□
원 저□
폐 상피세포에서 PI3K/Akt 경로가 I κ B/NF- κ B 경로의 활성화에 미치는 영향*
서울대학교 의과대학 내과학교실 및 의학연구원 폐 연구소
이상민, 김윤경, 황윤하, 이창훈, 이희석 이춘택, 김영환, 한성구, 심영수, 유철규
=Abstract=
Role of PI3K/Akt Pathway in the Activation of IκB/NF-κB Pathway in Lung Epithelial Cells
Sang-Min Lee, M.D., Yoon Kyung Kim, M.Sc., Yoon-Ha Hwang, M.Sc., Chang-Hoon Lee, M.D., Hee-Seok Lee, M.D., Choon-Taek Lee, M.D.,
Young Whan Kim, M.D., Sung Koo Han, M.D., Young-Soo Shim, M.D., Chul-Gyu Yoo, M.D.
**Division of Pulmonary and Critical Care Medicine, Department of Internal Medicine, Seoul National University College of Medicine, Clinical Research Institute,
Seoul National University Hospital, Lung Institute, Medical Research Center, Seoul National University
Background : NF-κB is a characteristic transcriptional factor which has been shown to regulate production of acute inflammatory mediators and to be involved in the pathogenesis of many inflammatory lung diseases.
There has been some evidence that PI3K/Akt pathway could activate NF-κB in human cell lines.
However, the effect of PI3K/Akt pathway on the activation of NF-κB varied depending on the cell lines used in the experiments. In this study we evaluated the effect of PI3K/Akt pathway on the activation of NF-κB in human respiratory epithelial cell lines.
Methods : BEAS-2B, A549 and NCI-H157 cell lines were used in this experiment. To evaluate the activation of Akt activation and IκB degradation, cells were analysed by western blot assay using
*본 연구는 2001년 서울대학교병원 일반연구비의 보조로 이루어졌음 (04-2001-009-0).
Address for correspondence : Chul-Gyu Yoo, M.D., Ph.D.
Division of Pulmonary and Critical Care Medicine, Department of Internal Medicine, Seoul National University Hospital, 28 Yongon-dong, Chongno-gu, Seoul, 110-744, Korea
Phone : 02-760-3760 Fax : 02-762-9662 E-mail : [email protected]
phospho-specific Akt Ab and IκB Ab. To block PI3K/Akt pathway, cells were pretreated with wortmannin or LY294002 and transfected with dominant negative Akt (DN-Akt). For IKK activity, immune complex kinase assay was performed. To evaluate the DNA binding affinity and transcriptional activity of NF-κB, electrophoretic mobility shift assay (EMSA) and luciferase assay were performed, respectively.
Results : In BEAS-2B, A549 and NCI-H157 cell lines, Akt was activated by TNF-α and insulin.
Activation of Akt by insulin did not induce IκBα degradation. Blocking of PI3K/Akt pathway via wortmannin/LY294002 or DN-Akt did not inhibit TNF-α-induced IκBα degradation or IKK activation.
Inhibition of PI3K/Akt did not affect TNF-α-induced NF-κB activation. Overexpression of DN-Akt did not block TNF-α-induced transcriptional activation of NF-κB, but wortmannin enhanced TNF-α -induced in NF-κB transcriptional activity.
Conclusion : PI3K/Akt was not involved in TNF-α-induced IκBα degradation or transcriptional activity of NF-κB in human respiratory epithelial cell lines.(Tuberculosis and Respiratory Diseases 2003, 54:551-562)
Key words : PI3K/Akt, NF-κB, Activation, Respiratory epithelial cell.
서 론
인체의 중요한 방어 기전인 염증 반응은 대개 경 도의 조직 손상을 동반하지만 과도한 염증은 영구 적인 조직 손상을 초래한다. 대부분의 염증성 질환 은 염증매개 사이토카인의 생산이 적절하게 조절 되지 못하고 과도하게 발현되어 발생하는 병적인 상태의 조직 손상에 기인하는 것으로 이해되고 있 으며, 급성 폐손상, 천식 등의 염증성 폐질환의 발 병 기전에 있어 호중구, 폐포 대식세포, 림프구, 단 핵구 등에서 생산되는 염증매개 사이토카인이 중 요한 역할을 한다는 사실은 잘 알려져 있다1. 즉 외부 자극에 의하여 염증 세포에서 TNF-α와 IL-1이 생산되면 다른 염증 세포에서의 interle
ukin, protease, oxidant, prostagandins, leukotrie
nes 등 염증매개성 물질의 생산이 증가하게 되고, 국소 염증 부위로 분비된 염증매개성 물질은 다시 여러 세포에 작용하여 염증매개성 물질의 생산을 증가시키게 되는 일종의 cytokine network를 이루
면서 염증이 점점 증폭되고 그 결과로 조직 손상 이 야기되게 된다2,3.
이러한 염증매개 사이토카인의 발현은 유전자의 전사에 의해 조절되는데, 대부분의 염증매개 사이 토카인 유전자의 5' 위치에는 κB binding motif 가 존재하여 NF-κB 전사인자에 의해 발현이 조절 되는 것으로 알려져 있다. 즉, NF-κB 경로가 염 증매개 사이토카인의 발현에 중추적인 역할을 담 당하는 공통 경로 (converging pathway) 로 이해 되고 있다4-6.
