Copyrightⓒ 2011, The Korean Academy of Oral Biology
51
Journal of Oral Biology
Immunolocalization of Runx2 and Osterix in the Developing Periodontal Tissues of the Mouse
Byung-In Kim1, Seung-Hoon Na1, Ji-Youn Kim1, Je-Won Shin1, and Seong-Suk Jue1*
1Department of Oral Anatomy, Schoolof Dentistry, Kyung Hee University, Seoul 130-701, Korea (received March 7, 2011 ; revised May 24, 2011 ; accepted May 27, 2011)
Runx2 and Osterix, the transcription factors for osteoblast differentiation, are known as fundamental factors to regulate the development of calcified tissues. However, the biological functions of these factors in the development of the peri- odontal tissues remain unclear. In this study, we investigated the distribution of Runx2 and Osterix during periodontal tissue development of the mice. Mandibles from 14-day-old mice were prepared for paraffin section. Serial sections of the mandible containing 1st molar tooth germs were obtained as a thickness of 7µm. Some sections were stained with hematoxylin and eosin. Others were used for immunohistochemistry for PCNA, Runx2, and Osterix. Epithelial cells in growing end of Hertwig’s epithelial root sheath (HERS) and mesenchymal cells adjacent to the growing end of HERS expressed PCNA.
Undifferentiated mesenchymal cells and hard tissue forming cells like cementoblasts and osteoblasts in early stage of differ- entiation expressed Runx2. Fully differentiated cementoblasts and osteoblasts secreting matrix proteins expressed Osterix.
However, the cells terminated the matrix formation did not express Osterix. Periodontal ligament cells expressed Runx2 and Osterix. Pulp cells expressed Runx2 only.These results suggest that Runx2 and Osterix might regulate the differ- entiation of cementoblasts in the same manner as osteoblasts.
Runx2 might participate in the process of cementoblast differentiation in early stage, whether Osterix might regulate the maturation and matrix synthesis of the cells.
Key words: developing periodontal tissues, PCNA, Runx2, Osterix
서 론
손상된 치주조직(periodontal tissue)의 재생을 도모하기 위하여 시도되고 있는 여러 가지 방법들은 치아부착의 재 생을 그 목적으로 한다. 치주인대(periodontal ligament)는 치근표면을 덮고 있는 시멘트질(cementum)과 치조골(alveolar bone)에 각각 삽입되어 치아를 턱뼈에 고정시키는 역할을 한다(Nanci, 2008). 이 중 시멘트질은 치아표면을 얇게 덮 고 있는 경결합조직으로, 치아부착과 치주조직 재생과정에 서도 매우 중요한 역할을 담당하고 있다.
시멘트질은 발생과정중 치아주머니(dental follicle)의 외 배엽성중간엽세포의 분화 혹은 Hertwig상피뿌리집(Hertwig’s epithelial root sheath; HERS) 세포의 변형에 의해 형성 된 시멘트질모세포(cementoblast)에 의해 만들어지며, 성인 조직에서는 치주인대에 존재하는 미분화중간엽세포의 분 화에 의해 재생되는 것으로 알려져 있다(Nanci, 2008).
시멘트질의 성분이나 생화학적 조성은 뼈와 매우 유사하 여, 가장 많은 부분을 차지하는 유기바탕질 성분은 제1형 아교질(Type 1 collagen)이며, 뼈조직의 주요 비아교단백질 성분인 bone sialoprotein(BSP)이나 osteocalcin(OCN)은 시 멘트질 바탕질에도 존재한다(Kim et al., 2008; Ko et al., 2005). 시멘트질모세포는 또한 뼈모세포(osteoblast)와 마찬 가지로 알칼리성인산분해효소(alkaline phosphatase, ALP)를 발현하며, runt-related gene 2(Runx2)도 발현하는 것으로 알려져 있다(D'Errico et al., 1997; D'Errico et al., 2000;
Kitagawa et al., 2006). 하지만 시멘트질은 뼈와는 다른 조 직학적 구조를 가지고 있는데, 조직학적으로 뼈와 시멘트 질의 가장 큰 차이점은 시멘트질은 조직 내에 신경과 혈 관이 포함되어 있지 않다는 점이다(Saygin et al., 2000;
Bosshardt, 2005). 이 때문에 시멘트질은 뼈에 비해 압력 에 대한 저항성이 높고, 개조(remodeling)가 쉽게 일어나 지 않는다는 특징을 보이게 된다. 시멘트질은 또한 치주인
*Corresponding author: Seong-Suk Jue, Department of Oral Anatomy, Graduate School of Dentistry, Kyung Hee Univer- sity, Seoul, 130-701, Korea. Tel.: +82-2-961-0351,
Fax.: +82-2-960-1747, E-mail: [email protected]
52 Byung-In Kim, Seung-Hoon Na, Ji-Youn Kim, Je-Won Shin, and Seong-Suk Jue 대를 사이에 두고 치조골과 인접하여 있는데, 이 두 조직
을 각각 형성하는 세포인 시멘트질모세포와 뼈모세포는 모 두 치주인대에서 유래하는 것으로 알려져 있지만, 이 두 세포가 공통된 전구세포를 가지는지 혹은 서로 다른 전구 세포로부터 분화하는지는 아직 알려져 있지 않다(Nanci, 2008). 또한 어떠한 분자생물학적기전에 의해 치주인대내 의 전구세포로부터 시멘트질모세포가 분화하는지 그 과정 도 알려져 있지 않다.
