시간 동안 감염시킨 뒤 cell death detection ELISA로 apoptosis의 정도를
Kim et al. Apoptosis by H. pylori Infection 769
측정하였다. 그 결과 모든 세포에서 대조군에 비하 여 평균 2.4-4.2배의 apoptosis가 증가되는 것을 관찰 할 수 있었다(Fig. 2).
2. 각종 병독인자를 지닌 H . py lori 균주 감염에 의한 인체 위상피세포주 a poptos is 의 시간 대별 변화
CagA 또는 세포독소와 같은 병독인자들은 위장 질환의 정도와 관련이 높다는 보고들이 제시되고 있 다.27-29 그래서 이들 병독인자들의 보유 유무에 따른 apoptosis를 시간대별로 관찰하였다. 그 결과 cagA 또는 세포독소의 보유 유무에 관계없이 모든 H.
pylori 균주는 거의 동일한 수준의 apoptosis를 나타 내었고, 친염증성 cytokine 발현에 비해 상대적으로 늦게 유도되고 있었다(Fig. 3). 즉, H. pylori 감염 12 시간까지는 apoptosis가 나타나지 않고 있다가 18시 간이 되어서야 비로소 apoptosis가 관찰되기 시작하 여 48시간까지 지속되었다.
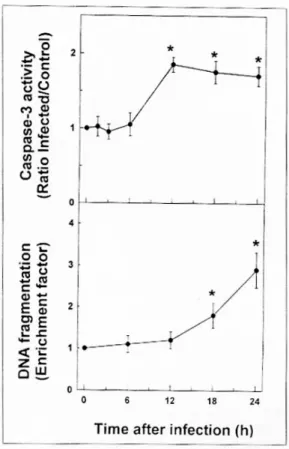
3. H . p y lori 감염에 의한 인체 위상피세포주의 ca s pa s e - 3 활성
위상피세포에서의 caspase-3 활성은 H. pylori 감 염 12시간에 대조군에 비해 뚜렷한 증가를 관찰할 수 있었고, 24시간 내내 대조군에 비해 약 1.8배 이 상 증가를 나타내고 있었다(Fig. 4). 이와 같은 실험 system에서의 실제적인 apoptosis는 caspase-3이 활 성화된 후 6시간이 경과한 감염 18시간부터 나타나 기 시작하였다. 한편 세포 lysate에 50 μM의 DEVD- fmk를 첨가했을 경우 caspase-3의 활성은 완전히 차 단되었다. 또한 H. pylori를 감염시킨 Hs746T 세포 에 200 μM의 DEVD-fmk를 첨가한 뒤, 24시간 후 cell death detection ELISA로 apoptosis를 측정한 결 과 H. pylori를 감염시킨 군에서의 DNA 분절화는 대조군에 비하여 평균 2.3배 증가한 데 비하여 H.
pylori와 DEVD-fmk를 동시에 처리한 군에서는 평균 1.2배를 나타내었다. 이와 같은 결과는 H. pylori 감 염증에 의한 위상피세포의 apoptosis는 caspase-3의 활성이 선행됨과 동시에 caspase 경로를 통해 apop- tosis가 유도되고 있음을 시사해 준다.
770 대한소화기학회지 : 제 33 권 제 6 호 1999
4. H . p y lori 감염에 의한 인체 위상피세포주 a popt os is 에 있어서의 T N F α의 역할 H. pylori에 감염된 위상피세포에서는 세포 RNA 1 μg 당 약 2×106 정도의 TNFα mRNA가 발현된 다.4 따라서 이와 같은 TNFα가 위상피세포의 apop- tosis에 미치는 영향을 알아보기 위하여 H. pylori를 감염시킨 Hs746T 세포에 20 ng/ml의 TNFα를 첨가
한 뒤, 24시간 후 cell death detection ELISA로 apoptosis를 측정하였다. 그 결과 H. pylori만을 감염 시킨 군(enrichment factor, 2.77±0.62)에 비하여 H.
pylori를 감염시킴과 동시에 TNFα를 첨가한 군 (enrichment factor, 4.47±0.34)에서 평균 61.4%의 DNA fragmentation의 증가가 관찰되었다(Fig. 5). 그 리고 이 실험 system에 항-인체 TNFα 단세포군 항 체(20 μg/ml)를 가하여 TNFα를 중화시켰을 때의 Fig. 1. Flow cytometric analysis of apoptosis of gastric epithelial cells infected with H.
pylori. Confluent monolayers of Hs746T cells in 6-well plates were left uninfected as a control (A), or were infected with cagA+cytotoxin+ H. pylori (B) for 48 hours.
