pISSN 1598-642X eISSN 2234-7305
미생물제제시용 고추경작지로부터 식물생장홀몬과 항진균물질을 동시에 생산하는 식물생장촉진근권세균의 선발 및 동정
정병권·임종희·안창환·김요환·김상달*
영남대학교 미생물생명공학과
Received : July 3, 2012 / Accepted : September 6, 2012
Selection and Identification of Phytohormones and Antifungal Substances Simultaneously Producing Plant Growth Promoting Rhizobacteria from Microbial Agent Treated Red-pepper Fields. Jung, Byung- Kwon, Jong-Hui Lim, Chang-Hwan An, Yo-Hwan Kim, and Sang-Dal Kim*. Department of Applied Microbiology and Biotechnology, School of Biotechnology, Yeungnam University, Gyeongsan 712-749, Korea − In this study, a total of more than 1,000 bacteria, including 739 species of aerobic bacteria, 80 species of ure- ase producing bacteria and 303 species of photosynthetic bacteria, were isolated from red-pepper field soils located in the Gyeongsan Province of the Republic of Korea. Amongst these, 158 species of aerobic bacteria, 70 species of urease producing bacteria and 228 species of photosynthetic bacteria were found to be auxin pro- ducing soil bacteria through quantification analysis with the Salkowski test. The latter groupings were then tested for antifungal activities to β-Glucanase and siderophore using CMC congo red agar and CAS blue agar media. In addition, the selected strains were examined for antifungal activity against various phytopathogenic fungi on PDN agar media. Six strains; BCB14, BCB17, C10, HA46, HA143, and HJ5, were noted for their ability to both produce auxin and act as antifungal substances. 16S rDNA sequence comparison analyses of these six strains identified them as Bacillus subtilis BCB14, B. methylotrophicus BCB17, B. methylotrophicus C10, B. sonorensis HA46, B. subtilis HA143, and B. safensis HJ5.
Keywords: Microbial agent, phytohormone, PGPR, antifungal activity
서 론
현대농업은 경작면적에 비해 상대적으로 많은 노동력과 자본을 이용하는 경작방식인 집약농업이 대부분 이루어지고 있으며, 작물 재배 시 수율의 향상을 위해 화학비료 및 화학 농약이 광범위하게 사용되고 있다. 하지만 비용이 많이 소 모되고 병해충의 내성과 잔류독성과 같은 많은 문제점들을 발생시키고 있다. 때문에 최근에는 환경친화적인 유기농업 에 대한 관심이 집중되고 있으며, 식물병해 방제와 작물의 성장을 촉진시키는 생물농약이나 미생물제제를 개발하기 위 한 연구가 활발히 진행되고 있다[9, 13].
지난 20년 동안 전 세계적으로 친환경 농업을 위해 Plant growth promoting rhizobacteria (PGPR)를 이용한 미생물비 료와 생물농약의 사용이 꾸준히 증가하였으며[14], 그 예로 Chen과 Amara에 의해 고추와 같이 경제적으로 중요한 작물 의 근권에 PGPR을 처리할 경우 작물의 생장과 수율이 향상
된다는 것이 보고되어 왔다[1, 6].
PGPR은 식물의 근권에 서식하는 미생물로 근권 군서 (Rhizosphere community)를 이루고 있으며 다양한 특성을 가지는 화학물질을 생산하여 식물의 생장이나 영양분 흡수 를 촉진시키거나, 면역성 증대와 식물병원성 진균의 길항작 용을 통해 질병의 감염을 억제시키는 기능을 가진다[12, 22].
