LPS 유도 RAW 264.7 세포와 마우스 모델에서 참치(Katsuwonus pelamis) 유의 항염증 효과
강보경1, 김민지2, 김꽃봉우리2, 안나경1, 최연욱1, 박시우1, 박원민1, 김보람1, 박지혜1, 배난영1, 안동현1*
1부경대학교식품공학과/식품연구소
2부경대학교수산과학연구소
Received: December 2, 2014 / Revised: January 30, 2015 / Accepted: March 5, 2015
서 론
염증은외부감염이나, 생체내의대사산물로인한다양 한자극에대한생체내방어기전으로, 다양한염증매개인 자의활성화에따라홍반, 부종, 열, 통증및기능이상등을 유발하는 반응이다[34]. 특히 대식세포(macrophage)는 lipopolysaccharide (LPS)의자극에의해활성화되어 nuclear factor-kappa B (NF-κB) 의존 경로와 mitogen activated protein kinases (MAPKs) 의존경로의두가지경로를통해
염증반응을유도하게 된다. 그결과 inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2)가 발현되고, nitric oxide (NO) 및 pro-inflammatory cytokine을분비하여
염증반응을유도한다[38]. 이러한염증반응은다양한질병
의생리및병리학적과정에폭넓게관여하는것으로, 특히 만성질환의유병률이증가하고있는현대에서염증반응은 치매, 심혈관질환, 비만, 대사성증후군및비만등의만성질
환의원인으로주목받고있다[10]. 따라서다양한염증반응
매개인자와신호전달경로의단백질발현을조절할수있는 물질은만성질환의예방및억제를위한기능성소재로활
용할수있는가능성이높다고여겨지고있다[9]. 이러한염
증반응의억제를위해다양한스테로이드및비스테로이드 성항염증제가사용되고있으나, 부작용발생우려로인해장 The Anti-inflammatory Effect of Skipjack Tuna (Katsuwonus pelamis) Oil in LPS-induced RAW 264.7 Cells and Mouse Models
Bo-Kyeong Kang1, Min-Ji Kim2, Koth-Bong-Woo-Ri Kim2, Na-Kyung Ahn1, Yeon-Uk Choi1, Si-Woo Bark1, Won-Min Pak1, Bo-Ram Kim1, Ji-Hye Park1, Nan-Young Bae1, and Dong-Hyun Ahn1*
1Department of Food Science & Technology/Institute of Food Science, Pukyong National University, Busan 608-737, Republic of Korea
2Institute of Fisheries Sciences, Pukyong National University, Busan 619-911, Republic of Korea
This study was carried out to demonstrate the anti-inflammatory effect of tuna oil (TO) using LPS-induced inflammation responses and mouse models. First, nitric oxide (NO) and pro-inflammatory cytokines levels were suppressed up to 50% with increasing concentrations of TO without causing any cytotoxicity. Also, the expression of a variety of proteins, such as inducible nitric oxide synthase (iNOS), cyclooxygenase-2 (COX-2) and nuclear factor kappa B (NF-κB), was suppressed in a dose- dependent manner by treatment with TO. Furthermore, TO also inhibited the phosphorylation of mitogen-activated protein kinases (MAPKs), including c-Jun N-terminal kinase (JNK), extracellular signal-regulated kinase (ERK), and p38 protein kinase (p38). Moreover, in in vivo testing the formation of ear edema was reduced at the highest dose tested compared to that in the control, and a reduction of ear thickness and the number of mast cells was observed in histological analysis. In acute toxicity test, no mortalities occurred in mice administrated 5,000 mg/kg body weight of TO over a two-week observation period. Our results suggest that TO has a considerable anti-inflammatory property through the suppression of inflammatory mediator pro- ductions and that it could prove to be useful as a potential anti-inflammatory therapeutic material.
Keywords: Tuna oil, anti-inflammatory effect, ear edema, cytokine
*Corresponding author
Tel: +82-51-629-5831, Fax: +82-51-629-5824 E-mail: [email protected]
© 2015, The Korean Society for Microbiology and Biotechnology
기간사용이어려워, 최근이러한제제를대체할수있는천 연물유래유효성분을얻으려는노력이증가하고있다[28].