Adherent and nonadherent cells were pooled, incubated with FITC-conjugated annexin V and propidium iodide, and analyzed by flow cytometry. Data are from a representative experiment. Early apoptotic cells have increased annexin, but no propidium iodide staining as indicated by the area marked R1. Necrotic and late apoptotic cells were stained with both annexin V and propidium iodide as indicated by the area marked R2. Quantitative data for R1 obtained in several independent experiments are presented in Table 1. (C and D) Culture and infection conditions are the same as in A and B. Panel C shows uninfected cells as a control and panel D shows infected cells. Adherent and nonadherent cells were incubated with 40 nM DiOC6(3) for 15 minutes at 37℃ and analyzed by flow cytometry. The area marked as M1 to the left of the major peak contains the apoptotic cell population.
김정목 외 4인. H. pylori 감염에 의한 Apoptosis 771
apoptosis는 (enrichment factor, 2.80±0.71) H. pylori 만을 감염시킨 군에서와 거의 비슷한 수준을 유지하 고 있다. 한편 H. pylori를 감염시킨 Hs746T 세포에 TNFα (20 ng/ml)을 가한 뒤, 12시간 후에 caspase-3 의 활성도를 측정한 결과 H. pylori만을 감염시킨 군 에 비하여 약 38%의 증가를 나타내었다.
Fig. 2. Apoptosis of human gastric epithelial cell lines in response to infection with H. pylori. Confluent monolayers of various gastric epithelial cells in 6-well plates were infected with cagA+cytotoxin+ H. pylori for 48 hours. Cells were assessed for apoptosis using the cell death detection ELISA. Numbers refer to DNA fragmentation as measured by the enrichment factor. Values are means±SEM of three separate experiments.
Fig. 3. Time course of apoptosis in Hs746T cells after infection with H. pylori. Confluent monolayers of Hs746T cells in 6-well plates were infected with cagA+cytotoxin+
( ), cagA+cytotoxin- ( ), and cagA-cytotoxin- (o) H.
pylori strains. Apoptosis was assessed using the cell death detection ELISA at the indicated times after infection.
Numbers refer to DNA fragmentation as measured by the enrichment factor. Values are means of three or more separate experiments.
Fig. 4. Time course of caspase-3 activation following H.
pylori infection of Hs746T cells. (A) Confluent mono- layers of Hs746T cells in 6-well plates were infected with cagA+cytotoxin+ H. pylori and cells were assayed for caspase-3 activity at the indicated times after infection.
Values are means±SEM of 3 separate experiments. As a comparison, caspase-3 activation was increased 3.8 fold in Jurkat cells treated with anti-Fas monoclonal antibody (clone CH-11, 50 ng/ml) for eight hours. Asterisks indi- cate values significantly different from H. pylori-infected cells at 0 hour postinfection (p<0.05). (B) Parallel cultures were assessed for apoptosis using the cell death detection ELISA at the indicated times after infection. Numbers refer to DNA fragmentation as measured by the enrich- ment factor. Values are means±SEM of three separate experiments.
772 The Korean Journal of Gastroenterology : Vol. 33, No. 6, 1999
고 찰
본 연구에서 H. pylori는 직접 위상피세포의 apo- ptosis를 유도할 수 있음을 제시하였다. 즉, 감염 12 시간 후에 caspase-3이 활성화되고 난 뒤에 실제적인 위상피세포의 apoptosis가 유도되었다. 특히 H.
pylori가 감염된 위상피세포 apoptosis의 시작은 친염 증성 cytokine 유전자의 발현이 H. pylori 감염 초기 인 약 1-3시간 이내에 위상피세포부터 발현된다는 점4,5에 비하여 상대적으로 늦게 나타나고 있었다. 이 와 같이 상대적인 발현의 지연은 부분적으로는 H.
pylori 감염에 의한 NF-kB의 활성30에 의해 초래될 가능성이 있다. 이와 같은 추정의 근거는 TNFα로 자극을 가한 세포에서의 apoptosis는 NF-kB의 활성
을 억제시켜 나타난다는 보고31,32와 Rickettsia ric- kettsii에 감염된 내피세포의 apoptosis는 NF-kB의 활 성에 의하여 억제된다는 보고들에 의해 가능하다.33 이런 의미에서 NF-kB의 활성은 세포의 증식을 조장 하고 세포 사멸의 signal을 억제하는 기능을 행할 가 능성도 제시된다.12,21 또한 H. pylori 감염 초기에 apoptosis가 억제된다는 점은 위상피세포로부터 각 종 친염증성 매개체를 생성시켜 위점막조직에서 염 증반응이 유발되게끔 하는 데 필요한 시간을 제공한 다는 점에서 H. pylori에 의한 병리기전의 중요한 단 계일 것으로 추정된다.