이들이 생산하는 유용한 물질들 중에는 식물의 생장에 직접 적으로 관여하는 것으로 auxin, gibberellin 등의 식물호르몬 이 있다. Auxin은 식물의 세포신장, 발아, 기관의 분화, 신 호전달 등에 관여하는 식물생장조절물질 중 하나로 1930년 대 Thianmann과 Link는 Rhizobium에 의하여 auxin이 유도 될 수 있다는 것을 보고 하였으며 그 후 미생물에 의해서 많 은 유도체들이 생성될 수 있음이 보고되어 왔다[23]. 다른 식물생장조절물질인 gibberellin은 줄기 및 세포신장, 휴면타 파, 종자발아촉진, 개화촉진 등에 관여하며[4] Gibberella fujikuroi와 Aspergillus niger 등에 의해 생산되는 것으로 알 려져 있다[2]. 또한 PGPR은 식물에 필요한 영양분의 공급 을 위해 유용한 효소 및 생리활성물질을 생산하는데 그 물 질로는 토양에 존재하는 urea를 가수분해하여 생장에 필요 한 질소원을 공급하는 urease [7] 등의 효소와 광합성미생물
*Corresponding author
Tel: +82-53-810-3053, Fax: +82-53-810-4663 E-mail: [email protected]
에 의해 생산되는 비타민, 아미노산 등 생리활성물질이 있 다[24]. 또한 β-Glucanase와 같은 효소를 생산하여 식물병원 성 진균의 세포외벽을 가수분해하거나 토양에 존재하는 철 이온(Fe3+)과 결합하는 siderophore를 생산하여 식물병원성 진균의 포자발아를 억제시키는 것과 같이 간접적으로 식물 의 생장에 큰 영향을 끼치고 있다[11].
본 연구에서는 Bacillus 속 2균주와 Pseudomonas 속 1균 주로 조성된 미생물제제를 1년간 시용한 고추경작지로부터 auxin 생산성을 가지는 근권미생물을 분리하여 식물호르몬 을 생산함과 동시에 식물병원성 진균의 생장을 억제시키는 길항물질을 생산하는 토양 미생물을 선발, 동정하여 향후 미 생물제제로서의 이용 가능성을 알아보고자 하였다.
재료 및 방법 고추 경작지로부터 균주의 분리
식물생장촉진 호르몬인 auxin을 생산하는 균주를 분리하 기 위해 혼합제의 미생물제제를 1년간 시용한 경북 경산시 대동소재 시험포장에서 고추뿌리에 부착되어있는 고추 근권 토양 0.1 g을 멸균 증류수 0.9 mL에 첨가한 후 단계적으로 희석하여 Nutrient agar (beef extract 0.3%, peptone 0.5%, agar 1.5%) 배지에 0.1 mL씩 평판도말 하였으며, 기능성 세 균의 분리를 위해 urease 생산 균주 분리 배지인 Urea agar base (peptone 0.1%, dextrose 0.1%, NaCl 0.5%, KH2PO4 0.2%, urea 2% phenol red 0.001%, agar 1.5%) 배지와 광 합성 균주 분리 배지인 Athiorhodaceae base agar (NH4Cl 0.1%, NaHCO3 0.1%, K2HPO4 0.02%, CH3COONa 0.3%, MgSO4·7H2O 0.02%, NaCl 0.1%, yeast extract 1%, trace element 1%, agar 2%) 배지[10]에 0.1 mL씩 평판도말 하여 30oC에서 24~48시간 배양하였다.
Auxin 생산 균주의 선발
Auxin 생산 균주의 선발을 위하여 L-tryptophan을 0.1%
첨가한 King’s B broth 배지에 1 loop씩 접종하여 30oC에서 24시간 배양한 후 8,000 rpm에서 10분간 원심분리하여 상등 액을 분리한 뒤 Salkowski reagent (5% perchloric acid 100 mL, 0.05 M ferric chloride 2 mL)를 1:2(v/v)로 혼합하 고 암조건, 실온에서 30분간 반응시킨 후 색상이 분홍색으 로 전환된 것을 auxin 생성능이 있는 것으로 결정하였으며 [28], 반응물을 spectrophotometer를 이용하여 535 nm 파장 에서 측정하여 정량하였다. 표준곡선은 IAA (Sigma, USA) 를 이용하여 작성하였으며, auxin의 생산성을 가지는 균주 를 선발하였다. 또한 선발된 균주의 gibberellin의 생산량은 Holbrook 등의 방법[21]을 사용하여 gibberellic acid을 gibberellenic acid로 전환시킨 뒤 spectrophotometer로 254 nm 파장에서 측정하였으며, 표준곡선은 GA3 (Sigma, USA) 를 이용하여 작성하였다.
