서 론
어류 양식에서 사육중인 어체가 주로 받는 스트레스로 는 크게 화학적 요인과 물리적 요인 등으로 나눌 수 있 는데, 이들 중 물리적 요인으로는 염분(Singley and Chavin, 1971), 사육밀도(Wedemeyer and Mcleay, 1981), 수온(Fryer, 1975) 등이 있으며, 화학적 요인으로는 어류 의 기생충 구제제로 사용되는 포르말린(Cho and Yang,
1996) 및 황산동(Varanka et al., 2001)과 같은 소독약제 투여 등이 있다. 그 중 염분 및 수온 등과 같은 환경변화 에서 발생하는 물리적 스트레스는 사육시 성장, 번식, 대 사, 삼투압조절 등 어류의 생리적 변화를 유발시켜 양식 생산성에 큰 영향을 미친다.
최근, 수온의 변화는 연안해역의 대규모 산업 시설인 발전소, 제철공장, LNG 저장기지로부터 냉∙온배수의 연 안해역 방류에 의해서도 수산생물상의 변화 및 수산 자 원량에 영향을 미치는 것으로 보고되고 있으며(Kim and Kim, 2001), 이러한 수온 변화에 의해서 해양생물들은 대 사율이 변하고 동시에 산소 소모량도 변화하는 것으로
─
─ 19 ──
넙치(Paralichthys olivaceus)에서의 급격한 수온변화 스트레스에 관한 생리학적 연구
양 정 환ㆍ여 인 규*
제주대학교 해양과학부
Physiological Studies on Acute Water-temperature Stress of Olive Flounder (Paralichthys olivaceus)
Joung-Hwan Yang and In-Kyu Yeo*
Faculty of Applied Marine Science, Cheju National University, Jeju 690-756, Korea
This study was carried out to investigate variation in blood and antioxidant enzyme activity due to acute changes in water temperature in the flounder (Paralichthys olivaceus). The groups were cultured at 10, 15, 20, 25 and 30�C, for 1, 2, 4, 6 and 8 hours. The hematocrit (Ht), the hemoglobin (Hb) and red blood cell (RBC) concentrations tend to increase when the water tem- perature rises. The cortisol concentration of plasma was decreased at 15�C when the starting temperature was 20�C and recovered to starting level after increased at 10 and 30�C, but was maintained for a longer period at 25�C. The superoxide dismutase (SOD) activity in the kidney increased at high water temperatures (25�C), and decreased at low water temperatures (10 and 15�C). Catalase (CAT) activity in the liver was observed to increased in all groups except at 10
�C. Thus, according to our study, when flounder is cultured at 25�C, high stress remained for a longer period. However, further analysis should be performed involving the physiological effects of temperature over longer periods and at different life stages.
Key words : Olive flounder (paralichthys olivaceus), superoxid dismutase (SOD), catalase (CAT), cortisol, stress, temperature
*Corresponding author: [email protected]
알려져 있다(정과 옥, 1996). 그 중 성장촉진을 위하여 고 수온에 순치된 어류는 저수온에 약한 경향을 나타내는 것으로 알려져 있다(Horning and Pearson, 1973). 또한 고수온에서는 항체 형성시간이 빠르며 항체가도 높게 나 타나는 반면, 저수온에서는 항체 형성이 늦어지고 항체가 도 낮아진다고 보고되고 있다(Avtalion et al., 1976;
Salati et al., 1988; 방과 박, 1994). 이처럼 수온은 어체의 면역 및 기초 생리학적인 변화를 초래하는 주요한 인자 로 여겨지고 있다.
특히, 급격한 수온 변화는 어류에게 있어 스트레스로 작 용하여 어류의 건강도를 저하시켜 질병을 유발시키는 요 인으로 작용한다(Wedemyer and Mcleay, 1981). 또한 여 름철 우리 나라 동해안에서 발생하는 냉수대로 인한 급격 한 저수온 변화는 어류의 성장을 느리게 하고 질병을 발생 시키는 요인으로 알려져 있다(Park, 1978; Schreck, 1981).
