서 론
넙치(Paralichthys olivaceus)는 광염성 어류로서 우리 나라 전 연안, 쿠릴열도, 사할린, 일본 및 중국해 연안 등 에 분포하며, 서식 온도는10~27�C 범위이고 저서에 서
식하는 어류이다(Chyung, 1991). 그리고 이들 넙치는 우 리 나라의 주요 양식 대상종으로 제주도 양식 생산량의 대부분을 차지하고 있으며 2003년의 경우 제주도의 넙 치 양식 생산량은13,000톤에서15,000톤 정도로 추정되 고 있다.
제주도는 지형적인 특성상 내만이 발달되지 못하였기 때문에 넙치를 위주로 한 양식산업은 육상 수조식 양식 에 의존하여 발전하여 왔으며, 제주도의2002년 12월 현
─
─ 100 ──
염분에 따른 넙치(Paralichthys olivaceus)의 성장과 생존율
김 맹 진∙정 상 철1∙송 춘 복*
제주대학교 해양과학부 어류유전육종실험실, 1수산자원학실험실
Effect of Salinity on Growth and Survival of Olive Flounder, Paralichthys olivaceus
Maeng Jin Kim, Sang Chul Chung1and Choon Bok Song*
Laboratory of Fish Genetics and Breeding, 1Laboratory of Fisheries Resources, College of Ocean Science, Cheju National University, Jeju City 690-756, Korea
This experiment was conducted to learn the effects of salinity on growth and survival of juve- nile olive flounder, Paralichthys olivaceus. Fish were reared in completely recirculating fish tanks for 14 weeks. Six tanks (250 L/tank) were established with three different salinities (5, 20 and 35‰), and thus a pair of tanks was assigned to each salinity. Water temperature was maintained at 20 to 22�C throughout this study. The results showed that growth of the fish was best at 35‰, less at 20‰, and least at 5‰. However, growth differences in total length and body weight of fish in 5, 20 and 35‰ were not statistically significant until the 12th week (P¤
0.05). Growth at 5‰ became significantly slower than at 20 or 35‰ on the 14th week (P⁄0.05).
Although specific growth rate at 20‰ was slightly greater than at 35‰, the difference was not obvious (P¤0.05). However, significant differences in specific growth rate existed between 5 and 20‰, or 5 and 35‰. When considering three food-related factors, such as condition factor (CF), feed intake, and food conversion rate (FCR), there were no meaningful differences among all salinities (P¤0.05). At the end of this experiment, survival rate at 35‰ was 97.5%, and those at 20‰ and 5‰ were 88.8% and 86.2%, respectively. These results indicate that low salinity underground seawater is worth to be utilized for flounder farming industry in Jeju Island.
Key words : Salinity, growth, survival, olive flounder
*Corresponding author: [email protected]
재 육상양식장 어업허가 현황에 따르면 넙치 양식장이 247개소에 이른다.
제주도 연안 수온은 지역에 따라서 겨울철에14�C 이 하의 저수온을 나타내거나 여름철에 25�C 이상의 고수 온의 영향으로 때때로 어류의 대량 폐사를 초래하는데 이러한 문제를 극복하기 위해 제주도의 많은 양식장에서 연중 16~18�C의 수온을 유지하는 지하해수를 사용한다 (Chun and Rho, 1991). 그러나 지하해수는 지역적인 특 성에 따라서 때때로20~30‰ 범위의 저염분을 나타내고 있다. 또한 여름철 집중 강우 및 중국 연안수의 유입으로 인해 황해, 남해안 및 제주도 연안에 저염분이 나타나고 있으며, 특히 8월경에 제주도 연안은 27.7‰이라는 낮은 염분을 나타내기도 하며(Kim et al., 1999) 이로 인하여 수산생물에 많은 피해를 주기도 한다.
저염분은 해산어류의 삼투압 조절에 영향을 미침으로 써 이온과 수분 평형에 혼란을 일으켜 어체의 생리조건 악화 및 성장지연을 초래한다(Singley and Chavin, 1971). 또한, 광염성 어류를 담수에 이동시키면 염류세포 가 증가하고(Hwang, 1987; Cioni et al., 1991) 생리적 변 화로 인한 스트레스를 받거나 심할 경우 사망하는 것으 로 알려져 있다(Hur and Chang, 1999; Chang et al., 2002). 따라서 어류는 이러한 염분 변화에 대처하기 위해 삼투압 조절을 함으로써 체내의 항상성을 유지한다. 어류 의 삼투압 조절과 관련하여 틸라피아, Oreochromis niloticus (Chang and Hur., 1999), 연어과 어류(Toften et al., 2003), 숭어, Mugil cephalus (Chang and Hur, 1999;
Chang et al., 1997), 감성돔, Acanthopagrus schlegeli (Chang et al., 2002) 등에서 염분변화에 따른 생리적 변 동이나 아가미 염류세포의 구조적 변화와 같은 연구들이 발표된 바 있다.