Auxin 생산 균주의 β-Glucanase와 siderphore 생산성 조사 선발된 auxin 생산성 균주의 β-Glucanase와 siderophore의 생산성을 확인하기 위해 각각의 선별배지를 사용하였다. β- Glucanase 생산능은 1% carboxymethyl-cellulose (CMC)를 함유한 Plate count agar (pancreatic digest of casein 0.5%, yeast extract 0.25%, dextrose 0.1%, agar 15%) 배지에 선 발된 균주를 toothpicking하여 30oC에서 2일간 배양한 후 Congo red plate 방법을 이용하여 색깔의 변화를 통해 β- Glucanase의 생산을 확인하였으며[27], siderophore의 생산 확인은 CAS (chrome azurol S)가 포함된 CAS blue agar 배지에 선발된 균주를 toothpicking하여 30oC에서 4일간 배 양한 후 orange halo zone의 형성에 따라 생산 유무를 확인 하였다[19].
Auxin 생산 균주의 식물병원성 진균에 대한 길항범위 조사 식물생장촉진 호르몬인 auxin과 식물병원성 진균의 생육 을 억제하는 항진균물질 생산 균주의 방제력은 대표적 식물 병원성 진균 6종 Fusarium oxysporum [17], Phytophthora capsici [29], Corynespora cassiicola [3], Rhizoctonia solani AG-1 (IA), R. solani AG-1 (IB) [18], Pythium ultimum [5]을 대상으로 하여 대치배양을 실시한 뒤 생육저지대 측정 법을 통해 조사하였으며[15], 사용균주는 농촌진흥청 농업유 전자원정보센터 (KACC)로부터 분양받아 사용하였다(Table 1). 선발된 균주 및 병원성 진균의 대치배양은 PDN agar (Potato starch 0.4%, dextrose 2%, nutrient broth 0.8%, agar 1.5%) 배지를 사용하였으며, 배지 왼쪽에 병원성 진균 을 2×2 cm 크기의 균체 disc로 접종하고 3 cm 떨어진 곳에 선발 균주를 이용하여 획선을 그은 뒤, 30oC에서 3~5일간 배양하여 생육저지대 형성을 관찰한 후 다양한 길항범위를 가지는 균주를 최종 선발하였다.
균주의 동정
최종 선발된 균주의 분류학적 동정을 위하여 Solgent Co., Ltd. Korea에 의뢰하였으며, 16S-27F (5’-AGA GTT TGA TCC TGG CTC AG-3’)와 16S-1492R (5’-GGT TAC CTT GTT ACG ACT T-3’)을 이용하여 선발된 균주의 16S rDNA 의 염기서열을 분석하였다. 분석된 염기서열은 NCBI의 Basic Table 1. Phytopathogenic fungal strains used in dual culture test.
Strains KACC No. Plant diseases Fusarium oxysporum f. sp.
Lycopersici
40032 Fusarium wilt Rhizoctonia solani AG-1 (IA) 40101 Sheath blight Rhizoctonia solani AG-1 (IB) 40111 Damping-off Phytophthora capsici 40476 Blight
Pythium ultimum 40705 Stem rot, Damping-off Corynespora cassicola 40964 Leaf blight, Brown spot
Local Alignment Search Tool (BLAST) algorithm을 통해 GenBank database에 등록된 염기서열과 비교하여 상동성을 조사한 후 서울대학교 EzTaxon server (ver. 2.1) [8]와 Molecular Evolutionary Genetics Analysis (MEGA) version 4 software [26]를 사용하여 계통도를 조사한 다음 최종 동 정하였다.