일반적으로 스트레스를 받게되면 생체 내에 활성산소 (superoxide, hydrogen peroxide, peroxyl radical, hyd- roxy radical 등)가 발생하는 것으로 알려져 있다. 활성산 소는 생체 내에서 다른 물질과 결합하려는 화학적 친화 력이 강해서 세포나 기관의 막을 공격하여 세포의 기능 을 손상시킨다(Ferraris et al., 2002). 이러한 활성산소에 대하여 체내에서는 항산화 효소인superoxide dismutase (SOD) 및 catalase (CAT) 등을 생성하여 세포기능손상을 막는 것으로 알려져 있다(Chance et al., 1979; Wendel and Feuerstein, 1981).
한편, 이러한 항산화 효소는 온도 변화에 의해 효소활 성이 증가하는 것으로 보고되어 있으며(Parihar et al., 1996; 1997), 중금속 이온의 노출에 의해서도 항산화 효 소 활성이 변화한다고 알려져 있다(Livingstone et al., 1992; Chen et al., 2000; Zikic et al., 2001). 이처럼 항산 화 효소의 활성은 생체내의 스트레스 반응과 밀접한 관 계가 있는 것으로 알려져 있다. 그러나 수온 변화와 같은 물리적 스트레스에 대한 어체내의 항산화 효소 변화에 관한 연구는 아직도 미흡한 실정이다.
따라서 본 연구에서는 넙치(Paralichthys olivaceus)를 이용하여 다양한 수온조건에서의 혈액 성상과 항산화 효 소인 CAT 및 SOD 등의 효소 변화를 조사하여 스트레스 에 대한 생리적인 변화에 대한 지표를 설정하고자 실행 하였다.
재료 및 방법 1. 실험어
실험어는 제주도 남제주군 표선읍에 위치한 육상 수조
식 양식장에서 사육된 넙치(Paralichthys olivaceus)로써 평균 전장의 29.0±1.5 cm, 평균 체중 246.5±25.0 g인 것을 사용하였다. 실험어는 수온 조절이 가능한 순환 여 과식 사육시스템에서 실험 전 2주 동안 예비 사육하여 안정시킨 다음 실험에 사용하였으며, 이때 염분 농도는 30.0±0.5‰, 수온은 20.0±1.0�C이었다. 예비사육 동안 사료는 시판되는 넙치용pellet 사료를 사용하여1일1회 만복 투여하였다. 실험수조는 용량의 1 ton인 FRP 사각 수조로 사용 해수는1차 모래 여과한 다음, 수온조절 제 어시스템에 의해 시간당 10�C씩 하강 또는 상승되도록 하였다. 또한 실험 기간 중 사육수에는 공기공급을 충분 히 하여 용존 산소가 평균5 ppm 이상 유지되도록 하였 다. 실험구는 각각 10, 15, 20 (대조구), 25 및30�C로 설정 하였으며, 각 실험구당 20마리씩 사육하였다. 평균 호흡 수는 1분 동안에 아가미가 움직이는 횟수를 각 그룹당 20개체를3회씩 측정하였다.