NF-κB는 면역기능, 염증반응, 혈관 내피세포의 활성화, 세포성장 등에 관여하는 전사인자로서 거 의 모든 세포에 존재한다. 다른 전사인자와 마찬가 지로 NF-κB는 표적 유전자(target gene)의 promoter 부위에 결합하여 전사를 촉진시키게 된 다. NF-κB는 구조상 같은 계열(Rel family)에 속 한 단백의 heterodimer나 혹은 homodimer 형태로 존재하는 데 가장 대표적인 것이 p50과 p65의
heterodimer형태이다6,7. 이 물질은 세포질 내에서 억제 인자인 IκB 단백질과 결합한 비활성화 형태 (dormant complex) 로서 존재하게 되는데, 여러 가지 다양한 세포 외의 자극에 의하여 IκB가 인산 화되고, ubiquitination 되어 proteasome system 에 의하여 IκB 가 분해되고 나면5,8, 유리된 NF-κB 복합체의 형태로 세포핵 내로 이동하여 여러 유전 자의 κB element (GGGAATTCCC)에 결합함으로 써 표적 유전자(target gene)의 전사 활성을 조절 하게 된다9. 이런 경로를 통해 전사가 활성화되는 대표적인 염증매개 물질로는 TNF-α, IL-2, IL-6, IL-8, lymphotoxin, GM-CSF, γ-IFN, adehesion molecule 등을 들 수 있다4-6,10.
이러한 NF-κB의 활성화에는 lipopolysaccharide (LPS)를 포함한 여러 병원체나 tumor-necrosis factor (TNF), interleukin-1(IL-1) 등의 염증성 사 이토카인, T-cell receptor(TCR) signaling 등 여러 신호전달 체계가 관여한다는 사실이 밝혀지고 있 으며11, 최근 phosphatidylinositol 3-kinase/Akt (PI3K/Akt) 경로도 NF-κB 활성화에 관여한다는 연구 결과가 보고되고 있다.
PI3K는 세포막 구성 성분인 phosphoinositide 의 지질 성분을 인산화 시키는 lipid kinase 로서, 2차 전달자(second messenger)를 생성하여 Akt 등의 여 러 하위 신호 전달 체계를 활성화시키게 된다12,13. Akt는 protein kinase B (PKB) 라고도 불리는 일 종의 serine/threonine kinase 로서 다양한 기질을 인산화시켜 세포 증식, 세포 성장, apoptosis 등에 관여한다고 알려져 있다14. 그 동안 Akt 와 NF-κB 의 상호 관련성을 입증 하기 위한 여러 연구 결과 들이 발표되었다15-20. Kane 등16은 백혈병 세포주 에서 Akt 에 의해 NF-κB 활성화가 유도될 수 있 음을 보여주었고, Beraud 등17도 같은 세포주를 대 상으로 연구하여 유사한 실험 결과를 발표하였다.
한편 Reddy 등은 인체 간암 세포주에서도 IL-1β 에 의한 NF-κB 활성화에 Akt가 관여함을 보고하
였다15. 그렇지만, 실험 대상 세포주마다 활성화 기 전이 각기 다르고 호흡기 상피세포에 대한 결과는 아직 알려져 있지 않은 상태이기에, 저자들은 호흡 기 상피세포에서의 NF-κB 활성화에 Akt 경로가 관여하는지, 관여한다면 어떤 기전에 의한 것인지 를 밝히기 위하여 본 연구를 시행하였다.
대상 및 방법 1. 세포주
본 연구에는 정상 폐 상피세포주인 BEAS-2B와 폐암세포주인 A549, NCI-H157을 사용하였다.
BEAS-2B 세포주는 KGM media, 폐암세포주는 10% 우태혈청, 페니실린 30 mg/ml, 스트렙토마이 신 50 mg/ml이 첨가된 RPMI-1640배지를 각각 이 용하여 37℃, 5% CO2 incubator에서 배양하였다.
2. Akt 활성화 유도 및 Akt 경로 억제
Akt 경로의 활성화를 유도하기 위하여 각 세포주 에 5 ng/ml 농도의 TNF-α 혹은 50 μunit/ml 농도의 insulin 을 처리하고, 일정 시간이 경과한 뒤 인산화 Akt의 발현을 측정하여 Akt 경로의 활 성화 정도를 평가하였다.
Akt 경로를 억제하기 위해서는 PI3K의 화학적 저해제로 알려져 있는 wortmannin 및 LY294002 를 각각 40μM과 10μM의 농도로 각 세포주에 전처치하는 방법과, dominant negative Akt(DN- Akt)를 제작하여 세포주에 이입하는 방법을 사용 하였다.
3. Western 분석법
whole lysis buffer(0.1% Nonidet P-40, 5 mM EDTA, 50 mM Tris(pH 7.5~8.0), 250 mM NaCl,
50 mF) 를 이용하여 총세포 단백을 추출하였다.
30μg의 세포 단백을 10% SDS-polyacrylamide gel 에서 전기영동시켰다. 4시간 동안 400 mA의 일정한 전류로 단백질들을 nitrocellulose membrane 으로 transfer 시키고, 이 membrane을 blocking solution(5% skim milk in 1×PBS/Tween 20) 으 로 1시간동안 block 시킨 후 anti-Akt 항체, anti- phospho-Akt 항체, anti-IκBα 항체, 혹은 anti- IKKα항체를 1 : 1,000으로 첨가하여 12시간 동안 반응시켰다. 세척 후 이차 항체를 1 : 2,000으로 첨 가하여 반응시킨 후 면역신호의 검출은 ECL Western blotting detection system 을 이용하였다.
4. Electrophorectic Mobility Shift Assay
Electrophoretic Mobility Shift Assay(EMSA)는 Yoo 등21의 방법을 이용하였다. NF-κB consensus oligonucleotide(Santa Cruz Biotechnology, Cat.