결과 및 고찰 Auxin 생산 균주의 선발
미생물제제 시용 고추경작지 근권토양을 단계희석하여 Nutrient agar 배지에 배양한 결과 739종의 일반호기성 균주 를 분리할 수 있었다[16, 20]. 또한 urease 생산 균주 80종 및 광합성 균주 303종을 분리할 수 있었으며[25], 분리된 균 주들의 auxin 생산능을 확인하기 위해 배양 후 원심분리하 여 분리한 상등액을 Salkowski reagent와 1:2(v/v)로 혼합하 여 반응시켰다. 그 후 분홍색으로 발색반응을 보이는 반응 액을 선별하여 spectrophotometer를 이용해 535 nm 파장에 서 측정하여 정량분석한 결과, auxin을 생산하는 158종의 일 반호기성 균주와 70종의 urease 생산균주 및 228종의 광합 성 균주를 선발할 수 있었다(Table 2). 선발된 균주는 auxin 을 생산할 뿐만 아니라 urea를 ammonia로 가수분해하여 식 물에 질소원을 공급하고, 광합성을 통해 황화수소와 같은 유 해가스를 기질로하여 각종 생리활성물질, 유용물질을 생합 성함으로써 식물의 생장을 촉진할 수 있을 것이다.
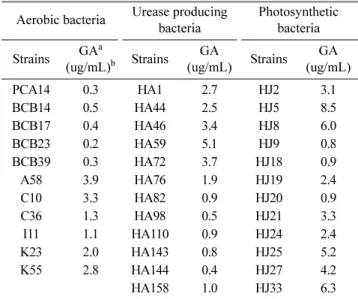
Auxin 생산 균주의 gibberellin 생산성 조사
선발된 균주의 gibberellin의 생산성은 Holbrook 등의 방 법[21]을 통해 조사하였으며, 그 결과 대부분의 주요 auxin
생산 균주에서 gibberellin을 생산한다는 것을 확인할 수 있 었다(Table 3). 이와 같은 결과를 근거로 하여 선발된 균주 를 고추와 같은 작물의 근권에 처리하였을 경우 auxin과 gibberellin의 동시생산을 통해 식물의 세포신장, 발아, 기관 의 분화, 종자발아촉진 및 개화촉진을 유도함으로 인해 미 생물 제제로써 이용할만한 가능성이 있다고 생각된다.
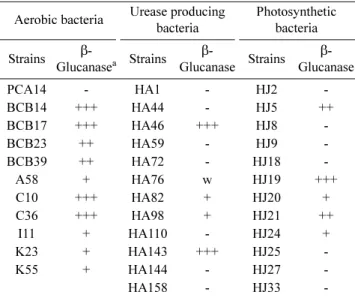
선발된 auxin 생산 균주의 β-Glucanase 및 siderophore 생산성 조사
Auxin 생산 균주의 β-Glucanase 생산유무 확인을 위해 Congo red plate 법을 이용하여 기질인 CMC가 분해되어 형 성되는 색깔 변화에 의해 조사하였다. 그 결과, 일반호기성 균주 117종, urease 생산 균주 7종 및 광합성 균주 38종에 서 β-Glucanase를 생산하는 것을 확인할 수 있었다(Table 4, Fig. 1). 또 다른 항진균 물질인 siderophore의 생산성은 CAS blue agar 배지에 선발된 auxin 생산균주를 toothpick 하여 orange halo zone의 형성유무를 통해 조사하였으며, 호 기성균주 45종, urease 생산균주 12종 및 광합성 균주 188 종에서 siderophore를 생산하는 것을 확인할 수 있었다(Table 5, Fig. 2). 이는 세포벽이 cellulose와 β-Glucan으로 이루어 져있는 식물병원성 진균의 세포벽을 분해하고, siderophore 의 생산으로 식물병원성 진균과의 철흡수 경쟁에서 우위를 점함으로써 방제력을 나타낼 수 있을 것이다.
항진균 물질을 생산하는 선발균주의 식물병원성 진균에 대 한 길항범위 조사
선발된 auxin 생산균주 중에서 다양한 항진균물질을 생산 Table 2. Selection of auxin producing PGPR by Salkowski test.
Aerobic bacteria Urease producing bacteria
Photosynthetic bacteria Strains Auxin
(ug/mL)a Strains Auxin
(ug/mL) Strains Auxin (ug/mL)
PCA14 12.1 HA1 12.2 HJ2 1.8
BCB14 17.8 HA44 9.2 HJ5 12.6
BCB17 15.6 HA46 18.3 HJ8 3.4
BCB23 3.1 HA59 7.7 HJ9 2.3
BCB39 1.2 HA72 2.5 HJ18 2.5
A58 3.9 HA76 9.8 HJ19 1.9
C10 13.3 HA82 6.5 HJ20 3.4
C36 1.3 HA98 4.0 HJ21 1.9
I11 1.1 HA110 1.6 HJ24 1.5
K23 2.0 HA143 19.0 HJ25 3.3
K55 2.8 HA144 3.3 HJ27 1.5
HA158 2.5 HJ33 1.3
aAuxin production had been estimated absorbance at 535 nm by Salkowski test with Indol-3-acetic acid strandard calibration curve.
