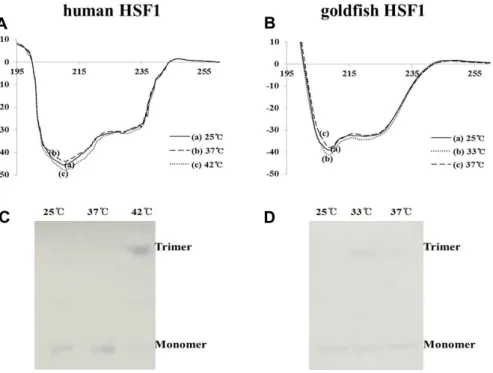
Comparison of Thermal Stress Induced Heat Shock Factor 1 (HSF1) in Goldfish and Mouse Hepatocyte Cultures
So-Sun Kim
1, Jae-Hyeong So
1and Jang-Su Park
1,2*
1
Department of Chemistry, Pusan National University, Busan 609-735, Korea
2
전체 글
1
2
수치

관련 문서
연병장에서의 입소행사가 끝나고 나를 위해 훈련소까지 와 준 친구들과 헤어지는 순간 내 자신이 스스로 군복무를 이행하기 위해 모든 것을 뒤로 하고
Sharing about the 'Hyundai Motor India Foundation ( HMIF)' , he said HMIF was formed in the year 2006 with the purpose of 'giving back' to society and to
제9조(수당의 지급) ① 식약처장은 해외정보리포터의 보고서에 대해 제8조제1항의 평가를 거쳐 각 보고서에 대한 수당을 별표 7의 수당 지급 기준에 따라 분기별로
○ 해외 시장에서의 높은 신뢰도와 인지도를 바탕으로 한국 경제의 성장 동력으로 자리잡아가는 국내 유명 화장품 제조업체를 보호하고 , 국내 뿐만
Recognizing that a S&T Co–operation Fund of 10 million USD, contributed for the period 2011–15, has been utilized to implement joint research.. projects in the
- CERAGEM center owners, staffs and customers from Bihar state participated by donation to support daily necessities & living expenses
Rule 6(1)에 따라 원칙적으로 매입 재료 또는 서비스가 면세 재화의 제조 또는 면세 서비스의 공급에 사용될 경우 Cenvat Credit를 이용할 수 없다. 따라서
In addition, to investigate whether the difference in differentiation patterns between wild-type and Dab1 −/− cells was due to a loss of Reelin- Dab1 signaling, we