한편, 참치(Katsuwonus pelamis)는농어목고등어과의바 닷물고기로, 단백질함량이높아영양학적으로우수하며, 특 히 eicosapentaenoic acid (EPA, 22:5n-3)와 docosahexaenoic acid (DHA, 22:6n-3)와같은다가불포화지방산(polyunsaturated fatty acids, PUFA)를다량함유하는대표적인등푸른생선 이다[18]. 일반적으로정어리, 참치와같은등푸른생선의어 유에는 EPA 및 DHA가다량함유되어있는데[2], 최근다양 한역학조사및인체와동물을대상으로한연구결과이러 한 n-3계고도불포화지방산이체내에서다양한생리활성기 능을가지는것으로보고되었다. 특히혈액내중성지방, 콜 레스테롤수치를감소시키고, 혈소판응집을감소시켜심혈 관계질병을예방하고[33, 44], 뇌, 망막, 신경조직의성장과
발달에직접적인영향을미치는것으로알려져있다[40]. 이
렇듯 EPA 및 DHA의우수한생리활성효과에대해다방면
의연구가국내외에서활발히이루어지고있으며, 특히염증 mediator인프로스타글란딘, 트롬복산, 류코트리엔등의생
합성을억제하고[35], 인체단핵구및마우스의대식세포에
대해 tumor necrosis factor-α (TNF-α) 및 interleukin-1β (IL-1β)와같은 cytokine을감소시키고[5], NF-κB 전사단계 의구성요소에영향을미쳐[45] 항염효과를가지는것으로 보고된바있다.
그러나아직까지참치로부터분리한참치유에대한연구 로는그이용을위한분리정제법[39] 및탈취공정에대한연 구[3]와, 항암효과[20] 및지방대사에미치는영향[22]에대 한연구가대부분이며, 항염증효과및그기전에대한연구 는이루어진바없다.
따라서본연구에서는인체유용물질인 EPA 및 DHA를
다량함유하는참치유의항염증효과및기전을in vitro 및
in vivo type의실험모델을이용하여밝히고자하였으며, 본 연구결과가참치유의다양한식품분야에대한이용및우 수한염증예방및치료제로의소재개발에기여할수있을 것으로사료된다.
재료 및 방법
실험재료
본실험에사용한참치유는 skipjack tuna (Katsuwonus pelamis)의 oil로 (주)동원 F&B에서제공받았으며, 밀봉하여 4oC에서저장하며실험전상온에약 5분간방치후사용하 였다.
실험 동물
생후 8주령의수컷 ICR 마우스를오리엔트바이오(Orient
Co., Seongnam, Korea)로부터구입하여귀부종및조직관 찰실험에사용하였다. 단기독성평가에는생후 10주령의 암컷 Balb/c 마우스를이용하였다. 마우스는온도 20±2oC,
습도 50±10%, 12시간명암주기가유지되는동물사육실에
서 1주일간예비사육한후실험에사용하였으며, 동물실험 은부경대학교동물실험윤리지침을준수하여, 부경대학교 동물실험윤리위원회의승인을받아시행되었다.
세포배양
RAW 264.7 세포는한국세포주은행(KCLB 40071, Seoul,
Korea)에서 분양받아 사용하였으며, DMEM (GIBCO,
Grand Island, NY, USA)에 10% inactivated fetal bovine serum 및 1% penicillin-streptomycin을첨가한배지를배양 액으로 37oC, 5% CO2조건에서배양하였다.
세포 독성 측정
시료의세포독성을 MTT assay를통해 살펴보았다[32].
RAW 264.7 세포를 1 × 106 cells/ml의농도로 96-well plate 에 seeding하고, 20시간동안전배양후 TO를농도별(0.1, 1, 10, 50, 100 μg/ml)로첨가하여 37oC, 5% CO2 incubator (MCO-15AC, Sanyo, Osaka, Japan)에서 24시간동안배양 하였다. 배양후, 5 mg/ml의 MTT 시약(3-(4,5-dimethylthiazol- 2-yl)-2,5-diphenyltetrazolium bromide (MTT))을첨가하여 2시간재배양한후이를 4oC, 2,000 rpm에서 10분간원심 분리(UNION 32R, Hanil Co., Incheon, Korea)하여상층액 을제거하였다. 그후, 각 well에 DMSO를첨가하고생성된 formazan을용해시켰으며, 이를 microplate reader (Model 550, Bio-rad, Richmond, CA, USA)를이용하여 540 nm에 서흡광도를측정하였다. 세포증식능은다음식에의해계산 하였다.