Runx2는 중간엽세포로부터 뼈모세포가 분화하는데 있어 서 필수적인 전사인자로, 뼈모세포와 연관된 OCN, Type I collagen, osteopontin(OPN) 등의 gene의 전사를 직접적으로 촉진하는 것으로 알려져 있다(Aberg et al., 2004; Franceschi et al., 2003). 치주인대세포도 Runx2를 발현한다는 것이 최 근 알려지면서, 치주인대세포가 뼈모세포로 분화하는 과정 도 Runx2에 의해 조절될 것으로 예상되고 있다(Saito et al., 2002).
Osterix도 뼈모세포의 분화에 필수적인 전사인자로(Matsubara et al., 2008), Osterix의 기능은 Runx2에 의해 조절되는 것 으로 알려져 있다(Koga et al., 2005; Nakashima et al., 2002; Tai et al., 2005). 최근에는 뼈조직 뿐 아니라 연골조 직에서도 Osterix가 Runx2에 의해 조절되고 있음이 밝혀졌 으며(Nishio et al., 2006), 치아싹의 중간엽조직에서도 Osterix 발현이 확인되면서 이들 전사인자가 치아발생 과정에서 어 떠한 역할을 하는지 많은 관심이 모아지고 있다(Nakashima et al., 2002).
따라서 본 연구에서는 마우스 치근의 발생과정중 치아 주위조직에서 뼈모세포의 분화과정에서 필수적인 역할을 하는 전사인자인 Runx2와 Osterix의 분포 양상을 살펴보 고자 한다. 또한 세포분열중인 핵에서 발현되는 Proliferating Cell Nuclear Antigen(PCNA)의 분포도 함께 관찰하여, 치 근 발생과정중 세포증식이 활발하게 이루어지는 위치와 더 불어 Runx2와 Osterix의 분포를 함께 살펴봄으로써 시멘 트질모세포의 분화과정과 뼈모세포의 분화과정에서 이들 전사인자들의 역할을 밝히고자 한다.
재료 및 방법
실험재료
실험동물로는 생후 14일 된 ICR 마우스 14마리를 사용 하였다. 모든 동물실험 과정은 경희대학교 동물윤리위원회 의 승인을 받아 진행되었다.
조직학적 관찰을 위한 시편준비
생후 14일 된 마우스의 아래턱을 획득하여 10% formal- dehyde에 넣고 4oC에서 하룻동안 고정하였다. 수세 후 10%
EDTA(ethylenediamine tetraacetic acid, pH 7.4)에 넣어 4oC에서 2주간 탈회하였다. 탈회한 흰쥐의 아래턱은 탈수,
투명화 과정을 거쳐 파라핀으로 포매하였다. 포매한 조직 은 박절기(Leica RM2145)로 7 µm 두께의 시상 연속절편 을 얻었다. 이렇게 얻은 절편의 일부는 조직학적 관찰을 위하여 헤마톡실린-에오신(hematoxylin-eosin) 염색을 시행 하였고, 나머지는 면역조직화학염색을 시행하였다.