H. pylori에 감염된 위상피세포에서 caspase-3의 활성이 증가되었다는 결과와 caspase-3 억제제에 의 하여 위상피세포 apoptosis가 감소되었다는 결과들 은 H. pylori에 의한 apoptosis 유도 경로의 일부를 밝혔다는 점에서 중요하다. Caspase-3의 활성화는 여러 경로를 통해 이루어진다. 대표적으로 TNF 수 용체를 통한 것과 Fas 수용체를 통한 것을 들 수 있 다. 즉, TNF 수용체에 TNFα가 결합될 경우 signal이 TRADD (TNF receptor-associated death domain), FADD (Fas-associating protein with a death domain) 를 통해 caspase-8을 활성화시키고 이것은 결국 caspase-3을 활성화시킨다. 또한 Fas ligand가 Fas 수 용체에 결합될 경우에도 이 signal이 FADD를 거쳐 caspase-8을 활성화시키고 이어서 앞서와 같이 cas- pase-3이 활성화된다. 결국 caspase-3은 apoptosis의 signal이 전달되는 경로에서 중심적인 역할을 담당 하고 있다.12,21 한편 활성화된 caspase-3은 세포질에 존재하는 DNase를 활성화시켜 핵 내로 이동시킴으 로써 염색체 DNA의 분절화를 초래케 하여 비로소 apoptosis를 유도하게 된다.22 그런데 H. pylori에 의 한 위상피세포의 apoptosis는 Bak-의존성 경로에 의 해 유도될 수 있다는 보고가 있다.34 Bcl-2에 속한 것 들 중 Bcl-2와 Bcl-XL은 apoptosis를 억제하지만, Bax 등은 apoptosis를 촉진하는 역할을 한다.21 최근 TNFα와 같은 cytokine이 Bax/Bcl-X의 비율을 변화 시키고 이어서 caspase-3의 활성을 조절할 수 있다는 보고35로 미루어 H. pylori 감염에 의한 Bax-의존성 경로는 결국 caspase-3의 활성을 통해 그 효과가 나 타날 가능성이 있다. 그러나 아직까지 위상피세포에 Fig. 5. Exogenous TNFα increased apoptosis in H. pylori-
infected gastric epithelial cells. Confluent monolayers of Hs746T cells in 6-well plates were incubated for 24 hours with TNFα (20 ng/ml), cagA+cytotoxin+ H. pylori, or anti-TNFα monoclonal antibody (20 μg/ml), alone or in combinations as indicated in the figure. Cells were asses- sed for apoptosis using the cell death detection ELISA.
Numbers refer to DNA fragmentation as measured by the enrichment factor. Values are means±SEM of three sepa- rate experiments. An asterisk indicates a value signifi- cantly different from H. pylori-infected cells without added anti-TNFα antibody (p<0.05).
Kim et al. Apoptosis by H. pylori Infection 773
서 상기의 경로가 확인된 바 없으나 본 연구는 이와 같은 추론의 한 단서를 제공해 주고 있는 것으로 사 료된다. 한편 본 연구자들은 이미 H. pylori에 감염 된 위상피세포 RNA 1 μg 당 약 2×106 정도의 TNF αmRNA가 발현된다고 보고한 바 있다.4 또한 본 연 구에서 위상피세포에서 발현되는 TNFα는 caspase-3 의 활성을 증가시키고 궁극적으로는 위상피세포의 apoptosis를 유도할 수 있음을 제시하였다. 이와 같 은 결과들은 H. pylori는 직접적으로 위상피세포의 apoptosis를 유도할 뿐만 아니라 위상피세포로부터 매개물질을 생성시켜 간접적으로 앞서 기술한 경로 를 통해 apoptosis를 유도할 가능성을 시사하고 있 다. 최근 침습성 세균에 감염된 장상피세포에서는 inducible nitric oxide synthase (iNOS)가 발현되고,36 이 효소에 의해 생성되는 nitric oxide (NO)가 장상 피 세포의 apoptosis를 유도할 수 있음이 발표되었 다.15 그런데 H. pylori의 감염증에서 NO가 증가된다 는 보고37를 참고로 해볼 때, NO 또한 간접적으로 위상피세포의 apoptosis를 매개할 가능성이 높다. 따 라서 본 연구진에서는 이에 대한 연구를 진행 중에 있으며 동시에 H. pylori 감염에 의한 Fas 수용체의 발현과 이를 통한 apoptosis 유도에 관해서도 연구를 진행 중에 있다.