Proliferation Index (%)
= sample 흡광도/control 흡광도 × 100
NO 생성량 측정
Griess 반응[26]을이용하여배양액내에존재하는 nitrite 농도를측정하였다. RAW 264.7 세포를 2.5 × 105 cells/ml의 농도로 24-well plate에 접종하고, 5% CO2 incubator (MCO-15AC, Sanyo, Osaka, Japan)에서 20시간전배양하 였다. 세포에 1 μg/ml의 LPS와 시료를농도별(0.1, 1, 10, 50, 100 μg/ml)로처리하여 24시간재배양하였다. 배양액의 상층액을얻은후, 상층액과 griess 시약(1% sulfanilamide + 0.1% naphthylendiamine dihydrochloride, 1:1)을실온에서 10분간반응시키고, microplate reader를이용하여 540 nm 에서 흡광도를측정하였다. 세포 배양액내 NO의농도는
sodium nitrite (NaNO2)의농도별표준곡선과비교하여산 출하였다.
Pro-inflammatory cytokine 분비량 측정
Pro-inflammatory cytokine 분비 억제 효능을 알아보기 위하여, RAW 264.7 세포를 2.5 × 105 cells/ml로 24-well plate에접종하고 18시간동안전배양하였다. 세포에 1 μg/
ml의 LPS와 TO를처리하여 12시간재배양하였다. 배양액 내의 IL-6, TNF-α 및 IL-1β cytokine의 분비량을 ELISA kit (Mouse ELISA set, BD Bioscience, San Diego, USA) 를이용하여 측정하였다. 이를 위해 ELISA microplate에 capture antibody로 anti-mouse IL-6, TNF-α및 IL-1β를 분주하여 4oC에서하룻밤동안 coating하였으며, 이를 0.05%
tween 20을 포함하는 PBST (Phosphate buffered saline with 0.05% tween 20)로 세척하고 10% FBS 용액으로 blocking 하였다. PBST로세척후, 각배양상층액을분주 하고실온에서 2시간반응시켰다. 다시 PBST로세척한후 biotinylated anti-mouse IL-6, TNF-α detection antibody 와 streptavidin-horseradish peroxidase conjugate를첨가 하여실온에서 1시간반응시켰다. IL-1β의경우, biotinylated anti-mouse IL-1β detection antibody를첨가하고 1시간반 응 후, streptavidin-horseradish peroxidase conjugate를 첨가하여 30분반응시켰다. 그후, 이를다시 PBST로세척
한다음, OPD 용액을첨가하여실온에서 30분암반응시켰
다. 2 N H2SO4로 반응을 종료시킨 후 microplate reader (Model 550, Bio-rad)를이용하여 490 nm에서흡광도를측 정하였다.