2. 혈액 성상 분석
혈액은 실험개시전 넙치를 3일간 절식시킨 다음, 미부 동맥으로부터 헤파린을 처리하지 않은 주사기를 사용하 여 마취 없이 1분 이내에 개체별로 채취하였다. 채취된 혈액은 원심분리용 튜브에 넣어 4�C에서 2시간 방치한 뒤 3,000 rpm에서 20분간 원심 분리하여 혈청을 분리 한 다음 -70�C에서 동결보관 한 후 Glutamic pyruvic transminase (GPT) 및 Glutamic oxalacetic transminase (GOT) 분석에 사용하였다. 또한 헤파린을 처리한 주사기 를 사용하여 개체별로 채취한 혈액은 즉시 헤파린 처리 된 튜브에 분주하여 상온에서20분간 방치한 후, 원심 분 리하여 혈장을 분리하였으며, 추출한 혈장은 -70�C에서 동결보관 한 후 코티졸(cortisol)량을 측정하였다. 적혈구 용적(Ht)은 혈액을 채취한 후 4,000 rpm으로 30분간 원 심분리한 후 적혈구와 혈장부분의 길이에 대한 적혈구 부분의 비율로 구하였다. 혈색소 농도(Hb)는cyanmethe- moglobin법을 이용하여 540 nm에서 측정하였으며, 적혈 구 수(red blood cell count, RBC)는 헤파린 처리한 전혈 을Hayem씨액(Ishisu)으로 희석시켜Thoma 혈구계산판 과 red cell pipet을 사용하여 측정하였다. GPT 및 GOT 효소활성 측정은 GPT의 경우 L-alanine과 α-ketoglu- taric acid, GOT의 경우 L-aspartic acid와 α-ketoglu- taric acid를 기질로 사용하여37�C에서30분간 반응시킬 때 생성되는 pyruvic acid가 알칼리성 하에서 2, 4-dini- trophenyl hydrizine과 작용하여 발색되는 hydrazine의 비색을 정량하는 Reitman-Frankel법(1957)에 의해 조 제된 진단용 kit (아산제약, 한국)를 사용하여 파장 505
nm에서 흡광도를 측정하였다. 코티졸 측정은 cortisol RIA kit (DSL, USA)를 사용하여 항원ㆍ항체반응을 유도 한 다음, Wizard 1470 counter (Hewlett Packard, USA) 를 사용하여radioimmunoassay (RIA)로 분석하였다.
3. 항산화 효소 분석
항산화 효소는 간장과 신장을 적출 하여SOD 및 CAT 를 각각 분석하였다. 간과 신장은 적출 후 약1 g을0.9 % NaCl에3회 세척한 다음KCl (1.17%)을 함유한 100 mM phosphate buffer (pH 7.4)를 첨가하여 균질화 하였다. 균 질화된 시료는 원심분리(1,000 rpm, 15 min, 4�C)에 의해 지방 및 침전물을 제거하였다. 균질액은 다시 원심분리 (13,000 rpm, 20 min, 4�C)한 후 상등액을 항산화 효소 측 정용 시료로 사용하였다.
SOD는 pyrogallol의 자동 산화율이 억제되는 양을 측 정하는 Marklund and Marklund (1974)의 방법으로 측 정하였으며, 50 mM phosphate buffer (pH 8.24) 8.7 ml에 간장 균질액을 넣은 후0.3 ml의3 mM pyrogallol 용액을 첨가하여spectrophotometer를 이용 325 nm에서 측정하 였다. 그리고 효소활성의1단위는 반응액 중의pyrogallol 의 산화를 50% 억제하는 효소의 양으로 정하였다. CAT 활성도의 측정은H2O2를 기질로 사용하여 spectrophoto- meter에 의해 240 nm 파장에서 H2O2가 환원되어 감소 하는 흡광도로서 효소 활성도를 측정하는 Nelson and Kiesow (1972)의 방법에 의하여 측정하였으며, 효소 활성 도의 단위는 1분간에 1 mg의 단백질이 반응하여 환원시 킨 H2O2를 nmol로 나타내었다. 단백질 함량은 Lowry et al. (1951)의 방법에 따라 표준 단백질로서 BSA (bovine serum albumin)를 사용하여 Spectrophotometer를 이용 540 nm에서 측정하였다.
4. 통계 처리
각 실험에서 얻어진 자료에 대한 값의 유의차 유무는 SPSS-통계 패키지에 의한ANOVA로 분석하여Duncan’s multiple range test로 검정하였다.
결 과
1. 평균 호흡수
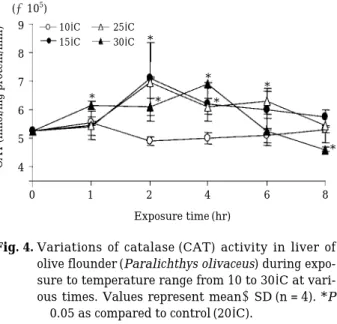
온도에 따른 넙치의 평균 아가미 운동횟수는 대조구 31.7±1.3회/min에 비해 30 및 25�C에서 유의한 증가를 나타내었고, 저온인 15�C에서는 1시간째 39.3±2.8회
/min을 제외한 2, 4, 6, 8시간에는 급속히 낮아졌다(Fig.