외국의 여러 연구에서 H. pylori의 cagA 또는 세 포독소의 존재와 위장질환의 심각도가 비례한다는 보고들이 많이 발표되고 있다.27-29 그러나 우리 나라 에서의 위장질환과 상기의 병독인자와는 별 상관관 계가 없다는 보고도 제시되고 있다.23 이와 같은 차 이점을 규명하고자 본 연구에서는 우리 나라에서 분 리한 균의 병독인자를 검사한 뒤 병독인자에 따른 apoptosis 유도능의 차이를 검사하였으나 cagA 또는 세포독소와 같은 독성인자의 유무와 위상피세포 apoptosis 사이에는 별다른 차이를 발견하지 못했다.
이와 같은 결과는 외국의 다른 연구자들의 보고에 의해 뒷받침되고 있다.17,34 동시에 앞서 본 연구자들 이 보고한 H. pylori의 병독인자와 위상피세포의 친 염증성 cytokine 유도능 사이에는 상관관계가 없다 는 성적38과도 일치한다. 이런 여러 결과들을 토대로 미루어 볼 때 외국에서 강조되고 있는 H. pylori 균 의 cagA 또는 세포독소인자 보유 유무는 우리 나라
현실에서는 실질적인 병독인자로 작용할 가능성은 낮은 것으로 추정된다. 따라서 새로운 병독인자 규 명의 필요성이 제기된다.
Apoptosis는 위조직의 세포수를 조절하는 중요한 과정이다.9 이 과정은 위점막 표면에서 주로 발생하 며 경우에 따라서는 위선의 하부로 이어지기도 한 다. 그런데 위선 하부의 과도한 apoptosis는 궁극적 으로 위점막상피세포의 간세포(stem cell)의 파괴로 이어지기 때문에 위축성 위염 또는 장상피화생으로 발전할 가능성이 있다. 이같은 관점을 고려해 볼 때, H. pylori 감염에 의해 초래되는 과도한 위점막 상피 세포의 apoptosis는 결국 급성기에는 위염 또는 궤양 을 유발할 수 있을 뿐만 아니라 만성적으로는 위축 성 위염과 같은 질환을 초래할 것으로 추정된다.
요 약
목적: H. pylori의 감염은 위점막상피세포로부터 각종 친염증성 cytokine의 유도뿐만 아니라 궁극적 으로는 상피세포층의 파괴로 이어진다. 본 연구에서 는 H. pylori의 감염이 caspase-3의 활성을 통해 apoptosis가 유도될 수 있는지의 여부를 검사하였다.
대상 및 방법: 인체 위상피세포에 H. pylori를 감염 시킨 뒤, Hoechst 염색, flow cytometry 분석 및 cell death detection ELISA를 이용하여 apoptosis를 측정 하였다. 또한 caspase-3의 활성은 첨가한 기질 DEVD-pNA로부터 분해되는 pNA를 측정하여 결정 하였다. 결과: H. pylori에 감염된 위상피세포에서는 감염 12시간 후에 caspase-3이 활성화되었고 이어서 감염 18시간이 지나서야 비로소 apoptosis가 유도되 었다. 또한 caspase-3 억제제를 투여했을 때 apo- ptosis가 감소하였다. H. pylori에 감염된 Hs746T 세 포주에 TNFα를 첨가했을 경우 apoptosis와 cas- pase-3의 활성이 유의하게 증가하였다. H. pylori의 cagA 또는 세포독소와 같은 독성인자의 유무와 위 상피세포 apoptosis 사이에는 별다른 차이를 관찰하 지 못했다. 결론: 본 연구 결과들은 H. pylori가 감염 된 후 위점막상피 세포에서는 caspase-3이 활성화된 후 이어서 apoptosis가 유도된다는 점을 제시해 준 다. 아울러 이와 같은 apoptosis는 균 자체에 의해 유
774 대한소화기학회지 : 제 33 권 제 6 호 1999
도될 뿐만 아니라 TNFα와 같은 면역매개체에 의해 서도 조절될 수 있음을 시사해 준다.