대부분 어류의 성장은 외적인자인 온도, 용존산소, 염분 등에 의해서 크게 영향을 받는 것으로 알려져 있다. 그러 나 연어과 어류나 숭어 등과 같은 어종들은 발생이나 성 장에 있어서 염분변화에 의한 영향은 거의 받지 않는 것 으로 알려져 있다. 염분에 관련된 성장률 및 생존율 실험 은 어린 농어, Lateolabrax japonicus (Han et al., 2001), Acanthopagrus butcheri (Partridge and Jeckins, 2002), turbot, Scophthalmus maximus (Gaumet et al., 1995), spotted wolffish, Anarhichas minor (Foss et al., 2001) 및 숭어(Lee et al., 1996)를 대상으로 연구가 수행되어 왔다. 한편, 넙치를 대상으로 한 관련 연구로서는 알발생과 자치어의 염분 내성(Chun and Rho, 1991), 그리고 저염 분에 의한 생리적 변화에 관한 연구(Hur et al., 2002)가 있으며, Hur et al. (2003)은 30일 동안15‰과 30‰의 염 분에서 넙치를 사육하면서 이들의 생리적 변화를 위주로
성장과 생존율에 관한 결과를 보고하면서 보다 장기적인 사육을 통한 연구의 필요성을 피력한 바 있다. 따라서 이 연구는 보다 장기적이고 보다 저염분에서의 넙치 사육시 이들의 성장률과 생존율에 미치는 영향을 조사함으로써 넙치의 적정 사육 염분 및 사육가능 최저염분을 구명하 여 넙치 양식장에서 다양한 용도로의 저염분 지하해수의 사용 가능성을 제시하고자 수행하였다.
재료 및 방법
1. 실험어, 사육 조건 및 실험설계
실험에 사용된 넙치 치어는 제주도 표선면에 있는 넙 치양식장에서 종묘생산한 치어로 제주대학교 해양과학대 학 어류유전육종 실험실로 운반하여 어병 예방을 위한 약욕을 실시한 후 사육실에 있는 1 ton 순환여과수조에 15일간 수용하였으며, 실험조건에 미리 적응시키기 위해 서 수온은 21±1�C를 유지하였으며 실험 전 1일 동안 절식시켰다.
실험에 사용된 넙치는 총 240마리로써 평균 체중은 13.9±0.2 g, 평균 전장은 11.5±0.6 cm였다. 순환여과수 조 시스템은 여과효율을 고려하고 섭취하지 못한 사료를 쉽게 수거할 수 있도록 직접 설계하고 의뢰하여 제작한 수용적250 L인 6개의 유리수조를 사용하였다. 사육수는 5‰, 20‰ 그리고35‰의3가지 염분조건을 주었으며 동 일 염분조건에 2개의 수조를 사용하여 실험을 수행하였 다. 실험 시작시 장기간의 성장을 고려하여 수조 당40마 리씩 수용하였으며 어체 크기의 증가로 인해5주차에 무 작위 선별한 후 실험 종결 시까지 수조 당 25마리를 수 용하였다. 그런데, 장기간 사육을 통한 성장 실험에서 때 때로 문제가 되는 것은 염분이 성장에 미치는 영향보다 동일 염분조건에서의 두 개 수조간에 일어나는 사망 개 체수의 차이(사육밀도의 차이)나 기타 미소한 사육환경 의 차이로 인한 실험 오차가 클 경우, 염분간 성장차이의 통계학적인 유무에 상당한 영향을 주게되어 실험의 목적 을 달성할 수 없게 된다. 따라서 이러한 문제를 최소화하 기 위하여 어체를 측정할 때마다 각 염분내의 수조간 사 육 중량의 차이가 최소화되도록 인위적으로 조정하면서 사육하였다.
먹이는 시판되고 있는 넙치용 건조침강사료(일본산)를 아침과 저녁으로 나누어서 먹이 유실을 최소화 할 수 있 도록 충분한 시간적 여유를 가지고 각 수조의 전체 사육 어 체중의 3% 정도를 공급하였으며 일주일마다 하루는 절식시켰다. 그리고 최적 사육환경 조성을 위하여 일주일 마다 각 수조 전체 수용적의 50%를 지하해수를 사용하
여 환수하였으며, 사육수온이 성장에 미치는 영향을 최소 화하기 위해서 에어컨과 자동 온도 조절 장치를 이용하 여 사육 수온을 20~22�C로 유지하였고, 사육기간 중 용 존산소량은5.5~6.1 ppm을 나타내었다.