No. sc-2505, Santa Cruz, California, U.S.A.)를 T4 polynucleotide kinase 를 사용하여 [γ32P] ATP로 radiolabelling 을 시행하였다. 4% polyacrylamide gel을 만들고 pre-electrophoresis 를 30분간 시행 한 뒤, 2 μl oligonucleotide probe, 2 μl nuclear protein extract, 500 mM NaCl, 5 mM dithio
threitol, 2 mM EDTA, 5% glycerol, 1μg poly(dI -dC), 0.05% Nonidet P-40, 0.05 mg/ml bovine serum albumin 을 총량 20 μl가 되게 혼합한 다 음 실온에서 20분 간 반응시켰다. 이후 electro
phoresis 를 시행하고 gel을 건조 시킨 후 -70℃에 서 autoradiography 를 시행하였다.
5. IκB kinase (IKK) Assay
세포내 IKK 활성도는 GST-IκBα를 기질로 이용 한 생체외 면역 침강법(in vitro immune complex kinase assay) 으로 측정하였는데, Yoo 등22의 방
법을 이용하였다. lysis buffer 를 20 mM Tris- HCl(pH 7.6), 150 mM NaCl, 25 mM ß-glycero
phosphate, 2 mM EDTA, 2 mM pyrophosphate, 1 mM sodium orthovanadate, 10% glycerol, 1%
Triton X-100, 1 mM DTT, 10 μg/ml leupeptin, 1 mM PMSF 의 조성으로 만든 뒤, 세포를 용해 시켜 4℃에서 10분간 16,000×g로 원심 분리하였 다. 상층액을 따서 1:100으로 희석한 anti-IKKα Ab와 50 μl protein-G Sepharose beads 를 섞어 4℃에서 overnight incubation 시켰다. 2번에 걸쳐 washing 을 시킨 뒤에 10μl의 buffer(20 mM HEPES(pH 7.6), 20 mM ß-glycerophosphate, 0.1 mM sodium orthovanadate, 10 mM MgCl2, 50 mM NaCl, and 1 mM DTT)에 0.5 μg GST-IκB α 와 10 μCi의 [γ-32P]ATP을 섞어 30℃에서 30분간 반응시켰다. 반응이 끝난 뒤 10% SDS- polyacrylamide gel로 전기영동을 시행하였고 nitrocellulose membrane에 전이(transfer) 시킨 뒤 autoradiography를 시행하였다.
6. Luciferase Assay
NF-κB의 전사 활성(transcriptional activity) 을 평가하기 위하여 luciferase assay system을 이용 하였다. κB site 가 포함된 pCDNA3 construct 를 제작한 뒤 실험하고자 하는 세포주에 이입 (transfection) 하였다. 배양이 끝난 뒤에는 1X PBS 로 세척하고 1X lysis buffer(10% Triton X-100, 1mM DTT, 0.2M KPO4, pH 7.8) 를 넣은 뒤, scraper로 긁어 세포를 떼어내고 12,000×g로 원심 분리한 뒤 상층액만 걷어내어 보관하였다.
Luciferase assay buffer(10mM ATP, 20mM MgCl2, 0.2M KPO4, pH 7.8) 를 luminometer tube 에 각각 100μl 씩 분주한 뒤 보관하고 있던 세포 상층액을 20μl 씩 넣고 잘 섞어주었다. 50μl의
Fig. 1. Akt is activated by TNF-α. BEAS-2B (A), A549 (B), and NCI-H157 (C) cells were treated with TNF-α(5 ng/ml) for the indicated times. The levels of phosphorylated Akt(p-Akt) and total Akt in cellular extracts were detected by Western blot analysis.
A TNF-α 20
Akt p-Akt
B
C
0 5 30 60 120 (min)
Akt p-Akt
Akt p-Akt
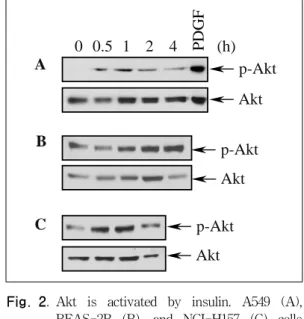
Fig. 2. Akt is activated by insulin. A549 (A), BEAS-2B (B), and NCI-H157 (C) cells were treated with insulin(50 munit/ml) for the indicated times. The levels of phosphorylated Akt(p-Akt) and total Akt in cellular extracts were detected by Western blot analysis.
PDGF 1
0 0.5 2 4 (h)
Akt p-Akt B
A
C
Akt p-Akt
Akt p-Akt
1mM luciferin을 섞은 뒤 luminometer 에 넣고 값 을 측정하였다.
결 과
1. 폐 상피세포에서 TNF-α자극에 의한 Akt의 활성화
폐 상피세포에서 TNF-α의 자극으로 Akt의 활성 화가 유도되는지를 평가하기 위하여, BEAS-2B, A549, NCI-H157 세포를 5 ng/ml 농도의 TNF-α 로 자극하고 5, 10, 20, 30, 60, 120분이 경과한 후 Akt의 활성화 유무를 평가하였다. Akt의 활성화는 인산화를 통해 이루어지기 때문에 인산화 Akt와 총 Akt의 발현을 Western 분석으로 평가하였다.
BEAS-2B 세포에서는 5분이 경과한 후부터 인산 화 Akt의 발현이 증가하기 시작하여 60분까지 지 속되었고, 120분이 경과한 후에는 기저 상태로 감 소되었다(Fig. 1A, upper panel). A549 세포에서는
TNF-α 자극 5분 후부터 인산화 Akt의 발현이 증가하기 시작해 120분까지 지속되었고, 20분과 30 분 후에 최고의 발현을 보였다(Fig. 1B, upper panel). NCI-H157 세포에서는 인산화 Akt의 발현 이 20분 후부터 증가하기 시작해 60분에 최고에 도달하였고, 120분 후에는 기저 상태로 감소하였다 (Fig. 1C, upper panel). 모든 세포에서 TNF-α로 자극하고 120분 후까지 총 Akt의 양은 일정하게 유지되었고(Fig. 1A, B, C, lower panels), β-actin 의 발현도 일정하여(data not shown) 동량의 단백 이 실험에 사용되었음을 확인할 수 있었다. 이상의 결과는 폐 상피세포에서 TNF-α 자극으로 Akt의 활성화가 유도됨을 시사하는 소견이다.
