Current Perspectives on the Effects of Plant Growth-promoting Rhizobacteria
Thien Tu Huynh Le
1†, Sang Eun Jun
2†and Gyung-Tae Kim
1,2*
1
Department of Applied Bioscience, Graduate School of Natural Science, Dong-A University, Busan 49315, Korea
2
Department of Molecular Genetics, College of Natural Resources and Life Science, Dong-A University, Busan 49315, Korea
Received November 6, 2019 /Revised November 19, 2019 /Accepted November 21, 2019
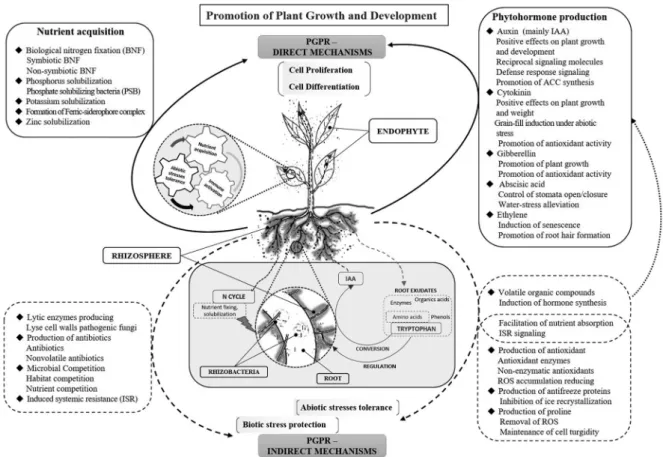
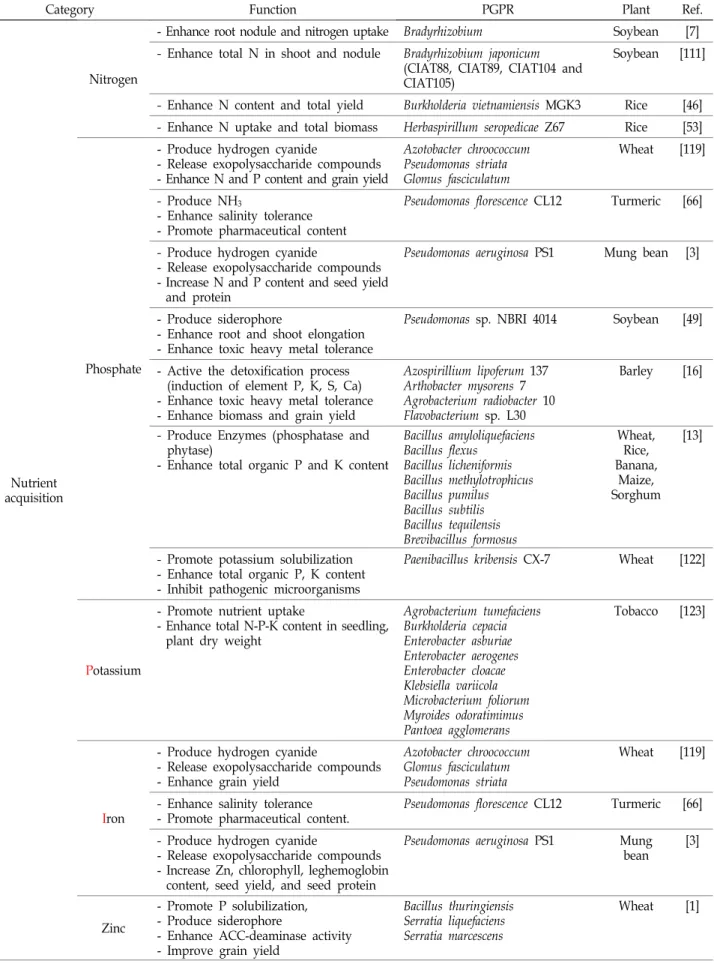
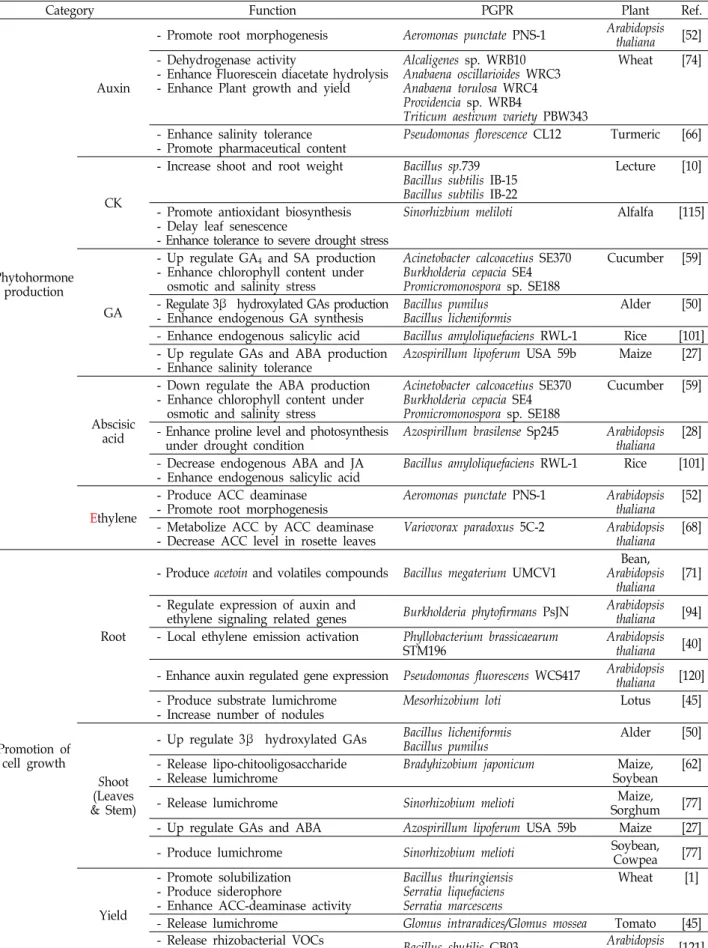
The rhizosphere is the active zone where plant roots communicate with the soil microbiome, each re- sponding to the other’s signals. The soil microbiome within the rhizosphere that is beneficial to plant growth and productivity is known as plant growth-promoting rhizobacteria (PGPR). PGPR take part in many pivotal plant processes, including plant growth, development, immunity, and productivity, by influencing acquisition and utilization of nutrient molecules, regulation of phytohormone biosyn- thesis, signaling, and response, and resistance to biotic- and abiotic-stresses. PGPR also produce secon- dary compounds and volatile organic compounds (VOCs) that elicit plant growth. Moreover, plant roots exude attractants that cause PGPR to aggregate in the rhizosphere zone for colonization, improv- ing soil properties and protecting plants against pathogenic factors. The interactions between PGPR and plant roots in rhizosphere are essential and interdependent. Many studies have reported that PGPR function in multiple ways under the same or diverse conditions, directly and indirectly. This review focuses on the roles and strategies of PGPR in enhancing nutrient acquisition by nutrient fix- ation/solubilization/mineralization, inducing plant growth regulators/phytohormones, and promoting growth and development of root and shoot by affecting cell division, elongation, and differentiation.
We also summarize the current knowledge of the effects of PGPR and the soil microbiota on plants.
Key words : Nutrient, PGPR, phytohormone, plant growth and development, rhizosphere
†