2. 어체 측정과 성장 및 생존율 계산
실험은14주 동안 실시하였으며, 실험 시작 전 전장과 체중을 측정하였고 실험기간 중에는 2주 간격으로 전장 과 체중 측정을 하였다. 측정시의 스트레스를 최소화하기 위하여 어류의 공기 중 노출 시간을 최소화하였다. 체중 은 전자저울을 이용하여0.1 g 단위까지 측정하였으며 전 장은 버어니어 캘로퍼를 이용하여 1 mm 단위까지 측정 하였다. 생존율을 구하기 위해서 매일 사망 개체수를 파 악하여 사망률을 계산한 후 생존율을 추정하였다. 증가된 체중과 전장을 이용하여 일간성장률, 비만도, 그리고 사 료 공급량에 따른 사료 섭식량, 사료전환효율은 다음 식 에 의해 계산하였다(Foss et al., 2001): 일간성장률(speci- fic growth rate) = {(In 실험종료시 평균체중)-(In 실험개 시시 평균체중)}/(사육기간)×100; 비만도(condition fac- tor) = (체중/전장3)×100; 사료섭식량(food intake) = (건조 된 공급한 먹이량)-(사료 부산물); 사료전환효율(food conversion rate) = {(실험종료시 평균체중)-(실험개시시 평균체중)}/(사료섭식량)×100.
3. 통계처리
조사기간 중 각 성장 시기별 체중과 전장의 염분별 차 이와 사료와 관련된 요인들(일간성장률, 비만도, 사료섭 취량, 사료전환효율)의 염분별 차이는 분산분석(analysis of variance, ANOVA)을 이용하여 95% 신뢰한계에서 유 의차를 검정하였다. 분산분석 결과 유의한 차이를 나타내 는 경우, 다중검정(multiple comparison test)을 실시하여 각 요인내 두가지 염분간의 평균값 사이에 유의한 차이 여부를 조사하였으며 검정 방법은 Tukey HSD test (see Zar, 1984)를 사용하였다. 체중과 전장의 상대성장식을 구하기 위하여 비선형분석(nonlinear regression analy- sis)을 하였으며 분석시Gauss-Newton 방법을 이용하였 다. 모든 통계분석은 ‘SYSTAT 10’ (SPSS Inc.)을 사용하 여 수행하였다.
결 과
1. 성장
실험기간 동안 넙치(Paralichthys olivaceus) 치어의 염
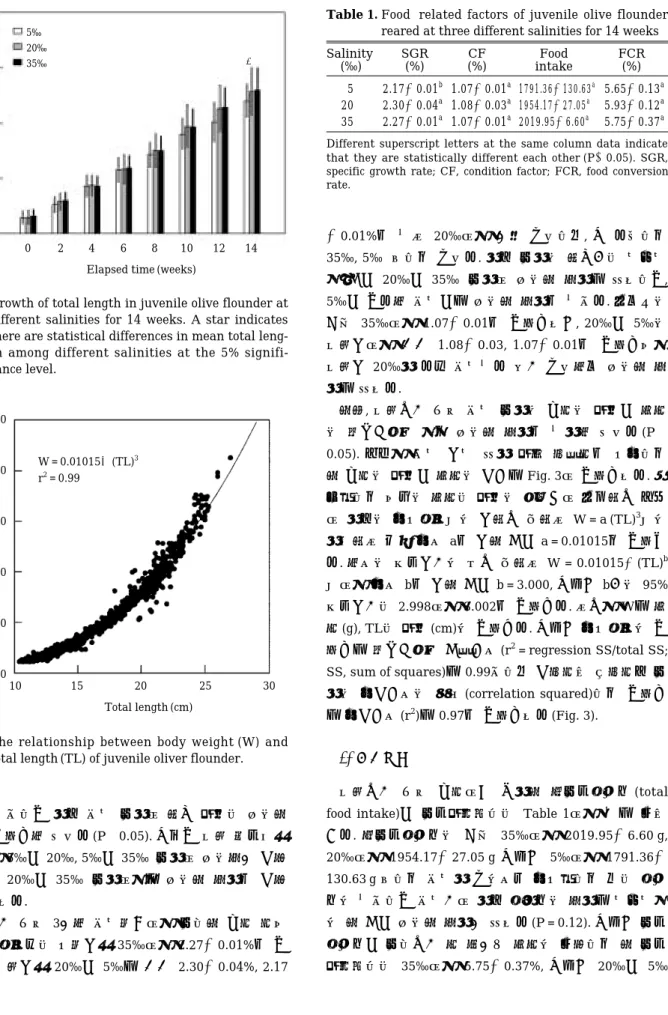
분 농도에 따른 시기별 체중 성장은 지수곡선 양상를 보 였다(Fig. 1). 실험개시시 35‰, 20‰, 5‰ 염분에 투입한 넙치의 평균 체중은 각각 14.37±2.02 g, 13.70±1.94 g, 13.74±1.87 g으로 염분별 평균 체중 사이에 유의한 차 이는 없었으나(P¤0.05), 실험 종결시에는 각각130.62± 31.49 g, 129.60±35.37 g, 114.86±26.13 g을 나타내어 평 균 체중 사이에 유의한 차이를 보였다(P⁄0.05). 실험 시 기별 염분별 평균 체중은 실험 후 6주간은 별로 차이를 보이지 않다가 실험 후8주부터는 실험 후 12주까지 다 소 차이를 보이며20‰에서 가장 빠른 성장을 보였으나 이들 염분 사이에 평균 체중의 유의한 차이는 나타내지 않았다(P¤0.05). 그러나 실험 종료시인 실험 후 14주에 서 평균 체중은35‰, 20‰ 그리고 5‰ 순으로 높게 나 타나서 순위에서 20‰과 35‰이 바뀌었으며 분산분석 결과 유의한 차이를 보였다. 따라서 다중검정을 실시한 결과5‰과20‰, 5‰과 35‰ 사이에 유의차가 관찰되었 으나 20‰과 35‰ 사이에서는 유의한 차이를 관찰할 수 없었다.