Table 3. Gibberellin simultaneous production of auxin producing PGPR.
Aerobic bacteria Urease producing bacteria
Photosynthetic bacteria Strains GAa
(ug/mL)b Strains GA
(ug/mL) Strains GA (ug/mL)
PCA14 0.3 HA1 2.7 HJ2 3.1
BCB14 0.5 HA44 2.5 HJ5 8.5
BCB17 0.4 HA46 3.4 HJ8 6.0
BCB23 0.2 HA59 5.1 HJ9 0.8
BCB39 0.3 HA72 3.7 HJ18 0.9
A58 3.9 HA76 1.9 HJ19 2.4
C10 3.3 HA82 0.9 HJ20 0.9
C36 1.3 HA98 0.5 HJ21 3.3
I11 1.1 HA110 0.9 HJ24 2.4
K23 2.0 HA143 0.8 HJ25 5.2
K55 2.8 HA144 0.4 HJ27 4.2
HA158 1.0 HJ33 6.3
aGA: Gibberellic acid
bGibberellic acid productivity was estimated absorbance 254 nm by Holbrook test with GA3 strandard calibration curve.
하는 균주를 이용하여 6종의 식물병원성 진균에 대치배양을 실시한 후 생육저지대 측정법을 이용하여 병원성 진균에 대 한 길항범위를 조사하였다. 그 결과 BCB17, C10, HA46,
HA143 균주가 식물병원성 진균 6종의 생육을 억제하는 것 으로 조사되었다. 또한 BCB14 균주는 F. oxysporum 균주를 제외한 식물병원성진균 5종에 대하여 생육을 억제하였으며, 광합성능을 가지는 HJ5 균주는 R. solani AG-1 (IA)와 P.
Table 4. Simultaneous production of β-Glucanase among auxin producing PGPR.
Aerobic bacteria Urease producing bacteria
Photosynthetic bacteria Strains β-
Glucanasea Strains β-
Glucanase Strains β- Glucanase
PCA14 - HA1 - HJ2 -
BCB14 +++ HA44 - HJ5 ++
BCB17 +++ HA46 +++ HJ8 -
BCB23 ++ HA59 - HJ9 -
BCB39 ++ HA72 - HJ18 -
A58 + HA76 w HJ19 +++
C10 +++ HA82 + HJ20 +
C36 +++ HA98 + HJ21 ++
I11 + HA110 - HJ24 +
K23 + HA143 +++ HJ25 -
K55 + HA144 - HJ27 -
HA158 - HJ33 -
+++: >3 mm, ++: >2 mm, <3 mm, +: >1 mm, <2 mm, w: <1 mm, -:
negative.
aβ-Glucanase production detected by color change on CMC-Congo red agar medium.
Fig. 1. Detection of β-Glucanase activity by auxin producing PGPR on CMC-Congo red agar medium. P: Bacillus subtilis AH18(siderophore producing strain); N: Escherichia coli ATCC 28922(siderophore non-producing strain); A: Strain BCB14; B:
Strain BCB17; C: Strain C10; D: Strain HJ5; E: Strain HA46; F:
Strain HA143.
Table 5. Simultaneous production of siderophore among auxin producing PGPR.
Aerobic bacteria Urease producing bacteria
Photosynthetic bacteria Strains Siderophorea Strains Siderophore Strains Siderophore
PCA14 + HA1 + HJ2 +++
BCB14 ++ HA44 + HJ5 +++
BCB17 ++ HA46 +++ HJ8 ++
BCB23 + HA59 + HJ9 ++
BCB39 + HA72 - HJ18 +++
A58 - HA76 + HJ19 +
C10 + HA82 - HJ20 +
C36 + HA98 w HJ21 +
I11 + HA110 + HJ24 +
K23 w HA143 + HJ25 +++
K55 - HA144 + HJ27 +++
HA158 ++ HJ33 +++
+++: >3 mm, ++: >2 mm, <3 mm, +: >1 mm, <2 mm, w: <1 mm, -:
negative.
aSiderophore production detected by orange halo zone on CAS blue agar medium.