iNOS, COX-2 및 NF-κB 발현량 측정
iNOS, COX-2 및 NF-κB 발현에미치는 TO의효과를알
아보기위해 RAW 264.7 세포를 DMEM 배지를이용하여
iNOS, COX-2의경우 1 × 106 cells/ml, NF-κB의경우 4 × 106 cells/ml으로조절하여 20 시간전배양하였다. 세포에 1 μg/ml 의 LPS와 0.1, 1, 10, 50, 100 μg/ml의 TO를처리하여 iNOS, COX-2의경우 18시간, NF-κB의경우 30분재배양하였다. 배양이끝난세포를수집하여 3회 PBS (phosphate buffered saline)로세척한후세포질내의 iNOS 및 COX-2 단백질측 정을 위하여 cytosol extraction buffer (50 mM HEPES (pH 7.4), 150 mM NaCl, 5 mM ethylenediaminetetraacetic acid (EDTA), 1% deoxycholate, 5 mM phenylmethylsulfonyl fluoride (PMSF), 1μg/ml aprotinin, 1% triton X-100, and 0.1% NP-40)를첨가하여 30분간 4oC에서 lysis시켰다. 핵내 의 NF-κB p65 단백질측정을위하여먼저 hypotonic lysis buffer (10 mM HEPES, 10 mM KCl, 1.5 mM MgCl2)를 첨가하여 15분간 4oC에서 lysis시켜 crude cytoplasmic
fraction을 제거한 후, 남은 pellet에 nuclear extraction buffer (10 mM HEPES, 100 mM NaCl, 1.5 mM MgCl, 0.1 mM EDTA, 0.1 mM dithiothreitol (DTT))을첨가하여 lysis시킨후, 12,000 rpm에서 20분간원심분리하여세포막 성분 등을 제거하였다. BCA protein assay kit (Pierce, Rockford, IL, USA)를 사용하여 단백질을 정량하였으며, 30μl의 lysate를 10% SDS-PAGE로분리하였다. 분리된단 백질은 PVDF (polyvinylidene difluoride) membrane (Bio- rad)에 200 mA에서 1시간동안전사시킨후, 5% Skim milk 가포함된 TBS (tris buffered saline; pH7.5) 용액으로상온 에서 2시간 동안 blocking하였다. iNOS, COX-2 및 NF-κB p65의항체로는 anti-mouse iNOS, COX-2 및 NF-κB p65 (Santa Cruz Biotechnology, Inc., Dallas, TX, USA)를 사
용하여 1:500으로 희석하고 상온에서 2시간 반응시키고
TBST로 3회세정하였다. 2차항체로 horseradish peroxidase가 결합된 anti-mouse IgG 및 anti-rabbit IgG를 1:2,000으로 희석하여상온에서 1시간반응시킨후, TBST로 3회세정하 여 ECL 기질과 1-3분간반응후각각의단백질밴드는 Gene tool (GeneGnome5, Syngene, Cambridge, UK)를이용하여 가시화하였다.
MAP kinase (JNK, ERK, p38) 발현량 측정
MAPKs의발현에미치는 TO의억제효과를알아보기위
해 RAW 264.7 세포를 DMEM 배지를 이용하여 1 × 106 cells/ml으로 18시간전배양하고 TO를농도별(0.1, 1, 10, 50, 100 μg/ml)로처리하여 30분동안본배양하였으며, 이 후의실험은 iNOS, COX-2 및 NF-κB와동일한방법으로진 행하였다. 인산화된 c-Jun N-terminal kinase (JNK), extracellular signal-regulated kinase (ERK), p38 protein kinase (p38)의 발현 양을 검토하기 위한 항체로는 anti- mouse JNK, ERK 및 p38 (Cell Signalling Technology, Inc., Denvers, MA, USA)을이용하여 1:5,000으로희석하여 사용하였다.
귀 부종 측정 및 조직 관찰
TO의항염증효과에대한in vivo type 실험의일환으로 귀부종억제율을측정하고조직관찰을진행하였다. ICR 마 우스에 TO를 10, 50 및 250 mg/kg body weight으로 200μl 씩경구투여하고, 1시간후, 오른쪽귀에 2.5% croton oil을 20 μl/ear 농도로도포하였다. Croton oil을처리 5시간후에 귀두께를측정하였으며, 두께의증가를부종의형성으로간
주하였다. 조직 관찰은 ICR 마우스의 오른쪽 귀에 TO를
100 mg/ml 농도로 20 μl씩도포하고 15분뒤, 5% croton oil 을 20 μl씩도포하였다. 6시간뒤, diethylether로마취사시 키고, 귀조직을절제하여 10% formaldehyde에 72시간고
정하였다. 고정후파라핀 block을만들어박편을제조하고 hematoxylin-eosin (H&E) 및 toluidine-blue (TB) 염색을 통해조직을관찰하였다.