염분 농도에 따른 시기별 실험어의 전장 성장은Fig. 2 에 나타내었다. 실험 개시시 35‰, 20‰, 5‰ 염분에서의 넙치의 평균 전장은 각각 11.58±0.60 cm, 11.49±0.62 cm, 11.47±0.59 cm로써 염분별 평균 전장 사이에 유의 한 차이는 없었으나(P¤0.05), 실험 종결시에는 23.02± 1.97 cm, 22.87±2.10 cm, 22.02±1.47 cm를 보여 평균 전장 사이에 유의한 차이를 보였다(P⁄0.05). 실험 시기 별 염분별 평균 전장은 평균 체중의 경우와 마찬가지로 실험 후6주간은 별로 차이를 보이지 않다가 실험 후8 주부터는 실험 후 12주까지 5‰과 다른 염분에서 다소
Fig. 1. Growth of body weight in juvenile olive flounder at different salinities for 14 weeks. A star indicates there are statistical differences in mean body wei- ght among different salinities at the 5% signifi- cance level.
5‰
20‰
35‰
150
100
50
0
Body weight(g)
0 2 4 6 8 10 12 14
Elapsed time (weeks)
★
차이를 보였으나 이들 염분 사이에 평균 전장은 유의한 차이를 나타내지 않았다(P¤0.05). 그러나 실험 종료시인 14주에서5‰과20‰, 5‰과35‰ 사이에 유의차가 관찰 되었으며 20‰과 35‰ 사이에서는 유의한 차이를 관찰 할 수 없었다.
실험기간 동안 3가지 염분조건에서 사육한 넙치 치어 의 일간 성장률은 대조구인35‰에서 2.27±0.01%를 나 타냈고, 실험구인 20‰과 5‰는 각각 2.30±0.04%, 2.17
±0.01%를 보여 20‰에서 가장 높았으며, 그 다음으로 35‰, 5‰ 순으로 높았다. 이들 사이의 평균값은 분산분 석 결과 20‰과 35‰ 사이에 유의한 차이는 없었으나, 5‰과 나머지 염분과는 유의한 차이를 보였다. 비만도의 경우 35‰에서 1.07±0.01를 나타내었고, 20‰과 5‰의 실험구에서 각각 1.08±0.03, 1.07±0.01를 나타내어서 실험구20‰이 다른 염분보다 약간 높았지만 유의한 차 이는 없었다.
한편, 실험기간 동안 염분 사이의 넙치의 전장과 체중 의 회귀곡선은 서로 유의한 차이를 보이지 않았다(P¤
0.05). 따라서 염분 구분 없이 전체 측정치를 대상으로
한 넙치의 전장과 체중의 관계는Fig. 3에 나타내었다. 일 반적으로 어류의 체중은 전장의 세제곱에 비례하기 때문 에 이들의 상대성장식을 구하기 위하여 W = a (TL)3식을 이용하여 먼저 상수a를 구한 결과 a = 0.01015로 나타났 다. 지수의 신뢰구간을 알기 위하여 W = 0.01015×(TL)b 식에서 상수b를 구한 결과b = 3.000, 그리고b값의95%
신뢰구간은2.998에서3.002를 나타냈다. 여기서W는 체 중(g), TL은 전장(cm)을 나타낸다. 그리고 상대성장을 나 타내는 회귀곡선의 결정계수(r2 = regression SS/total SS;
SS, sum of squares)는 0.99였으며 관측치와 예측치들 사 이의 상관계수의 자승(correlation squared)으로 나타내 는 상관계수(r2)는0.97를 나타내었다(Fig. 3).