면역조직화학염색
면역조직화학염색은 Vectastain® ABC kit (Vector Labora- tories, Burlingame, USA)를 이용하여 통법에 따라 다음과 같이 시행하였다. 절편의 파라핀을 제거하고 점차적으로 저 농도의 알코올로 수화하였다. 5분간 수세 후 0.3% 과산화 수소수(H2O2)로 30분간 처리하여 내인성 과산화효소를 억 제시켰다. 5분간 수세 후 일차항체와 비특이적으로 결합할 수 있는 단백을 억제하기 위해 정상혈청(normal serum)으 로 20분간 처리하였다. 절편 주변의 과도한 양의 정상혈청 은 제거하고, 다음 각각의 일차항체를 각각의 연속절편에 30분간 처리하였다. Rabbit anti-Proliferating Cell Nuclear Antigen (PCNA) antibody (Serotec, Oxford, UK)는 1:400 으로, Mouse anti-Runx2 monoclonal antibody (MBL, Woburn, Japan)와 Rabbit anti-Osterix polyclonal antibody (Abcam, Cambridge, UK)는 1:200으로 희석하여 사용하였 다. 5분간 수세 후 희석한 비오틴화이차항체(biotinylated secondary antibody)로 30분간 처리하고 5분간 수세하였다.
Vectastain® Elite ABC reagent를 30분간 처리한 뒤 5분간 수세하고 DAB (DAB substrate kit, Vector Laboratories, Burlingame, USA)으로 발색하였다. 헤마톡실린으로 대조 염색 후 봉입하여 현미경으로 관찰하였다.
결 과
마우스 치근 발생과정의 조직학적 관찰
생후 14일 된 마우스 아래턱에서는 각각의 치근 발생 단 계가 다른 첫째, 둘째, 셋째어금니를 관찰할 수 있었다(Fig.
1A). 이 중 첫째 어금니는 치관은 거의 완성되었고, 치근 은 2분의 1에서 3분의 1정도 형성된 것을 확인할 수 있 었다(Fig. 1B). 발생중인 치근의 치경부쪽에서는 일차시멘 트질과 치조골 그리고 그 사이를 주행하는 치주인대 섬유 가 관찰되었다(Fig. 1C). 성장하고 있는 치근끝 부위에는 두 층의 상피세포로 구성된 HERS가 존재하였고, 치근면 을 따라 치경부쪽으로 이동하면서 HERS의 상피세포들이 분절화되어 관찰되었다. 상아질모세포(odontoblast)가 상아 질을 따라 치수 표면에 존재하였으며, 치근끝 쪽에는 전상 아질모세포(preodontoblast)가 HERS 세포와 면하여 위치 하며 상아질모세포 전구세포인 것을 확인할 수 있었다(Fig.
1C, E). 치근분기부에서는 다양한 방향성을 가진 치주인대 섬유와 세포들을 관찰할 수 있었으며, 치근 사이로 뻗어 올라온 발생중인 치조중격을 관찰할 수 있었다(Fig. 1D).
마우스 치근 발생과정중 PCNA, Runx2, OPN의 발현 양상
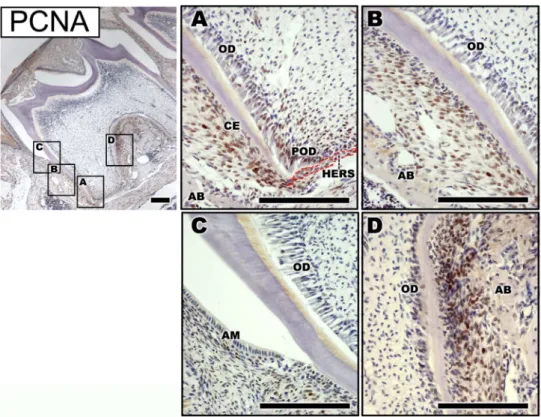
PCNA는 발생중인 치근끝부분과 치근분기부에서 강하게 발현되는 것을 관찰할 수 있었는데, 치근끝부분의 HERS 세포뿐 아니라 이와 인접한 전상아질모세포 및 치아주머 니의 세포에서 강하게 발현되었다. 그러나 PCNA가 강하 게 발현되는 이들 세포와 인접하여 있는, 분화된 상아질모 세포와 시멘트질모세포에서는 거의 발현되지 않았다(Fig.