Fig. 2. Detection of siderophore producing strains among auxin producing PGPR on CAS blue agar medium. P: Bacillus subti- lis AH18(siderophore producing strain); N: Escherichia coli ATCC 28922(siderophore non-producing strain); A: Strain BCB14; B:
Strain BCB17; C: Strain C10; D: Strain HA46; E: Strain HA146;
F: Strain HJ5.
ultimum, P. capsici 균주에 대하여 강한 생육억제능을 가지 는 것으로 조사되었다. 따라서 auxin을 생산함과 동시에 다 양한 식물병원성 진균에 대해 길항범위를 가지는 BCB14, BCB17, C10, HA46, HA143, HJ5 균주를 최종 선발하였다 (Table 6, Fig. 3).
선발된 다기능 길항균주의 동정
최종 선발된 auxin 생산 및 항진균성 물질을 동시에 생산 하며 다양한 식물병원성 진균에 대해 길항범위를 갖는 균주 의 16S rDNA 염기서열을 분석하였다. 분석한 염기서열은 NCBI의 BLAST search를 이용한 기존의 세균과 유사성을 비교한 결과 98% 이상의 상동성을 나타내었으며, EzTaxon
server (ver. 2.1)와 MEGA version 4 software를 사용하여 계통분석을 실시한 후 최종적으로 B. subtilis BCB14, B.
methylotrophicus BCB17, B. methylotrophicus C10, B.
Table 6. Antifungal spectrum of auxin producing PGPR against various phytopathogenic fungi.
Type of bacteria Strains
Phytopathogenic fungi R.* Solani AG-1
(IA)
R. Solani AG-1
(IB) F.** oxysporum P. ultimum P. capsici C. cassiicola
Aerobic bacteria
BCB14 + a +++ - + ++ ++++
BCB17 + ++ +++ ++ ++++ ++
C10 + + +++ + +++ ++
Urease producing bacteria
HA46 + + +++ ++ + +++
HA143 ++ + ++ +++ ++ +++
Photosynthetic bacteria HJ5 ++ - - +++ ++++ -
++++: >4 mm, +++: >3 mm, <4 mm ++: >2 mm, <3 mm, +: >1 mm, <2 mm, -: negative.
aAntifungal activity test by dual-culturing between selected strain and phytopathogenic fungi on PDN agar medium.
*R: Rhizoctonia
**F: Fusarium
Fig. 3. Antifungal activity against various phytopathogenic fungi of the selected strains on PDN agar medium. A: R. solani AG- 1(IA)(L) and Strain C10(R), B: R. solani AG-1(IB) (L) and Strain BCB14(R), C: F. oxysporum (L) and Strain HA143(R), D: P. ultimum (L) and Strain HA143(R), E: P. capsici (L) and Strain HJ5(R), F: C. cassiicola (L) and Strain HA46(R).
Table 7. Identification of auxin and antifungal substances pro- ducing PGPR.
Strains Identification Similarity (%)
BCB14 Bacillus subtilis 99.7
BCB17 Bacillus methylotrophicus 99.5 C10 Bacillus methylotrophicus 99.7
HA46 Bacillus sonorensis 98.9
HA143 Bacillus subtilis 99.9
HJ5 Bacillus safensis 99.8
sonorensis HA46, B. subtilis HA143, B. safensis HJ5으로 동정 및 명명하였다(Table 7, Fig. 4).
요 약
식물생장 촉진 홀몬 auxin을 생산하는 균주를 선발하기 위해 경북 경산시 소재 고추경작지 근권토양으로 부터 739 종의 일반호기성 균주와 urease 생산 균주 80종 및 광합성 균주 303종과 같이 총 1000여종 이상의 균주를 분리하였다.