2. 사료효율
실험기간 동안 넙치에게 급이한 총사료섭식량(total food intake)과 사료전환효율은 Table 1에서 보는 바와 같다. 총사료섭식량의 경우 35‰에서 2019.95±6.60 g, 20‰에서 1954.17±27.05 g 그리고 5‰에서 1791.36± 130.63 g 순으로 염분이 높을수록 상대적으로 많은 섭식 량을 보였으나 염분간에 이들 섭이량의 차이는 분산분석 을 한 결과 유의한 차이가 없었다(P = 0.12). 그리고 사료 섭식량과 사육기간 중 증가된 체중을 바탕으로 한 사료 전환효율은 35‰에서 5.75±0.37%, 그리고 20‰과 5‰
Fig. 2. Growth of total length in juvenile olive flounder at different salinities for 14 weeks. A star indicates there are statistical differences in mean total leng- th among different salinities at the 5% signifi- cance level.
Fig. 3. The relationship between body weight (W) and total length (TL) of juvenile oliver flounder.
Table 1. Food-related factors of juvenile olive flounder reared at three different salinities for 14 weeks
Salinity SGR CF Food FCR
(‰) (%) (%) intake (%)
5 2.17±0.01b 1.07±0.01a 1791.36±130.63a 5.65±0.13a 20 2.30±0.04a 1.08±0.03a 1954.17±27.05a 5.93±0.12a 35 2.27±0.01a 1.07±0.01a 2019.95±6.60a 5.75±0.37a Different superscript letters at the same column data indicate that they are statistically different each other (P⁄0.05). SGR, specific growth rate; CF, condition factor; FCR, food conversion rate.
0 2 4 6 8 10 12 14
Elapsed time (weeks)
10 15 20 25 30
Total length (cm) 5‰
20‰
35‰
30
25
20
15
10
★
Total length(cm)
250
200
150
100
50
0
Body weight(g)
W = 0.01015×(TL)3 r2= 0.99
에서 각각 5.93±0.12%, 5.65±0.13%를 나타내어 20‰
에서 가장 높은 것으로 나타났으나 이들 값 사이에 유의 한 차이는 보이지 않았다(P = 0.55).
3. 생존율
사육 시작7일에서35일 사이에 사망 개체가 출현하였 으며, 그 이후는 사망 개체가 없었다(Fig. 4). 각 염분별 사망시기와 사망개체수를 보면 5‰에서 실험 시작 후 7 일에서 23일째 까지 총 80마리 중 11마리가 폐사하여 생존율은86.2%를 보였고, 20‰에서는 실험 후13일째부 터 실험 후35일째 사이에 총 80마리 중9마리가 사망하 여 생존율은88.8%를 나타내었다. 그리고 대조구인 35‰
의 경우 실험 후13일째부터 실험 후21일 사이에 총 80 마리 중 2마리가 사망하여 생존율은 97.5%를 보였다. 그 러나 실험시작 후36일째부터는 더 이상 사망 개체가 관 찰되지 않아서 생존율의 변화는 없었다. 따라서, 전체적 으로 볼 때 넙치 치어들의 생존율은 35‰, 20‰ 그리고 5‰ 순으로 염분이 높을수록 높았으나 5‰과20‰ 사이 에 큰 차이는 보이지 않았다.
고 찰
어류는 어종에 따라서 다양한 염분 저항성을 갖는다. 즉 회유성 어류인 연어과 어류나 기수 지역에 주로 서식 하는 숭어나 망둥어과 어류, 그리고 해수에서 주로 서식 하는 감성돔이나 농어 등은 담수에서도 서식이 가능하 며, 해수지역에서만 서식하는 어류의 경우에도 넓은 염분 범위에서 서식할 수 있는 염분 저항성을 갖고 있다(Boeuf and Payan, 2001). 이 연구에서 넙치(Paralichthys oli- vaceus)를 대상으로 한14주간의 실험 결과35‰에서 생 존율을 97.5%를 보였고, 염분 20‰와 5‰에서의 생존율
은 각각 88.8%와 86.2%로 35‰에 비해서 다소 낮게 나 타났다. 이러한 결과는 Hur et al. (2003)이 30‰과 15‰
의 염분조건에서 1개월 동안 사육한 후 보고한 넙치의 생존율인96.7±0.0%과 88.3±2.4%와 유사하였다. 또한, Kang et al. (2000)은 염분이 낮은 기수지역에 서식하는 날개망둑(Favonigobius gymnauchen)을40일 동안3.4‰, 6.7‰ 염분에서 사육한 결과 생존율이 각각 70%와 80%
이었다고 보고하였다.