2A). 또한 발생중인 치근 중간부분에 존재하는, 완전히 분 화되어 바탕질을 분비하고 있는 상아질모세포나 시멘트질 모세포에서는 PCNA 발현을 거의 관찰할 수 없었다. 치아 유두에 존재하는 중간엽세포들은 PCNA를 거의 발현하지 않은 반면, 치주인대의 중간엽세포들은 많은 세포들이 염 색되었다(Fig. 2B). 마찬가지로 치경부쪽에서도 상아질모세 포나 법랑질모세포에서는 PCNA 발현을 관찰할 수 없었 고, 퇴축치아상피를 둘러싸고 있는 결합조직의 몇몇 세포 들만 일부 PCNA를 발현하였다(Fig. 2C). 치근분기부에서 도 상아질모세포 주위에 있는 중간엽세포 일부는 약하게 PCNA를 발현하였으나, 치주인대를 구성하는 세포들은 다 수의 세포가 PCNA를 발현하였다(Fig. 2D).
Runx2는 전체적으로 치근 끝부분에서 비교적 강하게 발 현되며 치주인대 내에서도 발현되는 것을 관찰할 수 있었 다. 발생중인 치근 끝부분의 HERS 세포 자체는 Runx2를 발현하지 않았지만 그 주변의 치아유두 및 치아주머니의 중간엽세포에서는 일부 발현되는 것을 관찰할 수 있었다.
그러나 상아질모세포와 시멘트질모세포에서는 Runx2의 발 현이 관찰되지 않았다(Fig. 3A). 치근의 치경부쪽에서는 Runx2의 발현이 상아질모세포, 시멘트질모세포, 그리고 뼈 모세포 모두에서 관찰되지 않지만, 치주인대내 세포에서는 일부 약하게 발현하고 있는 것이 관찰되었다. 또한 치주 인대내 바탕질, 특히 뼈로 삽입되는 Sharpey’s fiber에서 강한 염색성을 보이고 있었다(Fig. 3B). 치근 분기부에서 도 마찬가지로, 경조직 형성세포에서는 발현되지 않으나, 치주인대내 세포에서 일부 발현되었으며, 뼈의 표면 및 치 주인대 섬유와 같은 바탕질 성분에서 발현되고 있는 것이 관찰되었다(Fig. 3C).
Osterix는 전체적으로 치근 끝부분과 치근분기부에서 강 하게 발현되었고, 또한 뼈 표면에서도 강하게 발현되는 것 이 관찰되었다. 치근 끝부분에서는 HERS와 인접한 상아 질모세포에서 Osterix가 강하게 발현되는 것을 관찰할 수 있었다. 또한 HERS와 인접한 치아유두 중간엽세포에서도 발현되었다. 반면 HERS 자체나 치아유두 중심부의 세포 들은 Osterix를 발현하지 않았다. 치주인대를 살펴보면, 치 아유두에 비해 치주인대 전반에 걸쳐서 Osterix를 발현하 는 세포들이 다수 관찰되었으며, 상아질 표면에 부착되어 있는 시멘트질모세포 일부도 Osterix를 발현하였다(Fig.
4A). 발생중인 치근의 중간부분에서는 상아질 형성 초기 상아질모세포는 강하게 염색된 반면, 완전히 분화된 상아 질모세포는 염색성을 나타내지 않았다. 치주인대의 중간엽 세포들과 시멘트질모세포들은 치아유두쪽 세포에 비해 다 Fig. 1. Histological analysis of developing periodontal tissues during root development of postnatal day 14 (PN14) mice (H-E stain). (A) Three molars in different developmental stages were observed. (B) The root of the first molar was being formed at this stage. (C, D) On the pulpal side of root dentin, odontoblasts were lined in a row. Acellular and cellular cementum had formed on the root dentin. (E) HERS was located at the apical end of the root. Discontinuity of epithelial cells was observed at the upper end of HERS. C and D are high magnification views of square boxes in B. E is high magnification views of square boxes in C. The red dotted line demarcates the boundary of HERS. AB, alveolar bone; CE, cementoblast; HERS, Hertwig’s epithelial root sheath; OD, odontoblast; POD, preodontoblast. Bars in A-E = 50 µm.