단기 독성 평가
TO의안전성을평가하기위해단회투여에의한급성경
구독성평가를실시하였다. 본실험은식품의약품안전처고 시제 2014-136호(2014년 7월 30일개정) 의약품등의독성 시험기준[41]에의거하여진행하였다. Balb/c 마우스를실험 시작 전 4-6시간 정도 절식시킨후에 TO를 300, 2,000 및 5,000 mg/kg body weight 농도로경구투여하였으며, 이후 6시간동안비정상적인행동등의경과를관찰하였고 2주까 지사망여부를지속적으로관찰하였다.
통계처리
모든실험결과에대한유의차검정은 SAS software (SAS Institute, Inc., Cary, NC, USA)에서평균값을분산분석한 후, Duncan's multiple range test법에 따라 p < 0.05 수준 에서검정하였다.
결과 및 고찰
참치유의 세포독성
참치유의 RAW 264.7 cell에대한세포독성을알아보고실 험농도조건의설정을위해 MTT assay를실시하였다. 그 결과(Fig. 1A), TO를 0.1-100 μg/ml으로처리시농도의존 적으로세포증식률이증가하였으며, 특히 TO를 100 μg/ml 의농도로처리시무처리구에비해약 113%의증식능을나
타냈다. 이를통해, TO가 100 μg/ml의농도까지 RAW 264.7 cell에대한세포독성을나타내지않음을확인하였고따라서 본연구에사용된 TO의최고농도는세포에독성을나타내 지않는 100 μg/ml으로설정하였다. 이와동일한결과로 Cha
와 Lim 등[8]도치자추출물의처리농도가증가함에따라
유의적인대식세포의증식능을보였으며, 이에대해세포독 성을나타내지않았다고보고하였다.
NO 억제 효과
NO는체내염증반응시대식세포와같은면역세포에서 iNOS에의해 L-arginin으로부터생성되는 radical이다. 이 러한 NO는체내에서 2차신호전달자로서생체방어기능, 신 호전달기능등의주요한역할을하지만[24], 과도한 NO의 발생은숙주의염증반응을심화시켜숙주세포의파괴, shock 에의한혈관확장, 염증반응에의한세포및조직손상을일 으킬뿐만아니라 septic shock 등의만성질환및자가면역 질환을유발시켜문제가되고있다[15]. 따라서현재 NO 생 성저해효과를가지는물질에대하여염증반응을 비롯한
septic shock, 동맥경화조절제로서의가능성에관한연구가
활발히이루어지고있다[6]. TO의처리에의한 NO 생성정 도를측정하기위하여 RAW 264.7 세포에 LPS와 TO를농 도별(0.1, 1, 10, 50 및 100 μg/ml)로첨가하여생성된 NO를 griess 시약을이용하여측정하였다. 그결과(Fig. 1B), LPS 만을처리한대조구에서생성된 NO의양이 56.67±0.49 μM 이었으며, TO를 100 μg/ml의농도로처리시 30.83±0.42 μM 으로약 45%의 NO 억제효과를보임을확인하였다. 본실 험에사용한 TO와같이, 일반적으로어유에는 EPA 및 DHA 와같은 long-chain의 PUFA가다량함유되어있는것으로
Fig. 1. The in vitro anti-inflammatory effect of tuna oil (TO) in RAW 264.7 cells. (A) cell viability of RAW 264.7 cells, (B) NO levels determined from the culture supernatant of RAW 264.7 cells. Proliferation index = (sample O.D / control O.D) × 100. Means with different letters (a-g) above bars are significantly different (p < 0.05).
보고되었다[11]. 본연구결과를통해 TO가염증반응에서 NO 억제효과를가짐을알수있었으며, 이는 Khair-El-Din 등[16]이 fish oil의주성분인 DHA가염증 반응시핵내의 iNOS의전사단계에서 iNOS mRNA의발현및전사를억제 하며, iNOS promoter를억제하는 3단계의억제기작을통 해궁극적으로 NO 발생을현저히억제하여항염증효과를 가진다는보고와일치하였다.
Pro-inflammatory cytokine 억제 효과
염증반응에서 cytokine의분비에미치는 TO의효과를살 펴보기위해, LPS로염증반응을유도한 RAW 264.7 cell에 TO를농도별(0.1, 1, 10, 50, 100 μg/ml)로처리하여배양상 층액의 IL-6, TNF-α및 IL-1β의양을 ELISA kit를이용하 여정량하였다. 그결과, 모든 cytokine에대하여농도의존 적억제효과를보임을확인하였다. 먼저 IL-6의경우(Fig.