이 균주들을 대상으로 Salkowski test를 실시한 결과, auxin 을 생산하는 158종의 일반호기성 균주와 70종의 urease 생 산 균주 및 228종의 광합성 균주를 선발할 수 있었으며, Holbrook test를 통해 또 다른 식물생장 촉진 호르몬인 gibberellin도 대부분의 균주에서 생산되는 것을 확인할 수 있었다. 선발된 균주 중 항진균 물질인 β-Glucanase와 siderophore를 생산하고 다양한 병원성 진균에 대해 길항 범 위를 가지는 6가지 균주 BCB14, BCB17, C10, HA46, HA143, HJ5를 toothpicking 및 대치배양을 통해 최종 선발 할 수 있었으며, 분류학적으로 동정한 결과 6종 모두 B.
subtilis BCB14, B. methylotrophicus BCB17, B. methylo- trophicus C10, B. sonorensis HA46, B. subtilis HA143, B. safensis HJ5로 확인되었다.
ACKNOWLEDGEMENTS
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (NRF-2012-0001795)
REFERENCES
1. Amara, M. A. T. and M. S. A. Dahdoh. 1997. Effect of inoculation with plant growth promoting rhizobacteria (PGPR) on yield and uptake of nutrients by wheat grown on sandy soil. Egyptian J. Soil Science 37: 467-484.
2. Ates, S., S. Ozenir, and M. Gökdere. 2006. Effect of silicone oil on gibberellic acid production by Gibberella fujikuroi and Aspergillus niger. Appl. Biochem. Microbiol. 42: 500-501.
3. Barthe, P., V. Pujade-Renaud, F. Breton, D. Gargani, R. Thai, and C. Roumestand. 2007. Structural analysis of cassiicolin, a host-selective protein toxin from Corynespora cassiicola. J.
Mol. Biol. 367: 89-101.
4. Bomke, C. and B. Tudzynski. 2009. Diversity, regulation, and evolution of the gibberellin biosynthetic pathway in fungi compared to plants and bacteria. Phytochemistry 70:
1876-1893.
5. Chen, W., H. A. J. Hoitink, A. F. Schmitthenner, and O. H.
Tuovinen. 1988. The role of microbial activity in suppression of daming-off caused by Pythium ultimum. Phytopathology 78: 314-322.
6. Chen, Y., R. Mei, S. Lu, L. Liu, and J. W. Kloepper. 1994.
The use of yield increasing bacteria (YIB) as plant growth- promoting rhizobacteria in chinese agriculture, Utkhede, R.
S. and V. K. Gupta (ed), Management of Soil Borne Diseases, Kalyani Publishers, New Delhi, pp. 165-184.
7. Choi, J. G., Y. S. Kim, W. T. Lee, and S. D. Kim. 1997.
Urease gene transfer of antagonistic bacillus subtilis YB-70 and increased antagonistic effect. Kor. J. Microbiol. Bio- technol., 25: 30-36.
8. Chun, J. S., J. H. Lee, Y. Y. Jung, M. J. Kim, S. Kim, B. K.
Kim, and Y. W. Lim. 2007. EzTaxon: a web-based tool for the identification of prokaryotes based on 16S ribosomal RNA gene sequences. Int. J. Syst. Evol. Microbiol. 57: 2259- Fig. 4. Phylogenetic tree of 16S rDNA sequence of selected auxin producing PGPR. The branching pattern of tree was produced by neighbour-joining method with MEGA version 4 software.
2261.
9. Esitken, A., S. Ercisli, H. Karlidag, and F. Sahin. 2005.
Potential use of plant growth promoting rhizobacteria (PGPR) in organic apricot production. Proc. Int. Sci. Conf. Environ- mentally Friendly Fruit Growing Tartu-Estonia, September 7-9, pp. 90-97.
10. Imhoff, J. F., and H. G. Truper. 1989. Purple nonsulfur bacteria. pp. 1658-1662. In J. T. Staley et al. (ed.) Bergey’s manual of systematic bacteriology. Vol. 3. Williams and Wilkins, Baltimore.
11. Ishimaru, C. A. and J. E. Loper. 1992. High-affinity iron uptake systems present in Erwinia carotovora subsp.
carotovora include the hydroxamate siderophore aerobactin.