따라서, 5‰에서의 넙치의 생존율(86.2%)이 비슷한 염 분에서의 날개망둑 생존율보다 오히려 높게 나타나서 넙 치의 저염분에 대한 강한 저항성을 알 수 있었다. 그럼에 도 불구하고 실험개시 후10일부터23일 사이에 저염분 인20‰과5‰에서 일어난 생존율의 급격한 저하는 염분 변화에 의한 과도한 스트레스에 기인하는 것으로 생각된 다. 이와 관련하여 Woo and Fung (1981)은 사육수내 과 도한 염분의 변화는 어체 내 생리적 변화를 일으키게 되 며, 그 결과로 삼투조절의 실패를 가져와 사망한다고 보 고한 바 있다.
어류에 있어 염분은 대사활동, 삼투조절 및 생체리듬 등에 영향을 주기 때문에 연어와 같은 회유성 어류나 망 둑어와 같은 기수지역에 서식하는 광염성 어류의 생활사 에 있어서 대단히 중요한 요인으로 작용할 뿐만 아니라 (Kang et al., 2000) 경골어류의 성장과 밀접한 관계가 있 는 것으로 보고되었다. 즉, 어체는 염분변화에 직면하면 체내 항상성 유지를 위해 삼투 조절을 하게 되고, 이를 위해 생체 에너지를 사용하게 된다(Lasserre et al., 1978).
그 결과, 에너지 소비로 인해 성장이 현저히 둔화된다고 알려져 있다(Singly and Chavin, 1971).
이 조사 결과 넙치의 일간성장률은 5‰, 20‰, 35‰ 중 20‰의 염분 조건에서 가장 높게 나타나서 지금까지 밝혀 진 광염성 어류의 최적 성장 염분과 유사하였다(Chang et al., 1996; Kang et al., 2000; Patridge and Jenkins, 2002).
이러한 결과의 원인으로 체액과 등장의 매질에서 체내의 항상성 유지를 위한 삼투조절에 에너지가 가장 적게 이 용되며, 절약된 에너지가 성장에 이용될 수 있기 때문으 로 밝히고 있다(Morgan and Iwama, 1991). 즉, 삼투조절 에 소비되는 에너지가 가장 적게 소모되는 등장액과 비 슷한 수준의 염분범위가 성장에 최적 조건이라는 의미이 다.
또한, 여러 연구자들은 해수보다 낮은 염분에서 해수어 류의 순치 반응과 연관하여 성장에 관한 연구를 수행하 였다(Table 2). 그 결과, 해수보다 낮은 염분에 순응하는 일부 해수어류에서 성장증진 효과가 나타나는 것으로 알 려져 있다(Gaumet et al., 1995; Woo and Kelly, 1995, Partridge and Jenkins, 2002).
Fig. 4. Survival rates of juvenile olive flounder at differ- ent salinities of 35‰ (A), 20‰ (B) and 5‰ (C) for 14 weeks.
0 10 20 30 40 50 60 70 80 90 100
Elapsed time (days)
A
B
C 100
95 90 85 80 75
Survival rate(%)
Arnesen et al. (1993)은 어류가 급격한 염분의 변화에 노출될 경우 삼투적 스트레스로 인해 식욕감퇴를 가져와 서 낮은 사료섭식량이나 사료전환효율을 보이게 된다고 지적한 바 있다. 이 연구에서 14주간의 실험결과 총사료
섭식량은 35‰¤20‰¤5‰을 보였으며, 사료전환효율은
20‰¤35‰¤5‰을 나타내었으나 이들 값들의 염분간
유의한 차이는 보이지 않았다. 그러나, 5‰에서의 일간성 장률의 유의한 차이, 가장 낮은 사료섭식량과 사료전환효 율, 그리고14주째 나타난 체중과 전장 성장의 유의한 차 이는 저염분으로 인한 스트레스의 영향으로 판단된다. 20‰의 경우 일간성장률, 비만도, 사료전환효율이 다른 염분에 비해 가장 양호하였으며 체중과 전장 성장에서도 35‰과 유의한 차이를 보이지 않았다. 따라서, 20‰ 염분 은 넙치 치어의 성장에 해로운 영향을 주지 않는 것으로 밝혀졌다. 5‰에서는 약간의 성장 저하가 관찰되었으나 86.2%의 생존율을 보여 20‰에서의 생존율인 88.8%와 큰 차이를 보이지 않았기 때문에, 넙치 치어는 5‰의 낮 은 염분에서도 삼투 조절이 가능하여 지하해수나 담수가 유입된 해수에서도 안전한 사육이 가능할 것이라고 판단 된다. 따라서, 차후 넙치양식에 있어서 저염분 지하해수 의 이용이 보다 적극적으로 검토되어야 하겠다.