54 Byung-In Kim, Seung-Hoon Na, Ji-Youn Kim, Je-Won Shin, and Seong-Suk Jue
수의 세포가 약하게 Osterix를 발현하였다. 치조골 표면에 있는 뼈모세포들도 Osterix를 강하게 발현하는 것이 관찰 되었다(Fig. 4B). 치근의 치경부에서는 상아질모세포, 퇴 축치아상피, 그리고 그 주변을 둘러싸고 있는 결합조직 모두에서 Osterix의 발현을 거의 관찰할 수 없었다(Fig.
4C). 치근분기부에서는 상아질모세포, 시멘트질모세포, 뼈 모세포, 그리고 치주인대 세포들 모두가 Osterix를 강하게 발현하는 것이 관찰되었다(Fig. 4D).
고 찰
손상된 치주조직 재생을 통해 치아부착을 재생시키기 위 해서는 치근표면의 시멘트질, 치근을 담고 있는 치조골, 그리고 그 사이를 매개하고 있는 치주인대의 재생에 관 한 이해가 필수적이지만, 이제까지의 많은 연구들은 치주 인대와 치조골에 초점을 맞추어 이루어져 왔으며 시멘트 질의 재생에 관해서는 많은 부분이 아직 밝혀지지 않은 채 남아있다.
Fig. 3. Localization of Runx2 in the first mandibular molar of PN14. (A) Intense immunoreactivity of Runx2 was observed in the dental mesenchymal cells adjacent to HERS in apical region. (B, C) Immunoreactivity of Runx2 was weakly detected in the PDL but not in the fully differentiated odontoblasts and cementoblasts in cervical and furcation region. The red dotted line demarcates the boundary of HERS.AB, alveolar bone; HERS, Hertwig’s epithelial root sheath; OD, odontoblast; POD, preodontoblast. Bars in A-C = 100µm.
Fig. 2. Localization of PCNA in the first mandibular molar of PN14. (A) A few cells of HERS and mesenchymal cells adjacent to them strongly expressed PCNA in apical region. (B) Immunoreactivity of PCNA was observed in PDL cells but not in fully differentiated cemento- blasts, odontoblasts, and ameloblasts in the middle and cervical region. (D) Osteoblasts on the surface of alveolar septum and PDL cells strongly expressed PCNA in the furcation region. The red dotted line demarcates the boundary of HERS. AB, alveolar bone; AM, ameloblast;
CE, cementoblast; HERS, Hertwig’s epithelial root sheath; OD, odontoblast; POD, preodontoblast. Bars in A-D = 100µm.
시멘트질은 치아를 구성하는 경조직중 뼈와 가장 유사 한 조직으로, 그 생화학적 조성이나 성분이 뼈와 크게 다 르지 않지만, 조직학적 구조는 상당한 차이를 보이고 있 다(Nanci, 2008). 이에 뼈모세포와 시멘트질모세포의 분 화과정의 유사성과 차이점을 살펴보는 일은 의미 있는 일 이라 하겠다.
Runx2와 Osterix는 뼈모세포의 분화과정에서 필수적인 역 할을 하는 전사인자로, Osterix의 기능은 Runx2에 의해 조 절되는 것으로 알려져 있다(Koga et al., 2005; Nakashima et al., 2002; Tai et al., 2005). 따라서 본 연구에서는 발 생중인 치아주위조직에서 이들 전사인자의 분포를 확인함 으로써 이들 전사인자가 시멘트질모세포의 분화에 미치는 영향에 대해 조사하였다.
본 연구에 사용된 생후 14일 된 마우스 첫째 어금니는 치근발생이 끝나지 않은 상태로, 발육중인 치근끝에서 HERS 를 관찰할 수 있으며, 이 부위로부터 치경쪽으로 가면서 분화중인 상아질모세포, 분절화된 HERS, 분화중인 시멘 트질모세포와 이들 외측에서 관찰되는 치주인대 및 치조 골까지 모든 조직의 분화과정을 한번에 관찰할 수 있어서 분화중인 치아주위조직 세포를 관찰하는데 있어서 가장 적 합한 표본이라고 생각된다(Fig. 1).