J. bacteriol. 174: 2993-3003.
12. Jung, H. K., J. R. Kim, S. M. Woo, and S. D. Kim. 2006. An auxin producing plant growth promoting rhizobacterium Bacillus subtilis AH18 which has siderophore-producing biocontol activity. J. Kor. Soc. Appl. Biol. Chem. 7: 94-100.
13. Jung, H. K., J. R. Kim, S. M. Woo, and S. D. Kim. 2007.
Selection of the auxin, siderophore and cellulase- producing PGPR, Bacillus licheniformis K11 and its plant growth promoting mechanisms. J. Kor. Soc. Appl. Biol. Chem. 50:
23-28.
14. Khalid, A., M. Arshad, and Z. A. Zahir. 2004. Screening plant growth-promoting rhizobacteria for improving growth and yield of wheat. J. Appl. Microbiol. 96: 473-480.
15. Kim, B. S., J. W. A, and K. Y. Cho. 1998. Isolation and identification of Myxobacteria KR025 and searching of their bioactive compounds. Kor. J. Plant Pathol. 14: 345-349.
16. Kim, J. W., B. K. Jung, C. H. An, J. H. Lim, and S. D. Kim.
2011. Isolation of cellulase-producing plant growth promot- ing rhizobacteria in pepper farming soil. Yeungnam University.
17. Kistler, H. C. and U. K. Benny. 1988. Genetic transforma- tion of the fungal plant wilt pathogen, Fusarium oxysporum.
Curr. Genet. 13: 145-149.
18. Kousik, C. S., J. P. Snow, and R. A. Valverde. 1993. Com- parison of double-stranded RNA components and virulence among isolates of Rhizoctonia solani AG-1 IA and AG-1 IB.
Phytopathology 84: 44-49.
19. Lee, J. M., H. S. Lim, T. H. Chang, and S. D. Kim. 1999.
Isolation of siderophore-producing Pseudomonas fluorescens GL7 and its biocontrol activity against root-rot disease. Kor.
J. Appl. Microbiol. Biotechnol. 27: 427-432.
20. Lee, K. E., B. K. Jung, C. H. An, J. H. Lim, and S. D. Kim.
2011. Isolation and identification of siderophore-producing bacteria that has antifungal activity. Yeungnam University.
21. Gutierrez-Manero, F. J., B. Ramos-Solano, A. Probanza, J.
Mehouachi, F. R. Tadeo, and M. Talon. 2001. The plant growth promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiol. Plant. 111: 206-211.
22. Loon, L. C. van. 2007. Plant responses to plant growth- promoting rhizobacteria. Eur. J. Plant Pathol. 119: 243-254.
23. Lumsden, R. D. 1981. In The Fungal Community: Ecology of mycoparasitism. D. T. Carroll (ed.), Marcel Dekker Inc, N. Y. pp. 295-328.
24. Moon, S. W., M. Matsuyama, G. M. Go, Y. D. Lee, and Y. B.
Go. 1997. N2 fixation and H2 production by a phototrophic bacterium, Chromatium sp. Kor. J. Aquaculture 10: 69-76.
25. Shon, H. J., B. K. Jung, Y. H. Kim, J. H. Lim, and S. D. Kim.
2011. Isolation and identification of auxin and gibberellin producing photosynthetic bacteria in pepper farming. Yeung- nam University.
26. Tamura, K., J. Dudley, M. Nei, and Sudhir Kumar. 2007.
MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) Software Version 4.0. Mol. Biol. Evol. 24: 1596-1599.
27. Teather, R. and P. J. Wood. 1982. Use of congo red-polysac- charide interations in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl. Environ.
Microbiol. 43: 777-780.
28. Thomas, O., G. D. Dieter, and H. Dieter. 1991. IAA synthesis in the biocontrol strain CHO of Pseudomonas fluorescens:
role of tryptophan side chain oxidase. J. Gen. Microbiol.
137: 2273-2279.
29. Xu, J., X. Zhao, X. Han, and Y. Du. 2007. Antifungal activity of oligochitosan against Phytophthora capsici and other plant pathogenic fungi in vitro. Pesticide Biochem. Physiol.
87: 220-228.