2A), LPS 단독처리시약 271.93±8.33 pg/ml의분비를보 였으나, TO를 50 및 100 μg/ml로처리시각각 35.08±0.93, 18.07±2.78 pg/ml으로 매우 낮은 분비량을보였다. 특히 100 μg/ml로처리시에는 93% 이상의억제효과를보였다. 또한 TNF-α의경우(Fig. 2B)에도마찬가지로우수한농도 의존적억제효과를보였는데, 1 μg/ml의낮은농도로처리 시에도 50% 이상의억제효과를보임을확인하였다. IL-1β 의경우, LPS 단독처리시 24.64±4.13 pg/ml의분비를보 였으나, 100 μg/ml로 TO를처리하였을때 4.20±0.00 pg/
ml의분비를보여 83%의억제효과를보였다. TO의주성분
으로보고된 EPA 및 DHA는다양한연구에서항염증효과
가 보고된 바 있으며[1, 16, 27], 본 연구 결과에서 pro- inflammatory cytokine에대해우수한억제효과를보이는
것은주성분인 EPA 및 DHA의효과에기인한것으로사료
된다. 특히 Lo 등[27]의연구에따르면, LPS로유도한 RAW 264.7 cell의염증반응에서 EPA 처리시 TNF-α를효과적으 로억제한다고보고하였으며, Aldridge [1]의보고에따르면 n-3 지방산에멀젼인 omegaven 처리시우수한 TNF-α분 비억제 효과를가진다고보고하였다. LPS는대식세포의 TLR-4와결합하는 ligand로, 표면수용체복합체의활성화 를통해 MyD88, TIRAP/Mal, TRIF 및 TRAM과같은 TIR 영역을포함하는어댑터단백질과결합하여하위신호망의
활성화를매개한다[21]. 대식세포에서 LPS에의해분비되는
IL-6, TNF-α 및 IL-1β는 pro-inflammatory cytokine으로 세균의침입을효과적으로방어하는역할을한다. 그중 IL- 6는 B cell을 plasma cell으로분화시켜항체생산을촉진하 여급성염증반응을만성단계로전환시키는기능을한다[13].
또한 TNF-α는전신성염증에관여하는 cytokine으로, 대식 세포이외에도 NK cell이나 CD4+ lymphocyte에서도생산 된다. 이러한 TNF-α가과량으로생산되게되면발열, 세포 사멸등의반응이유도되며, IL-1β및 IL-6의발현을촉진시 킴으로써패혈증, 알츠하이머, 암, 염증성장질환등의다양 한질병의요인이된다[12, 13]. 다음으로 IL-1β는활성화된 대식세포에서 caspase-1에의해활성형으로전환되어염증 반응의주요매개체로작용하며, 특히 Th cell을활성화시키 고 T cell로하여금 IL-2와같은 cytokine의분비를증가시켜 면역반응을활성화시킨다[8]. 따라서본연구결과, TO가염 증반응에서활발하게작용하여염증반응을활성화시키는 Fig. 2. The in vitro anti-inflammatory effects of tuna oil (TO)
in RAW 264.7 cells. The levels of IL-6 (A), TNF-α (B), and IL-1β (C) from the culture supernatant of RAW 264.7 cells. Means with different letters (a-g) above bars are significantly different (p < 0.05).
pro-inflammatory cytokine의분비를효과적으로억제하는 것을확인하여우수한항염증효과를가짐을확인하였다.