2. Insulin 처치에 의한 Akt의 활성화
Insulin 처치는 PI3K의 활성화를 통해 Akt의 활성
Fig. 3. Activation of Akt by insulin treatment does not affect TNF-α-induced IkBα degradation.
BEAS-2B, A549, and NCI-H157 cells were treated with insulin (50 μunit/ml) for an hour.
The levels of IkBα, phosphorylated Akt (p-Akt), and total Akt in cellular extracts were detected by Western blot analysis.
−+ Insulin −
TNF-α − − + ++ − +
BEAS-2B A549 NCI-H157
− + − +
− − + +
− + +
− − +
Akt p-Akt IκBα
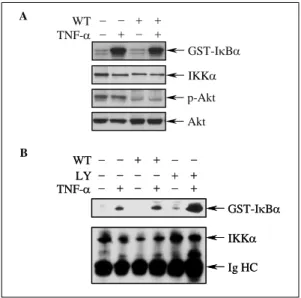
Fig. 4. Blocking of PI3K/Akt pathway does not inhibit TNF-α-induced IκB kinase(IKK) activation. BEAS-2B (A) and NCI-H157 (B) cells were pretreated with wortman nin(WT) or LY294002(LY) for 2 h prior to stimulation with TNF-α for 10 min.
The levels of IκBα, phosphorylated Akt (p-Akt), and total Akt in cellular extracts were detected by Western blot analysis.
WT −
TNF-α − − + ++ − +
p-Akt IKKα GST-IκBα
Akt
IKKα GST-IκBα
Ig HC LY −
TNF-αWT −− − − −+ − +− + +
− ++ +
− −
IKKα GST-IκBα
Ig HC LY −
TNF-αWT −− − − −+ − +− + +
− ++ +
− − A
B 화를 유도한다고 알려져 있다. 폐 상피세포에서
insulin 처치에 의한 Akt의 활성화 유무를 평가하 기 위하여, BEAS-2B, A549, NCI-H157 세포에 insulin(50 μunit/ml)을 처리하고, 30분, 1, 2, 4시 간이 경과한 후 인산화 Akt의 발현을 Western 분 석으로 평가하였다. BEAS-2B, A549, NCI-H157 세포에서 각각 30분, 1시간, 30분 후부터 인산화 Akt 발현의 증가가 관찰되기 시작하였고, BEAS- 2B와 A549 세포에서는 4시간까지, NCI-H157 세 포에서는 1시간까지 인산화 Akt 발현의 증가가 유 지되었다(Fig. 2A, B, C, upper panels). 총 Akt의 발현은 실험에 사용된 시간 동안 일정하게 유지되 었다(Fig. 2A, B, C, lower panels). Positive control 로서 PDGF 처리하였을 때도 Akt의 활성 화가 관찰되었다(Fig. 2A). 이상의 결과로부터 폐 상피세포에서 insulin 처리 후 Akt의 활성화를 일 어남을 확인할 수 있었다.
3. Akt 활성화가 IκBα의 분해에 미치는 영향
Akt 활성화가 IκBα의 분해에 미치는 영향을 평 가하기 위하여, 1시간의 insulin 처리로 Akt를 활 성화 시킨 후, IκBα의 발현을 Western 분석으로 관찰하였다. BEAS-2B, A549, NCI-H157 세포 모
두에서 TNF-α 자극으로 IκBα의 분해가 관찰되 었는데, insulin 처치로는 IκBα의 분해가 유도되 지 않았고, TNF-α에 의한 IκBα의 분해에도 영
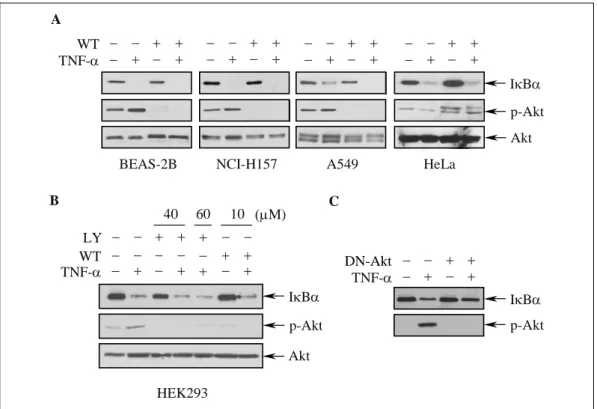
Fig. 5. Blocking of PI3K/Akt pathway does not inhibit TNF-a-induced IkBa degradation. Cells were pretreated with (A) wortmannin(WT) or (B) LY294002(LY) for 2 h, and then stimulated with TNF-a for 30 min. (C) Cells were transfected with dominant negative Akt(DN-Akt), and then stimulated with TNF-a for 30 min. The levels of IkBa, phosphorylated Akt(p-Akt), and total Akt in cellular extracts were detected by Western blot analysis.
BEAS-2B NCI-H157 A549 HeLa WT −
TNF-α − − + ++ − +
− + − +
− − + +
− + − +
− − + +
− + − +
− − + +
Akt p-Akt IκBα
HEK293 40 60 10
WT −
TNF-αLY −− − − −+ − +− + +
+ − +
− + + + − −
(μM)
Akt
p-Akt IκBα DN-Akt −
TNF-α − − + ++ − + A
B C
p-Akt IκBα
향을 미치지 않았다(Fig. 3). 이상의 결과는 폐 상 피세포에서 Akt의 활성화와 IκBα의 분해와는 밀 접한 상관관계가 없음을 시사하는 소견이다.