세포의 증식능을 알아보기 위해 PCNA 면역염색을 시
행하였다(Fig. 2). 발육중인 치근끝부위의 HERS의 상피세 포들이 PCNA를 발현하여, 이 세포들의 증식이 이루어지 고 있음을 알 수 있었다. 분화가 완성되어 바탕질을 분비 하고 있는 상아질모세포와 시멘트질모세포는 예상대로 PCNA 를 대부분 발현하지 않아 이들 세포들은 세포분열 하지 않음을 알 수 있었다. 반면, 치근끝에 가까운 부위에서는 이들 세포를 둘러싸는 중간엽 세포들은 PCNA를 발현하였 는데, 이는 상아질모세포와 시멘트질모세포로 분화할 수 있 는 세포 pool을 만들기 위한 과정인 것으로 생각되었다.
또한 치관부 치수의 세포들은 PCNA를 발현하지 않았지 만, 치주인대세포들은 전반적으로 강하게 PCNA를 발현하 여, 치근의 발생과 맹출에 이르기까지 치주인대의 활발한 활성을 알 수 있었다.
Runx2의 면역염색 반응을 관찰해보면(Fig. 3), HERS의 상피세포는 Runx2 를 발현하지 않았으며, 경조직을 형성 하고 있는 상아질모세포, 시멘트질모세포 및 뼈모세포도 Runx2를 발현하지 않았다. 반면, HERS 끝부분 주위를 둘 러싸는 치아유두 및 치아주머니의 중간엽세포들은 Runx2 를 발현하였다. 또한 치주인대와 치수의 중간엽세포들도 전반적으로 Runx2를 발현하였다.
Osterix도 HERS의 상피세포들에서는 발현되지 않았다 (Fig. 4). 하지만, Runx2의 경우와 달리 상아질모세포, 시 Fig. 4. Localization of Osterix in the first mandibular molar of PN14. (A) Odontoblasts and cementoblasts adjacent to HERS showed intense reactivity of Osterix. No cells of HERS showed osterix reactivity in the apical region. (B, C) Odontoblasts and cementoblasts showed no reac- tivity in the middle and cervical region. (D) Many osteoblasts on the surface of the alveolar septum and PDL cells showed intense reactivity in the furcation region. The red dotted line demarcates the boundary of HERS. AB, alveolar bone; AM, ameloblast; HERS, Hertwig’s epithelial root sheath; OD, odontoblast; POD, preodontoblast. Bars in A-D = 100µm.
56 Byung-In Kim, Seung-Hoon Na, Ji-Youn Kim, Je-Won Shin, and Seong-Suk Jue 멘트질모세포 및 뼈모세포는 모두 Osterix를 발현하였다.
상아질모세포의 경우에는 치근끝에 가까운 바탕질 분비 초 기의 상아질모세포는 Osterix를 강하게 발현한 반면, 치관 부위에 위치하는 분비가 거의 완료된 상아질모세포는 Osterix 를 발현하지 않았다. 또한, 치주인대의 중간엽세포들이 Runx2 의 경우와 마찬가지로 Osterix를 발현하였으나, 치수세포 들은 Osterix를 발현하지 않았다.
Runx2와 Osterix의 분포 변화를 관찰해보면, Runx2는 치근끝의 세포가 막 분화하기 시작하는 부위에서만 국소 적으로 발현되는 반면 Osterix는 치근끝부터 좀 더 치경 측까지 넓게 분포하는 것으로 보아, 뼈모세포의 경우에서 와 마찬가지로(Maruyama et al., 2007; Liu et al., 2001) 시멘트질모세포도 Runx2에 의해 Osterix 발현이 조절되는 것으로 생각되었다. 또한 Runx2와 Osterix 발현의 순서로 보아, Runx2는 시멘트질모세포의 초기 분화에 영향을 미 치고, Osterix는 분화된 시멘트질모세포의 성숙과 바탕질 분비에 영향을 미칠 것으로 생각되었다. 이는 상아질모세 포, 시멘트질모세포, 뼈모세포 모두 Runx2는 발현하지 않 았지만 Osterix를 발현한 결과로 보아 더욱 확실하다고 할 수 있다. 특히 상아질모세포의 경우, 분비초기에는 Osterix 를 강하게 발현하다가 분비가 종료되는 시점에서 Osterix 발현이 사라지는 결과를 보임으로써 이러한 가설을 뒷받 침해주고 있다. Hirata 등은(Hirata et al., 2009) 시멘트질 모세포는 Osterix를 발현하지만 시멘트질세포(cementocyte) 는 Osterix를 발현하지 않는다는 결과를 발표하여 본 연 구와 동일한 결과를 보고한 바 있다.