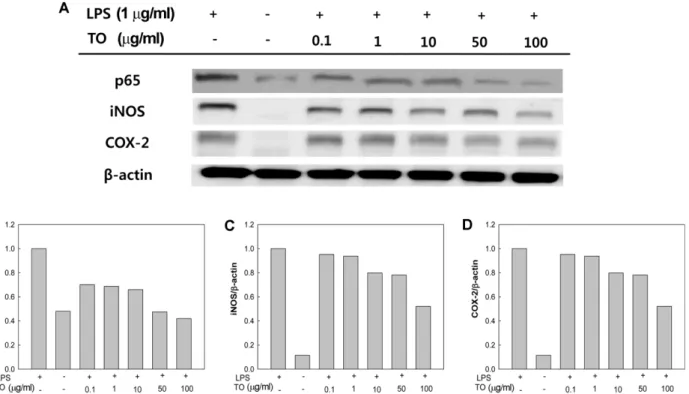
iNOS, COX-2, NF-kB 억제효과
LPS에의해유도되는염증반응은 NF-κB 및 MAPKs의두 가지 pathway에의해일어난다[14]. 특히 NF-κB는전사인 자로염증반응및면역반응의다양한유전자발현에관여 하여, 종양형성, 자가면역질환및염증반응에중요한역할 을 담당한다[23]. 이러한 NF-κB는 세포질 내에 inhibitor kappa B (IκB)와결합되어 불활성화 상태로 존재하는데, LPS와같은자극에의해이 IκB가인산화되고 NF-κB와분 리되면서활성을가지게된다. 이과정에서 inhibitor kappa B kinase (Iκk)가 활성화되어, IκB alpha (IκBα)의 N- terminal 영역의 serine 잔기를 인산화시키고, 인산화된 IκBα는그 lysine 잔기가 multi-ubiquitination되어분해된 다. 이때 IκBα가분해되면, NF-κB/IκBα의복합체상태에 서는불활성화상태였던 NF-κB의핵이동서열이노출되면 서 활성화되어 NF-κB가 핵 내로 이동하여 target gene promoter를일으켜 iNOS, COX-2, 전염증성 cytokine의생 성을유도한다[19]. 이렇게활성화되는인자중 iNOS는 NO 생성효소인 NOS의 한종류로, NOS에는 iNOS 이외에도 neuronal NOS (nNOS), endothelial NOS (eNOS)의동종 효소가알려져있다. 이중 iNOS는대식세포가 interferon-
γ나 LPS 등의다양한염증유도인자에노출된경우발현하
는효소로, 세포내에서 L-arginine을 NO로전환시키는기능 을하여, 과량의 NO를생성시킨다[19, 36]. 이렇게생성된 NO는염증매개물질로작용하여염증성 cytokine의발현을 유도하거나 COX-2의생성을촉진함과동시에활성산소와 결합하여과산화질소등다른독성유기물질을만들어조직 을손상시키게된다[36]. 염증반응에서또하나의주요염 증매개물에는 prostaglandin (PG)가있는데, 이는 COX-2에 의해 arachidonic acid로부터유도되는물질로, 염증반응시 통증의 원인 물질로알려져 있다. 이러한 PG를생성하는 COX-2는체내에 2가지동종효소로존재하는데, 정상조직 에서일정한수준으로상시발현되어위장관보호및신장 혈류조절, 혈소판응집등인체기능 유지의기능을하는 COX-1 외에, 염증반응시에만발생되어다량의 PG를생산 하는 COX-2가있다[36, 37]. 본연구에서는 LPS로염증반 응을 유도한 RAW 264.7 cell에 TO를농도별로 처리하여 NF-κB와, NF-κB의 downstream 단계의 iNOS 및 COX-2 단백질발현의억제효과를관찰하여, 앞선결과의 cytokine 및 NO 분비에대한 TO의효과가어떠한 pathway를억제함 으로써나타난결과인지알아보고자하였다. 먼저핵내부의 NF-κB 단백질발현을 western blot을통해측정한결과(Fig.
3A), TO를 100 μg/ml 농도로처리시 NF-κB의분비를무처 리구와유사한정도로억제하는것을확인하였다. 또한 iNOS
Fig. 3. Effect of tuna oil (TO) on western blot analysis of NF-κB p65, iNOS, and COX-2 protein expression on LPS-induced inflam- matory responses in RAW 264.7 cells, and (B-D) relative unit of densitometries of such an expression.