4. PI3K/Akt 경로의 억제가 IκBα 분해에 미치 는 영향
PI3K/Akt 경로의 억제가 IκBα의 분해 경로에 미 치는 영향을 규명하기 위하여, PI3K의 작용을 억 제하는 wortmannin 및 LY294002 로 전처치하였을 때, TNF-α에 의한 IκB kinase(IKK) 활성화에 미 치는 효과를 평가하였다. BEAS-2B 세포에서 wortmannin 자체는 IKK 활성도에 영향을 미치지
않았고, TNF-α 자극에 의한 IKK 활성도의 증가 에도 아무런 영향을 미치지 않았다(Fig. 4A). 면역 침강된 IKKα의 양은 모든 군에서 일정하여 kinase assay 에서 동량의 IKK가 사용되었음을 확인할 수 있었다(Fig. 4A). 또한 Akt의 활성도가 wor
tmannin 전처치에 의해 효과적으로 억제되었음도 확 인되었다(Fig. 4A). NCI-H157 세포에서도 BEAS- 2B 세포에서와 같이 wortmannin을 전처치한 경우 IKK 활성도에 아무런 변화가 관찰되지 않았다 (Fig. 4B). 다만 LY294002 를 전처치한 경우 IKK 활성화가 증가하는 것처럼 보였으나, 이는 면역침 강된 IKKα의 양이 LY294002 전처치군에서 증가 되어 있어 이에 의한 효과로 판단되었다(Fig. 4B).
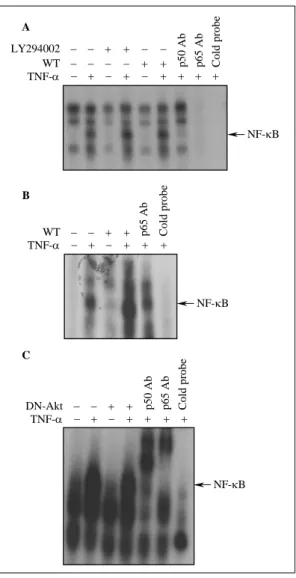
Fig. 6. Blocking of PI3K/Akt pathway does not affect TNF-α-induced NF-κB activation.
NCI-H157 (A) and A549 (B) cells were pretreated with wortmannin(WT) or LY294002(LY) for 2 h prior to stimulation with TNF-α for 30 min. (C) BEAS-2B cells were transfected with dominant negative Akt(DN-Akt), and then stimulated with TNF-α for 30 min. Nuclear extracts were subjected to EMSA for NF-κB.
LY294002
TNF-α − − − − +
− +
− + +
+ + + WT −
+ +
−
− − −
p50 Ab p65 Ab Cold probe +
NF-κB A
TNF-α − − + + +
− +
− + +
WT p65 Ab Cold probe
B
NF-κB
TNF-α − − + + +
− +
− + + +
DN-Akt p50 Ab p65 Ab Cold probe C
NF-κB
Akt 활성화의 억제가 IκBα의 분해에 미치는 영 향을 규명하기 위하여, PI3K의 작용을 억제하는 wortmannin 또는 LY294002 전처치한 뒤 TNF-α
에 의한 IκBα의 분해에 미치는 효과를 평가하였 다. BEAS-2B, A549, NCI-H157 세포에서 wor
tmannin 자체는 IκBα의 발현에 영향을 미치지 않 았고, TNF-α에 의한 IκBα의 분해에도 아무런 영 향을 미치지 않았다(Fig. 5A). HeLa 세포와 HEK293 세포에서도 같은 결과가 관찰되어, 위의 결 과가 폐 상피세포에만 특이적인 현상이 아님을 알 수 있었다(Fig. 5B). HEK293 세포에서는 LY294002 전처치로도 같은 결과가 관찰되었다(Fig. 5B). Akt 활성화와 IκBα 분해와의 관련성을 확실하게 규명 하기 위하여 dominant negative Akt (DN-Akt)를 세포주에 이입하여 Akt 활성화를 억제한 상태에서 TNF-α에 의한 IκBα의 분해 여부를 평가하였다.
DN-Akt의 이입으로 Akt의 활성화가 효과적으로 억제되었고, 이때 TNF-α에 의한 IκBα의 분해가 억제되지는 않았다(Fig. 5C). 이상의 결과는 폐 상 피세포에서 Akt 활성화와 IκBα의 분해와는 상관 관계가 없음을 시사하는 소견이다.
5. PI3K/Akt 경로의 억제가 NF-κB DNA binding activity에 미치는 영향
PI3K/Akt 경로의 억제가 NF-κB의 활성화에 미치는 영향을 관찰하기 위하여, wortmannin 또는 LY294002 전처치로 PI3K의 활성화를 억제한 상태에 서 TNF-α 자극에 의한 NF-κB DNA binding activity를 EMSA로 평가하였다. NCI-H157 세포에 서는 wortmannin 또는 LY294002의 전처치로 TNF- α 자극에 의한 NF-κB DNA binding activity의 증 가가 영향을 받지 않았다(Fig. 6A). 반면에, A549 세 포에서는 TNF-α 단독 처치에 비해, wortmannin 전처치시 TNF-α 자극에 의한 NF-κB DNA binding activity가 더욱 증가되었다 (Fig. 6B).