치주인대는 Runx2와 Osterix 모두를 발현하는데, 이는 치주인대의 세포들이 경조직형성세포로 분화할 수 있는 잠 재력을 가지고 있음을 나타내고 있는 것으로 생각된다 (D’Souza et al., 1999; Jiang et al., 1999).
치주인대세포의 경우와 달리 치수세포는 Runx2는 발현 하였지만 Osterix는 발현하지 않았다. 치주인대는 경조직 형성세포 분화 잠재능을 가졌기 때문에 뼈모세포의 분화 에 필수적인 전사인자인 Runx2나 Osterix를 발현한다고 생각할 수 있지만, 치수의 경우는 이와 다르다. 이는 Runx2 가 경조직형성세포의 분화뿐 아니라 치아의 발생과정에 중 요한 역할을 가지고 있을 수도 있음을 암시하는 것이라 생각할 수 있다. 이와 같은 가설은 많은 다른 연구자들의 연구결과(Nakashima et al., 2002; D’Souza et al., 1999;
Åberg et al., 2004)를 통해서 뒷받침되고 있으며, 치아 형 성과정에서 이들 전사인자의 역할에 관한 연구가 더 많이 이루어져야 할 것으로 생각된다.
흥미로운 사실은 HERS가 PCNA는 발현한 반면, Runx2 와 osterix는 발현하지 않았다는 것이다(Fig. 3, 4). 이는 치근끝부터 치경측까지 모든 부위에서 일관되게 나타난 결 과로, Hirata 등(Hirata et al., 2009)의 연구와 일치한다.
이제까지 고전적으로 시멘트질모세포는 치아주머니의 중간 엽세포들이 찢어진 HERS의 틈새로 이동하여 치근 표면
에 부착하여 분화함으로써 형성된다고 설명되어왔으나, 최 근에는 일부 시멘트질모세포는HERS의 상피세포들이 상피 -중간엽 변형(epithelium-mesenchyme transformation)에 의 해 형성된다는 보고가 있어왔다. 하지만 이번 연구에서 HERS의 상피세포들은 다른 시멘트질모세포의 분화과정에 서 보이는 Runx2와 Osterix 발현을 보이지 않는 것으로 보아 이러한 상피-중간엽 변형에 의한 시멘트질모세포 형 성 가설에 의문을 제기하는 바이다.
이상의 결과로 보아 뼈모세포의 경우와 마찬가지로 시멘 트질모세포의 분화는 Runx2와 Osterix에 의해 조절되는 것으로 생각되며, 이들 전사인자의 발현 순서 및 발현 양 상은 시멘트질모세포에서도 뼈모세포에서와 마찬가지로 나 타나는 것으로 생각된다. 이것으로 보아 시멘트질모세포 연구에 있어서 이들 전사인자를 유용한 표지자로 사용할 수 있을 것으로 생각된다. 나아가 시멘트질모세포의 초기 분화에는 Runx2가 관여하며 세포의 성숙과 바탕질분비과 정에는 Osterix가 관여하는 것으로 나타났다. 또한 이들 전 사인자는 경조직형성세포의 분화뿐 아니라 치아발생과정에 서 중요한 역할을 할 것으로 생각되며, 이에 관한 더 깊 은 연구가 필요할 것으로 생각된다.
참 고 문 헌
Åberg T, Cavender A, Gaikwad JS, Bronckers ALJJ, Wang X, Waltimo-Sirén J, Thesleff I, D’Souza N. Phenotypic changes in dentition of Runx2 homozygote-null mutant mice. J Histochem Cytochem. 2004;52:131-9.
Aberg T, Wang XP, Kim JH, Yamashiro T, Bei M, Rice R, Ryoo HM, Thesleff I. Runx2 mediates FGF signaling from epithelium to mesenchyme during tooth morphogenesis. Dev Biol. 2004;270:76-93.
Bosshardt DD. Are cementoblasts a subpopulation of osteoblasts or a unique phenotype? J Dent Res. 2005;84:390- 406.
D'Errico JA, Berry JE, Ouyang H, Strayhorn CL, Windle JJ, Somerman MJ. Employing a transgenic animal model to obtain cementoblasts in vitro. J Periodontol. 2000;71:63-72.
