Spore Associated Bacteria (SAB) of Arbuscular Mycorrhizal Fungi (AMF) and Plant Growth Promoting Rhizobacteria (PGPR) Increase Nutrient Uptake and Plant Growth Under Stress Conditions
11
0
0
전체 글
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
수치


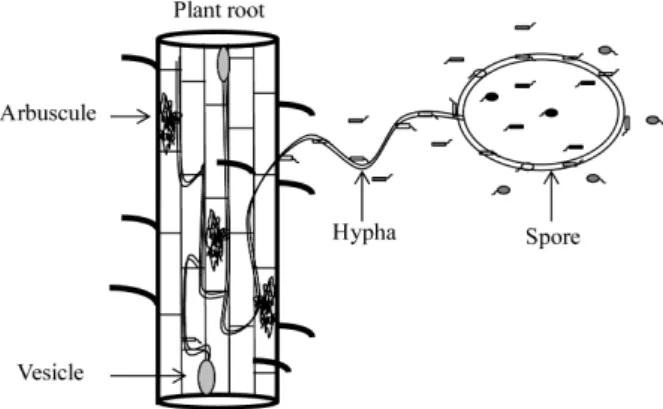
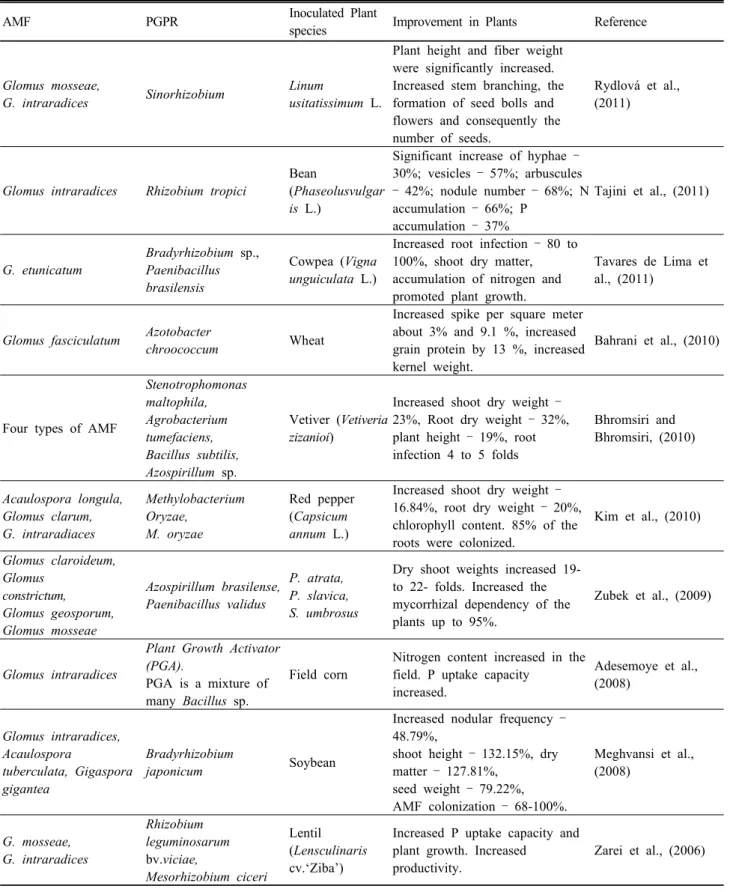
+2
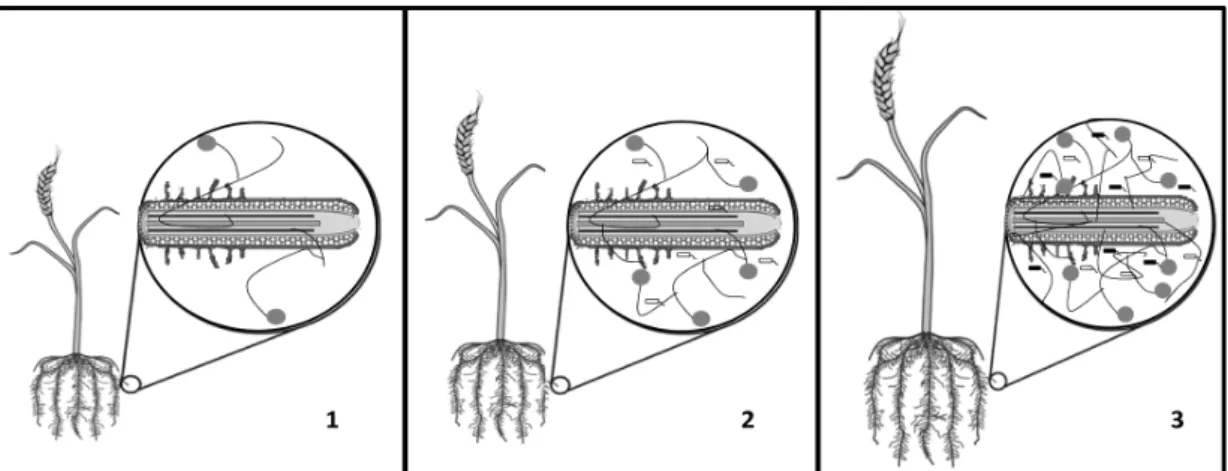
관련 문서
Characterization of Heavy Metal Tolerant and Plant Growth-Promoting Rhizobacteria Isolated from Soil Contaminated with Heavy Metal and Diesel.. Soo Yeon Lee, Yun-Yeong
The objective of this study was to determine the effect of electrical conductivity (EC) of nutrient solution on the growth, nutrient uptake of potted kalanchoe
ABSTRACT The aim of our study was to investigate the interactive effects of container size and nutrient supply on plant growth, chlorophyll synthesis, transpiration, CO
Canola Plant Growth Promotion by a Selected Plant Growth Promoting- Rhizobacteria, Burkholderia pyrrocinia Strain 13-1 in the Cold Condition.. Jae Eun Lee, Sang Min Cho, Young
Isolation and Characterization of Plant Growth Promoting Rhizobacteria From Button Mushroom Compost.. Korean Journal of