DN-Akt의 이입으로 Akt의 활성화를 억제하였을 때 도 TNF-α 자극에 의한 NF-κB DNA binding activity는 억제되지 않았다(Fig. 6C). 이상의 결과는
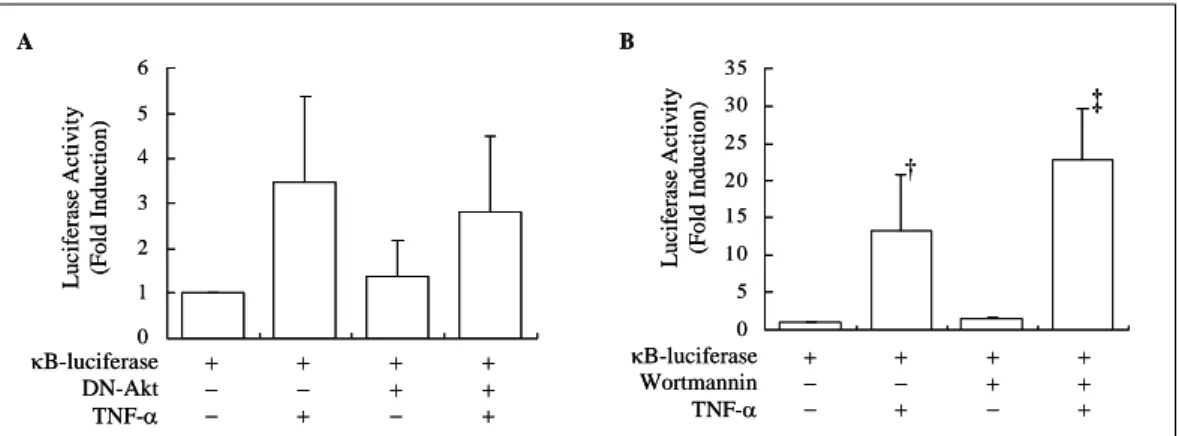
Fig. 7. The effect of PI3K/Akt inhibition on the transcriptional activity of NF-κB. (A) Adenovirus-mediated overexpression of dominant negative Akt (DN-Akt) did not block TNF-α-induced transcriptional activation of NF-κB. A549 cells were transiently cotransfected with plasmid vector containing dominant negative Akt and luciferase reporter construct driven by NF-κB. Twenty-four hours later, cells were stimulated with TNF-α for 30 min. (B) Wortmannin(WT) enhanced TNF-α-induced increase in NF-κB transcriptional activity. A549 cells were transiently transfected with pcDNA3 luciferase reporter construct driven by NF-κB. Twenty-four hours later, 10 mM of wortmannin was added and incubated for 2 h, and then stimulated with TNF-α for 30 min. Luciferase activity was measured and normalized by transfection efficiency, which was determined by β-galactosidase cotransfection.
†: p<0.05 compared to control group
‡: p<0.05 compared to TNF-α-treated group
0 5 10 15 20 25 30 35
LuciferaseActivity (Fold Induction)
κB-luciferase TNF-α Wortmannin
+ + + +
+
− +
−− + − +
B
†
‡
0 5 10 15 20 25 30 35
LuciferaseActivity (Fold Induction)
κB-luciferase TNF-α Wortmannin
+ + + +
+
− +
−− + − +
B
†
‡
0 1 2 3 4 5 6
LuciferaseActivity (Fold Induction)
κB-luciferase TNF-α DN-Akt
+ + +
+ − + +
−− + − +
A
0 1 2 3 4 5 6
LuciferaseActivity (Fold Induction)
κB-luciferase TNF-α DN-Akt
+ + +
+ − + +
−− + − +
A
TNF-α 자극에 의한 NF-κB DNA binding activity의 증가가 PI3K/Akt 경로와 무관하게 이루 어짐을 시사하는 소견이다.
6. PI3K/Akt 경로의 억제가 NF-κB의 전사 활성 에 미치는 영향
PI3K/Akt 경로의 억제가 NF-κB의 전사 활성에 미치는 영향을 관찰하기 위하여, DN-Akt의 과발 현으로 Akt의 활성화를 억제한 상태에서 TNF-α 로 자극한 후 luciferase assay를 시행하였다.
A549 세포에서 TNF-α 처치로 luciferase activity 가 대조군에 비해 증가하였는데, DN-Akt의 과발 현 후에도 TNF-α 처치한 군과 차이를 보이지 않았다(Fig. 7A). 다음 단계로 wortmannin으로 PI3K의 활성화를 억제한 상태에서 TNF-α 자극에
의한 luciferase activity를 측정하였다. Wortman
nin 단독 처치는 luciferase activity에 큰 영향을 미치지 않았지만, wortmannin 전처치 후 TNF-α 로 자극하였을 때는 luciferase activity가 TNF-α 로 자극한 군에 비해 더욱 증가하였다(Fig. 7B).
이상의 결과는 TNF-α 자극에 의한 NF-κB 전사 활성의 증가는 PI3K/Akt 경로와는 무관함을 시사 하는 소견이다. 또한 wortmannin 전처치에 의한 TNF-α유발 luciferase activity의 증강은 PI3K/
Akt 경로의 억제에 의한 것이 아니고 wortmannin 자체에 의한 특이적인 효과로 생각되었다.
고 찰
NF-κB는 체내의 염증 반응 및 면역 반응에서 중 심적인 역할을 담당하고 있는 전사 인자로서, 그
동안 활성화 기전에 대한 관심이 지대하였고 이에 대한 연구도 활발히 진행되어 왔다. 그 중 세포의 증식 및 성장에 관여한다고 알려진 PI3K/Akt 경 로에 의하여 NF-κB 가 활성화된다는 연구 결과들 이 발표되었으나 세포주에 따라 각기 활성화 기전 이 다르게 보고되었다. 백혈병 세포주인 Jurkat T 세포주에서 Akt 에 의해 NF-κB 활성화가 유도될 수 있으며16, pervanadate 에 의한 NF-κB 활성화에 도 PI3K가 관여함을 보고한 연구 결과가 있었다17. 인체 간암 세포주인 HepG2 세포주에서도 IL-1β 와 TNF-α에 의한 NF-κB 활성화에 Akt 경로가 관여된다는 연구 결과들이 보고되었는데15,19, Sizemore 등18은 HepG2 세포주에서 IL-1β에 의 한 NF-κB 활성화에 PI3K가 관여하는 기전이 IκB 분해나 NF-κB의 핵내 이동과 관련된 것이 아니라 NF-κB p65/RelA subunit의 인산화에 의한 것임을 보고하였고, 또 다른 세포주인 NIH-3T3를 대상으 로 한 실험에서도 Akt에 의한 NF-κB 활성화에 Iκ B 분해보다는 p65 subunit 의 활성화가 관여함을 보여주었다20.