적 요
지하 해수나 저염분의 해수가 넙치(Paralichthys oliv-
aceus)성장에 어떠한 영향을 주는지 알아보기 위해서 14
주 동안 염분에 따른 넙치의 성장과 생존율을 조사하였 다. 실험 조건은5‰, 20‰, 35‰이며 실험어는 평균 체중 13.9±0.2 g, 평균 전장 11.5±0.6 cm인 치어 240마리를 사용하였다. 실험 종결시, 전장과 체중의 성장은 35‰¤
20‰¤5‰ 순으로 나타났으나 35‰과 20‰사이에서는
성장의 유의한 차이가 없었고(P¤0.05), 이들 염분과 5‰
사이에서는 유의한 차이가 관찰되었다(P⁄0.05). 그러나, 12주까지는 이들 염분조건 사이에 성장의 유의한 차이는 관찰되지 않았다(P¤0.05). 사료공급에 따른 사료섭식량 은 35‰에서 가장 많았지만 염분간에 유의한 차이는 없 었다(P¤0.05). 사료전환효율은 20‰에서 가장 높았지만 염분간 유의차는 없었다(P¤0.05). 생존율은 실험 후 14 주까지 35‰에서는 97.5%, 20‰에서는 88.8%, 그리고 5‰에서는86.2%를 나타내어서 5‰과 20‰ 염분에서 큰 차이를 보이지 않았다. 이러한 결과를 종합해보면, 20‰
의 염분에서 사육된 넙치의 성장은 35‰의 일반해수의 결과와 유사하였으며, 일간성장률이나 먹이전환효율은 오히려 20‰의 염분 조건에서 높게 나타났다. 그리고, 5‰에서는 실험 종결시인 14주째에 약간의 성장 저하가 관찰되었으나(P⁄0.05), 생존율은 20‰과 유사하게 나타 났다. 따라서 제주도 넙치 양식 산업에서 다양한 목적으 로 저염분 지하해수를 사용할 수 있을 것으로 생각된다.
사 사
이 연구에 사용한 넙치 치어를 제공해 주신 동남수산 이준민 사장님께 감사의 뜻을 표합니다. 그리고 이 논문 은 2003년도 두뇌한국21사업에 의한 지원과 수산특정과 제 연구비의 지원을 받아 수행되었습니다.
인 용 문 헌
Arnesen, A.M., E.H. Jorgensen and J. Malcolm. 1993. Feed intake, growth and osmoregulation in arctic charr, Salvelinus alpinus (L), following abrupt transfer from freshwater to more saline water. Aquaculture, 114 : 327~338.
Boeuf, G. and P. Payan. 2001. How should salinity influ- ence fish growth? Comp. Biochem. Physiol., 130C : 411
~423.
Chang, Y.J., B.H. Min, H.J. Chang and J.W. Hur. 2002.
Comparison of blood physiology in juvenile black sea- bream (Acanthopagrus schlegeli) reared in converted freshwater from seawater and seawater from fresh- water. J. Korean Fish. Soc., 35(6) : 595~600 (in Korean).
Chang, Y.J. and J.W. Hur. 1999. Physiological responses of grey mullet (Mugil cephalus) and nile tilapia (Oreoch- romis niloticus) by rapid changes in salinity of rearing water. J. Korean Fish. Soc., 32 : 310~316 (in Korean).
Chang, Y.J., Y.C. Lee and B.K. Lee. 1996. Comparison of growth and survival rates of juvenile grey mullets Table 2. Salinity ranges (‰) for tolerance and best grow-
th of different fish species
Species Tolerance growthBest Reference Acanthopagrus
butcheri 0~60 24 Partridge and
(Black bream) Jenkins (2002)
Sparus sarba
Woo and Kelly (1995)
(Sea bream) 0~35 15
Scophthalmus
maximus 0~35 10~19 Gaumet et al. (1995) (Turbot)
Gadus morhus(Atlantic cod) 7~35 7~14 Lambert et al. (1994)
(Mugil cephalus) in different salinities. J. Aquaculture, 9 (4) : 311~320 (in Korean).
Chun, J.H. and S. Rho. 1991. Salinity tolerance of eggs and juveniles of flounder, Paralichthys olivaceus (Temminck et schlegel). J. Aquaculture, 4(2) : 73~84 (in Korean).
Chyung, M.K. 1991. The fish of Korea. Iljisa Pub. Co., Seoul. 727 pp. (in Korean).
Cioni, C., D. De Merich, E. Cataldi and S. Cataudella.
1991. Fine structure of chloride cells in freshwater- and seawater-adapted Oreochromis niloticus (Linna- eus) and Oreochromis mossambicus (Peters). J. Fish Biol. 39 : 197-209.
Foss, A., T.H. Evensen, I.A.K. Imsland and V. Oiestand.