및 COX-2의단백질발현도 TO를 100 μg/ml로처리시, 두 경우 모두 약 48%의 억제 효과가 있음을 확인하였다. Weldon 등[43]의연구에따르면 LPS로유도된염증반응에 서 DHA는 NF-κB의 subunit의일종인 p65의핵내수준을 상당히감소시키고, 세포질내의 IκB의발현을증가시키는 기작을통해항염효과를가진다고보고하였고, Lee 등[25]
DHA 및 EPA가 NF-κB의활성을억제함으로써 COX-2 발 현억제효과를가짐을보고하였다. 위결과로미루어볼때, TO 의 NF-κB, iNOS 및 COX-2에대한효과는 DHA 및 EPA에 의한효과인것으로사료된다. 본연구결과를통해, LPS 유 도염증반응에서 TO가전사인자인 NF-κB의활성화를억 제함에따라, 그 downstream 단계인 iNOS 및 COX-2를억 제함으로써 NO, cytokine의발현을억제하여항염증효과를 가짐을유추할수있으며, 본연구에서는다뤄지지않았으 나, TO가 COX-2에대해우수한억제효과를가지는것을통 해 PGs의발현억제효과도가질것으로기대할수있다. MAPKs 억제효과
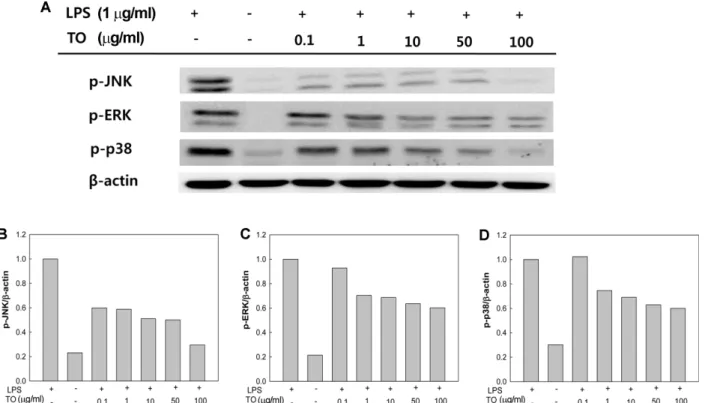
대식세포에서 LPS의자극에의해활성화되는기작에는 TLR4/MyD88/NF-κB 이외에도 MAPKs 의존경로가있다. 이러한 MAPK는포유류에서는 JNK, ERK, p38의 3종이알 려져있으며, 그활성은각효소의 tyrosine과 threonine 잔
기의인산화에의해조절된다[21]. MAPKs의활성형은 down- stream의 activator protein-1 (AP-1), c-AMP responsive element binding protein, activating transcription factor- 2, NF-κB의 serine 및 threonine 잔기를인산화시키는기능 을통해 염증 반응을심화한다[21, 36]. 특히 JNK의경우 Iκk의직접적인인산화를유도하여, NF-κB 신호네트워크 를활성화시키는것으로알려져있다[21]. 또한 ERK 및 p38 은염증의여러경로중에서 iNOS 및 NO 생성에주요한역 할을한다[17, 31]. 따라서본연구에서는 NF-κB 경로억제
효과를가지는 TO가 MAPKs 의존경로에도효과가있는지
를 입증하기 위하여 LPS 유도 염증 반응에서 세포 내
MAPKs의인산화정도를 western blot을이용하여측정하 였다. 그결과(Fig. 4), 세가지의 MAPKs 모두에대해억제 효과가있음을확인하였으며, 특히 p-JNK는 100 μg/ml의농 도로 TO를처리하였을시, LPS 단독처리구에비해약 70%
이상억제효과를보임을확인하였다. 이결과를통해 JNK 의활성이억제됨에따라 Iκk의활성화가일부억제되어 NF- κB에대한억제효과를가지는것을유추할수있다. 이외에 도 p-ERK 및 p-p38 또한 TO를 100 μg/ml으로처리시약 40%의억제효과를보임을확인하였다. Babcock 등[5]의연 구에따르면 omega-3 fatty acid가 AP-1 활성화의바로이전 단계인 JNK의인산화를억제하여 pro-inflammatory cytokine
Fig. 4. Effect of tuna oil (TO) on western blot analysis of p-JNK, p-ERK, and p-p38 protein expression on LPS-induced inflam- matory responses in RAW 264.7 cells, and (B-D) relative unit densitometries of such an expression.