이렇듯 지금까지 NF-κB 활성화에 PI3K/Akt 경 로가 관여한다는 증거는 각각 세포주마다 다르고, NF-κB 활성화 기전도 IκB 분해 혹은 p65의 활성 화 등 서로 다르게 보고되고 있는데, 인체 호흡기 상피 세포를 대상으로 한 연구 결과는 그 동안 없 었다.
인체 호흡기 상피세포주를 대상으로 한 본 연구 에서는 TNF-α 처리 후 BEAS-2B, A549, NCI-H157 세포 모두에서 Akt가 활성화됨을 확인 할 수 있었으나, 각 세포주 별로 Akt 활성화가 시 작되는 시기나 최고 발현이 나타나는 시기, 기저 상태로 감소되는 시기 등이 조금씩 달라, 같은 Akt activator 라고 할 지라도 세포에 따라 Akt 활성화 kinetics 에 차이가 있음을 보여주고 있다 (Fig. 1). 한편 또다른 Akt 활성화 인자인 insulin 으로 자극하였을 경우에도 3가지 세포주 모두에서
Akt 의 활성화를 관찰할 수 있었으나, 역시 세포 주 별로 시간별 활성화 정도에 차이가 있음을 알 수 있었고, 같은 세포주라도 TNF-α로 자극한 경 우보다 insulin 으로 자극하였을 경우 Akt 활성화 가 더 늦게 시작됨을 확인할 수 있었다(Fig. 2).
BEAS-2B, A549, NCI-H157 세포주에서 TNF- α 자극 후 IκBα 분해가 유도됨을 확인하였으나, Akt를 단독으로 활성화시킨 경우에는 이러한 IκB α 분해를 관찰할 수 없었다(Fig. 3.). 한편, NCI-H157 에 PI3K/Akt 경로의 억제제인 wor
tmannin를 전처치하여 PI3K/Akt 경로를 억제한 경우, TNF-α에 의한 IKK 활성화에 영향을 미치 지 않음을 관찰할 수 있었다. 그렇지만, LY294002 를 전처치한 경우에는 마치 IKK 활성화를 증가시 키는 것처럼 보였는데(Fig. 4B, upper panels), 이 는 면역침강된 IKKα의 양이 LY294002 전처치군 에서 증가되어 있어 이에 의한 효과로 판단되었다.
PI3K/Akt 경로를 억제한 경우 TNF-α에 의한 Iκ Bα 분해에도 영향을 미치지 않아(Fig. 5), 인체 호흡기 상피세포에서는 PI3K/Akt 경로가 IκBα 분해에 관여하지 않음을 확인할 수 있었다. 이에 다른 보고에서와 같이 PI3K/Akt 경로가 NF-κB의 전사 활성에 관여하는지를 알아보기 위하여 luciferase assay를 시행하였으나 이 역시 인체 호 흡기 상피세포에서는 PI3K/Akt 경로가 NF-κB 전 사 활성에는 관여하지 않음을 보여주었다(Fig. 7).
한 가지 흥미로운 사실은 A549 세포에 wor
tmannin을 전처치한 경우 TNF-α에 의한 luci
ferase activity가 오히려 증가하였는데, DN-Akt 이입한 경우에는 변화가 없는 것으로 보아 wortmannin의 특이적인 효과라고 판단되었다.
EMSA 결과에서도 A549 세포에서는 wortmannin 전처치 후 NF-κB의 DNA binding activity가 증가 하여, luciferase assay 결과와 일치하고 있다(Fig.
6B). Wang 등23도 결장암 세포주인 HT-29을 이용 한 실험에서 wortmannin 처리 후 NF-κB의 전사
활성이 증가함을 보고하여 본 연구의 결과를 뒷받 침하였다. 그렇지만, 같은 wortmannin 전처치를 한 경우라도 NCI-H158 세포에서는 NF-κB의 DNA binding activity의 증가를 관찰할 수 없어 (Fig. 6A), 이러한 wortmannin 의 효능이 나타나 는 양상이 세포에 따라 달라짐을 보여주고 있다.
이상의 결과들을 통해 기존에 백혈병 세포주나 간암 세포주에서 보고되었던 NF-κB와 Akt 경로 와의 연관성을 인체 호흡기 상피 세포에서는 관찰 할 수 없었고, NF-κB 경로와 Akt 경로가 서로 독 자적인 신호 전달 체제를 유지할 것이라고 추정할 수 있었다.
요 약
연구배경 :
NF-κB는 많은 염증 유발성 물질들을 발현시키는 데 필요한 전사 인자로서, 염증성 폐질환 발병에 관여한다는 사실이 확인되었다. 이러한 NF-κB의 활성화에는 여러 신호전달 체계가 관여한다는 사 실이 밝혀지고 있으며 최근 PI3K/Akt 경로도 NF- κB 활성화에 관여한다는 연구 결과가 보고되고 있 으나, 실험 대상 세포주마다 활성화 기전이 다르고 호흡기 상피세포에 대한 결과도 알려져 있지 않아 호흡기 상피세포에서의 NF-κB 활성화에 PI3K/
Akt 경로가 관여하는지를 밝히기 위하여 본 연구 를 시행하게 되었다.