색인단어: 아포토시스, 카스파제-3, 위상피세포, 헤 리코박터 파이로리
참 고 문 헌
1. Marshall BJ, Warren JR. Unidentifed curved bacill in the stomach of patients with gastritis and peptic ulceration. Lancet 1984;1:1311-1315.
2. Rauws EA, Langerberg W, Houthoff HJ, Zanen HC Tytgat GN. Campylobacter pyloridis-associated chron- ic active antral gastritis. A prospective study of it prevalence and the effect of antibacterial and antiul cer treatment. Gastroenterology 1988;94:33-40.
3. NIH Consensus Development Panel on Helicobacter pylori in peptic ulcer disease. JAMA 1994;272:65- 69.
4. Jung HC, Kim JM, Song IS, Kim CY. Helicobacte pylori induces an array of proinflammatory cyto kines in human gastric epithelial cells: quantification of mRNA for interleukin-8, -1α/β, granulocyte- macrophage colony-stimulating factor, monocyte chemoattractant protein-1 and tumor necrosis factor- α. J Gastroenterol Hepatol 1997;12:473-480.
5. Jung HC, Kim JM, Song IS, Kim CY. Increased motility of Helicobacter pylori by methylcellulose could upregulate the expression of proinflammatory cytokines in human gastric epithelial cells. Scand J Clin Lab Invest 1997;57:263-270.
6. Kim JS, Jung HC, Kim JM, Song IS, Kim CY Interleukin-8 expression by human neutrophils acti vated by Helicobacter pylori soluble proteins. Scand J Gastroenterol 1998:33;1153-1159.
7. Robert ME, Weinstein WM. Helicobacter pylori- associated gastric pathology. Gastroenterol Clin North Am 1993;22:59-72.
8. Fan XG, Kelleher D, Fan XJ, Xia HX, Keeling PW Helicobacter pylori increases proliferation of gastric epithelial cells. Gut 1996;38:19-22.
9. Crawford JM. The gastrointestinal tract; stomach. In
Cotrans RS, Qumar V, Robbins SL, eds. Pathologic basis of disease. 5th ed. Philadelphia: WB Saunders 1994:767-783.
10. Bursch W, Oberhammer F, Schulte-Hermann R. Cel death by apoptosis and its protective role agains disease. Trends Pharmacol Sci 1992;13:245-251.
11. Evans VG. Multiple pathways to apoptosis. Cel Biol Int 1993;17:461-475.
12. Nagata S. Apoptosis by death factor. Cell 1997;88 355-365.
13. White E, Sabbatini R, Debbas M, Wold WS, Kushe DI, Gooding LR. The 19 kD adenovirus E1B trans forming protein inhibits programmed cell death and prevent cytolysis by TNF. Mol Cell Biol 1992;12 2570-2580.
14. Robert JM, Searle JW, Cooksley WG. Histologica patterns of prolonged hepatitis C infection. Gastro enterology 1993;26:37-41.
15. Kim JM, Eckmann L, Savidge TC, Lowe DC Witthoft T, Kagnoff MF. Apoptosis of human intes tinal epithelial cells after bacterial invasion. J Clin Invest 1998;102:1815-1823.
16. Zychlinsk A, Sansonetti P. Apoptosis in bacteria pathogenesis. J Clin Invest 1997;100:493-495.
17. Wagner S, Beil W, Westermann J, et al. Regulation of gastric epithelial cell growth by Helicobacter pylori: evidence for major role of apoptosis. Gastro enterology 1997;113:1836-1847.
18. Moss SF, Calam J, Angarwal B, Wang S, Holt PR Induction of gastric epithelial apoptosis by Helico bacter pylori. Gut 1996;38:498-451.
19. Rudi J, Kuck D, Strand S, et al. Involvement of th CD95 (APO-1/Fas) receptor and ligand system in Helicobacter pylori-induced gastric epithelial apop tosis. J Clin Invest 1998;102:1506-1514.
20. Fan X, Crowe SE, Behar S, et al. The effect of clas II major histocompatibility complex expression on adherence of Helicobacter pylori and induction of apoptosis in gastric epithelial cells: a mechanism fo T helper cell type 1-mediated damage. J Exp Med 1998;187:1659-1669.