D'Errico JA, MacNeil RL, Takata T, Berry J, Strayhorn C, Somerman MJ. Expression of bone associated markers by tooth root lining cells, in situ and in vitro. Bone. 1997;20:117- 26.
D’Souza RN, Aberg T, Gaikwad J, Cavender A, Owen M, Karsenty G, Thesleff I. Cbfa1 is required for epithelial- mesenchymal interactions regulating tooth development in mice. Development. 1999;126:2911-20.
Franceschi RT, Xiao G. Regulation of the osteoblast-specific transcription factor, Runx2: responsiveness to multiple signal transduction pathways. J Cell Biochem. 2003;88:446-54.
Hirata A, Sugahara T, Nakamura H. Localization of runx2, osterix, and osteopontin in tooth root formation in rat molars. J Histochem Cytochem. 2009;57:397-403.
Jiang H, Sodek J, Karsenty G, Thomas H, Ranly D, Chen J.
Expression of core binding factor Osf2/Cbfa-1 and bone sialoprotein in tooth development. Mech Dev. 1999;81:169- 73.
Kim Mi-Ri, Yang Won-Kyung, Grzesik Wojciech, Ko Hyun- Jung. Inhibition of osteoclast formation by putative human cementoblasts. International Journal of Oral Biology. 2008;
33:113-6.
Kitagawa M, Tahara H, Kitagawa S, Oka H, Kudo Y, Sato S, Ogawa I, Miyaichi M, Takata T.Characterization of established cementoblast-like cell lines from human cementum-lining cells in vitro and in vivo. Bone. 2006;39:1035-42.
Ko Hyun-Jung, Grzesik Wojciech J. BMP Expression by Human Cementum-Derived Cells in vitro. International Journal of Oral Biology. 2005;30:99-103.
Koga T, Matsui Y, Asagiri M, Kodama T, de Crombrugghe B, Nakashima K, Takayanagi H. NFAT and Osterix cooperatively regulate bone formation. Nat Med. 2005;11:880-5.
Liu W, Toyosawa S, Furuichi T, Kanatani N, Yoshida C, Liu Y, Himeno M, Narai S, Yamaguchi A, Komori T. Over- expression of Cbfa1 in osteoblasts inhibits osteoblast maturation and causes osteopenia with multiple fractures. J Cell Biol. 2001;155:157-66.
Maruyama Z, Yoshida CA, Furuichi T, Amizuka N, Ito M, Fukuyama R, Miyazaki T, Kitaura H, Nakamura K, Fujita T, Kanatani N, Moriishi T, Yamana K, Liu W, Kawaguchi H, Nakamura K, Komori T. Runx2 determines bone maturity and turnover rate in postnatal bone development and is
involved in bone loss in estrogen deficiency. Develop Dynamic. 2007;236:1876-90.
Matsubara T, Kida K, Yamaguchi A, Hata K, Ichida F, Meguro H, Aburatani H, Nishimura R, Yoneda T.BMP2 regulates Osterix through Msx2 and Runx2 during osteoblast differentiation. J Biol Chem. 2008;283:29119-25.
Nakashima K, Zhou X, Kunkel G, Zhang Z, Deng JM, Behringer RR, de Crombrugghe B. The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone formation. Cell. 2002;108:17-29.
Nanci, A. Ten Cate's Oral histology: Development, structure, and function. pp 239-267, Mosby, 2008.
Nishio Y, Dong Y, Paris M, O'Keefe RJ, Schwarz EM, Drissi H. Runx2-mediated regulation of the zinc finger Osterix/
Sp7 gene. Gene. 2006;372:62-70.
Saito Y, Yoshizawa T, Takizawa F, Ikegame M, Ishibashi O, Okuda K, Hara K, Ishibashi K, Obinata M, Kawashima H. A cell line with characteristics of the periodontal ligament fibroblasts is negatively regulated for mineralization and Runx2/Cbfa1/Osf2 activity, part of which can be overcome by bone morphogenetic protein-2. J Cell Sci. 2002;115:4191-200.
Saygin NE, Giannobile WV, Somerman MJ. Molecular and cell biology of cementum. Periodontol 2000. 2000;24:73-98.
Tai G, Christodoulou I, Bishop AE, Polak JM. Use of green fluorescent fusion protein to track activation of the tran- scription factor osterix during early osteoblast differentiation.
Biochem Biophys Res Commun. 2005;333:1116-22.