2001. Effects of reduced salinities on growth, food con- version efficiency and osmoregulatory status in the spotted wolffish. J. Fish Biol. 56 : 416~426.
Gaumet, F., G. Boeuf, A. Severe, A. Le Roux and N. Mayer -Gostan. 1995. Effects of salinity on the ionic balance and growth of turbot. J. Fish Biol. 47 : 865~876.
Han, H.K., D.Y. Kang, S.B. Hur and S.W. Kim. 2001.
Effects of temperature and salinity on early develop- ment, survival and growth rate in seabass, Lateolabrax japonicus. J. Aquaculture, 14(1) : 17~27 (in Korean).
Hur, J.W., Y.J. Chang, B.K. Lee and J.Y. Lee. 2003. Effects of hypo-salinity on physiological response, survival and growth of cultured olive flounder (Paralichthys oli- vaceus). Korean J. Ichthyol., 15(2) : 77~86 (in Korean).
Hur, J.W., B.K. Lee, Y.J. Chang, J.K. Lee, Y.S. Lim., J.H.
Lee, C.H. Park and B.K. Kim. 2002. Stress responses of olive flounder, Paralichthys olivaceus to hyposalinity.
J. Aquaculture, 15(1) : 69~75 (in Korean).
Hur, J.W. and Y.J. Chang. 1999. Physiological responses of grey mullet (Mugil cephalus) and nile tilapia (Oreo- chromis niloticus) by gradual change in salinity of rear- ing water. J. Aquaculture, 12(4) : 283~292 (in Korean).
Hwang, P.P. 1987. Tolerance and ultrastructural response of branchial chloride cells on salinity change in euryha- line teleost, Oreochromis mossambicus. Mar. Biol., 94 : 643~649.
Kang, J.C., P. Chin, J.S. Lee, Y.K. Shin and K.S. Cho.
2000. Effects of salinity on survival, growth and oxygen consumption rates of the juvenile gobiid, Favonigobius gymnauchen. J. Korean Fish. Soc., 33(5) : 408~412 (in Korean).
Kim, J.T., D.G. Jeong and H.K. Rho. 1999. Environmental character and catch flutuation of set net ground in the coastal water of Hanlim in Cheju island, II. Fluctuation of temperature, salinity and current. J. Korean Fish.
Soc., 32 (1) : 98~104 (in Korean).
Lambert, Y., J.D. Dutil and J. Munro. 1994. Effects of intermediate and low salinity conditions on growth rate and food conversion of Atlantic cod (Gadus morhua).
Can. J. Fish. Aquat. Sci., 51 : 1569~1576.
Lasserre, P., G. Boeuf and Y. Harache. 1978. Osmotic ada- ptation of Oncorhynchus kisutch Walbaum. 1. Seasonal variations of gill Na±, K±-ATPase activity in coho salmon, 0±-age and yearling, reared in freshwater.
Aquaculture, 14 : 365~382.
Lee, Y.C., Y.J. Chang and B.K. Lee. 1997. Osmoregulation capability of juvenile grey mullets (Mugil cephalus) with the different salinities. J. Korean Fish. Soc., 30 (2) : 216~224 (in Korean).
Morgan, J.D. and G.K. Iwama. 1991. Effects of salinity on growth, metabolism, and ion regulation in juvenile rainbow trout and steelhead trout (Oncorhychus my- kiss) and fall chinook salmon (Oncorhynchus tshawy- tscha). Can. J. Fish. Aquat. Sci., 48 : 2083~2094.
Partridge, G.J. and G.I. Jenkins. 2002. The effect of salinity on growth and survival of juvenile black bream (Acan- thopagrus butcheri). Aquaculture, 210 : 219~230.
Singley, J.A. and W. Chavin. 1971. Cortisol levels of nor- mal goldfish, Carrassius auratus L., and response to osmotic change. Am. Zool., 11 : 653.
Toften, H., A.M. Arnesen and M. Obling. 2003. Feed intake, growth and ionoregulation in Atlantic salmon (Salmo salar L.) smolts in relation to dietary addition of a feeding stimulant and time of seawater transfer.
Aquaculture, 217 : 647~662.
Woo, N.Y.S. and S.P. Kelly. 1995. Effects of salinity and nutritional status on growth and metabolism of Sparus sarba in a closed seawater system. Aquaculture, 135 : 229~238.
Woo, N.Y.S. and A.C.Y. Fung. 1981. Studies on the biology of red sea bream. 2. Salinity adaptation. Comp. Bio- chem. Physiol., 69A : 237~242.
Zar, J.H. 1984. Biostatistical analysis. 2nd ed. Prentice- Hall, Inc. 718 pp.
Received : February 3, 2004 Accepted : March 12, 2004