방 법 :
인체 기관지 상피세포주인 BEAS-2B와 폐암 세포 주인 A549, NCI-H157을 사용하여 Akt 활성화와 I κBα 분해 여부를 확인하기 위해 western blot을 시행하였다. Wortmannin, LY294002 및 DN-Akt를 이용하여 Akt 경로를 억제하였고, NF-κB 활성화 와 전사 활성을 측정하기 위하여 각각 EMSA와 luciferase assay를 시행하였다.
결 과 :
BEAS-2B, A549 및 NCI-H157 세포주에 TNF-α 및 insulin을 처리한 경우 Akt 활성화가 유도되었다.
Insulin 으로 Akt 경로를 활성화시킨 경우 IκBα 분 해가 일어나지는 않았다. Wortmannin, LY294002 및 DN-Akt 를 이용하여 Akt 경로를 억제한 경우 TNF-α에 의한 IκBα 분해 및 IKK 활성화가 억 제되지는 않았으며, NF-κB 활성화도 억제되지 않 았다. Wortmannin을 처리한 경우 TNF-α에 의한 NF-κB 전사 활성이 오히려 증가하는 양상을 보였 으나, DN-Akt 이입시킨 경우에는 관찰되지 않았다.
결 론 :
인체 호흡기 상피세포에서는 IκB/NF-κB 경로의 활 성화는 PI3K/Akt 경로와 무관한 것으로 판단된다.
참 고 문 헌
1. Strieter RM, Phan SH, Ward PA. Chapter 17.
Inflammation, injury, and repair. In : Murray JF, Nadel JA. Textbook of Respiratory Medicine. 2nd ed. Philadelphia : W.B. Sau
ders Company ; 1994. p.478-83.
2. Levine SJ. Bronchial epithelial cell-cytokine interactions in airway inflammation. J Investigative Medicine 1995;43:241-49.
3. Raeburn D, Webber SE. Proinflammatory potential of the airway epithelium in bronchial astham. Eur Respir J 1994 ;7:2226-33.
4. Thanos D, Maniatis T. NF-κB; A lesson in family values. Cell 1995;80:529-35.
5. Baeuerle PA, Baltimore D. IκB; a specific inhibitor of the NF-κB transcriptional factor.
Science 1988;242:540-6.
6. Baeuerle PA, Baltimore D. NF-κB; Ten Years After. Cell 1996;87:13-21.
7. Siebenlist U, Franzoso G, Brown K. Stru
cture, regulation and function of NF-kB.
Annu Rev Cell Biol 1994;10:405-55.
8. Baeuerle PA, Baltimore D. A 65- kD subunit of active NF-κB is required for inhibition of NF-κB by IκB. Genes Dev 1989;3:1689- 98.
9. Baeuerle PA, Henkel T. Function and activation of NF-κB in the immune system.
Annu Rev Immunol 1994;12:141-79.
10. Chen Z, Parent L, Maniatis T. Site-specific phosphorylation of IκB by a novel ubiqui
tin-dependent kinase activity. Cell 1996;84:
853-61.
11. Li Q, Verma IM. NF-κB regulation in the immune system. Nat Rev Immunol 2002;2:
725-34.
12. Condliffer AM, Cadwallader KA, Walker TR, Rintoul RC, Cowburn AS, Chilvers ER.
Phosphoinositide 3-kinse : a critical signaling event in pulmonary cells. Respir Res 2000;1:
24-9.
13. Vivanco I, Sawyers CL. The phosphatidy
linositol 3-kinase-Akt pathway in human cancer. Nat Rev Cancer 2002;2:489-501.
14. Scheid MP, Woodgett JR. PKB/Akt: functio
nal insights from genetic models. Nat Rev Mol Cell Biol 2001;2:760-8.
15. Reddy SA, Huang JH, Liao WS. Phosphati
dylinositil 3-kianse in interleukin 1 singna
ling. J Biol Chem 1997;272:29167-73.
16. Kane LP, Shapiro VS, Stokoe D, Weiss A.
Induction of NF-κB by the Akt/PKB kinase.
Curr Biol 1999;9:601-4.
17. Beraud C, Henzel WJ, Baeuerle PA. Involve
ment of regulatory and catalytic subunits of phosphoinositide 3-kinase in NF-κB activa
tion. Proc Natl Acad Sci 1999;96:429-34.
18. Sizemore N, Leung S, Stark GR. Activation of phosphatidylinositol 3-kinse in response to interleukin-1 leads to phosphorylation and activation of the NF-κB p65/RelA subunit.
Mol Cell Biochem 1999;19:4798-805.
19. Reddy SA, Huang JH, Liao WS. Phosphati
dylinositol 3-kinase as a mediator of TNF- induced NF-κB activation. J Immunol 2000;
164:1355-1363.
20. Madrid LV, Wang CY, Guttridge DC, Schottelius AJ, Baldwin AS, Mayo MW. Akt suppresses apoptosis by stimulating the transactivation potential of the RelA/p65 subunit of NF-κB. Mol Cell Bilchem 2000;
20:1626-38.
21. Yoo CG, Lee S, Lee CT, Kim YW, Han SK, Shim YS. Effects of acetylsalicylic acid on endogenous IκB kinase activity in lung epithelial cells. Am J Physiol Lung Cell Mol Physiol 2001;280:L3-9.
22. Yoo CG, Lee S, Lee CT, Kim YW, Han SK, Shim YS. Anti-inflammatory effect of heat shock protein induction is related to stabi
lization of IκBα through preventing IκB kinase activation in respiratory epithelial cells. J Immunol 2000;164:5416-23.
23. Wang Q, Kim S, Wang X, Evers BM.
Activation of NF-κB binding in HT-29 colon cancer cells by inhibition of phosphati
dylinositol 3-kinase. Biochem Biophys Res Commun 2000;273:853-8.