21. Granville DJ, Carthy CM, Hunt DWS, McManus
김정목 외 4인. H. pylori 감염에 의한 Apoptosis 775
BM. Apoptosis: molecular aspects of cell death and diseases. Lab Invest 1998;78:893-913.
22. Enari M, Sakahira H, Yokoyama H, Okawa K Iwamatsu A, Nagata S. A caspase-activated DNase that degrades DNA during apoptosis, and its inhibi tor ICAD. Nature 1998;391:43-50.
23. 김정목, 정현채, 이주영 등. 한국에서 분리된 Helico bacter pylori 균주에서의 병리인자 분포 및 이에 대한 숙주 방어기전의 분자생물학적 연구. 대한내과학회지 1998;54:40-51.
24. Martin SJ, Reutelingsperger CP, McGahon AJ, et al Early redistribution of plasma membrane phospha tidylserine is a general feature of apoptosis regar dless of the initiating stimulus: inhibition by over expression on Bcl-2 and Abl. J Exp Med 1995;182 1545-1556.
25. Koopman G, Reutelingsperger CP, Kuijten GA Keehnen RM, Pals ST, van Oers MH. Annexin V for flow cytometric detection of phosphatidylserine expression on B cells undergoing apoptosis. Blood 1994;84:1415-1420.
26. Kluck RM, Martin SJ, Hoffman BM, Zhou JS Green DR, Newmeyer DD. Cytochrome c activation of CPP32-like proteolysys plays a critical role in a Xenopus cell-free apoptosis system. EMBO J 1997 16:4639-4649.
27. Blaser MJ, Perez-Perez GI, Kleanthous H, et al Infection with Helicobacter pylori strains possessing cagA is associated with an increased risk of develo ping adenocarcinoma of the stomach. Cancer Res 1995;55:2111-2115.
28. Queiroz DM, Mendes EN, Rocha GA, et al. CagA positive Helicobacter pylori and risk for developing gastric carcinoma in Brazil. Int J Cancer 1998;78 135-139.
29. de Figueiredo Soares T, de Magalhaes Queiroz DM Mendes EN, et al. The interrelationship between
Helicobacter pylori vacuolating cytotoxin and gastric carcinoma. Am J Gastroenterol 1998;93:1841-1847.
30. Keates S, Hitti YS, Upton M, Kelly CP. Helico bacter pylori infection activates NF-kB in gastric epithelial cells. Gastroenterology 1997;113:1099- 1109.
31. Van Antwerp DJ, Martin SJ, Kafri T, Green DR Verma IM. Suppression of TNFα-induced apoptosis by NF-kB. Science 1996;274:787-789.
32. Beg AA, Baltimore D. An essential role for NF-kB in preventing TNFα-induced cell death. Science 1996;274:782-784.
33. Clifton DR, Gross RA, Sahni SK, et al. NF-kB dependent inhibition of apoptosis is essential for host cell survival during Rickettsia rickettsii infec tion. Proc Natl Acad Sci USA 1998;95:4646-4651.
34. Chen G, Sordillo EM, Ramey WG, et al. Apoptosi in gastric epithelial cells is induced by Helicobacte pylori and accompanied by increased expression of BAK. Biochem Biophys Res Commun 1997;239:
626-632.
35. Weinmann P, Gaehtgens P, Walzog B. Bcl-Xl- and bax-alpha-mediated regulation of apoptosis of human neutrophils via caspase-3. Blood 1999;93:3106-3115.
36. Witthoft T, Eckmann L, Kim JM, Kagnoff MF Enteroinvasive bacteria directly activate expression of iNOS and NO production in human colon epithelial cells. Am J Physiol 1998;275:G564-G571 37. Mannick EE, Bravo LE, Zarama G, et al. Inducible
nitric oxide synthase, nitrotyrosine, and apoptosis in Helicobacter pylori gastritis: effect of antibiotics and antioxidants. Cancer Res 1996;56:3238-3243.
38. 정현채, 김정목, 진영주 등. Helicobacter pylori의 병 독성 유전자 및 운동성이 인체 위점막 염증 반응의 병 태 생리에 미치는 영향 (I)- 병독 인자 보유 유무에 따 른 친염증성 cytokine의 발현능. 대한소화기학회지 1997;29:579-590.