1, P⁄0.05). 또한 10�C에서는 대조구와 비교하여 1시간 째41.7±3.1회/min으로 일시적으로 증가하는 경향을 보 였으나 이후 대조구와 비슷한 수치를 유지하였다.
2. 혈액성상 변화
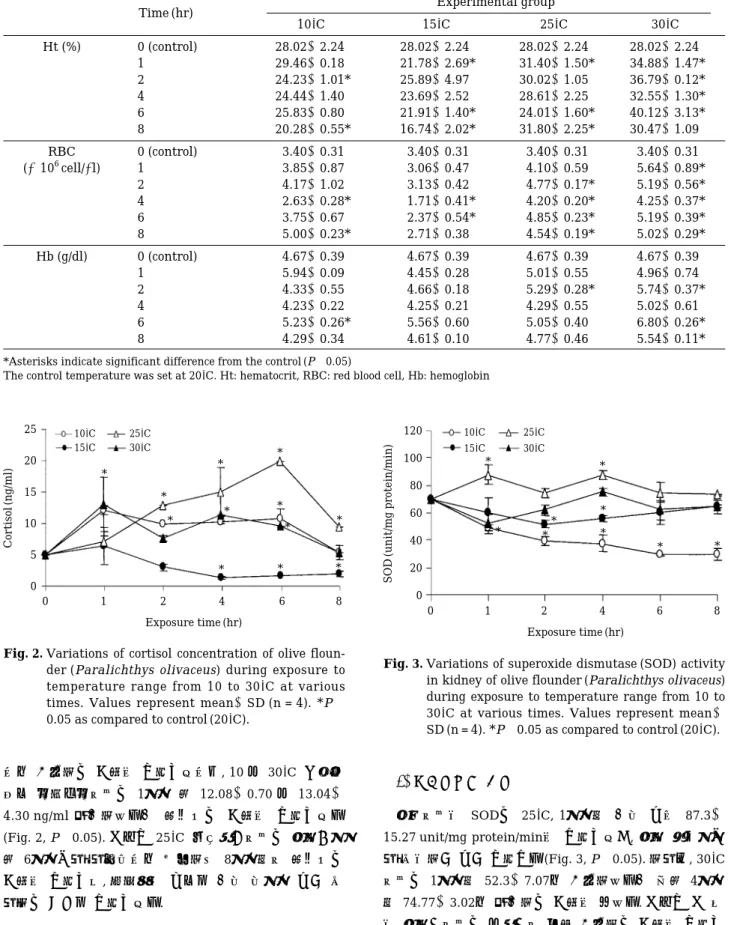
Ht수치는 대조구 28.02±2.24%에 비하여 30�C에서는 전반적으로 높은 수치를 나타내었으며, 6시간째에 40.12
±3.13%로 최대 값을 나타내었다(Table 1, P⁄0.05).
25�C에서는 실험개시 후 4시간까지 전반적으로 높은 값 을 나타내었으나6시간 후에는24.01±1.60%로 낮은 값 을 나타내었다. 그러나 저수온인15�C 및 10�C에서는 대 조구에 비해 유의하게 낮은 값을 나타내었다.
RBC는 실험개시 후 2시간까지는 대조구 3.4±0.3× 106 cell/µl에 비하여 30, 25 및 10�C에서 유의하게 높은 값을 나타내었다(Table 1, P⁄0.05). 그러나 실험개시 후 4시간째 모든 구에서 일시적으로 낮아지는 현상을 보였 다. 반면, 저수온인 15�C 처리 구에서는 1시간 3.06± 0.47, 2시간 3.13±0.42, 4시간 1.71±0.41, 6시간 2.37± 0.54 및 8시간째 2.71±0.38×106 cell/µl로 전 구간에서 대조구보다 낮은 수치를 나타내었다. Hb 농도에 있어서 도 Ht 및 RBC와 유사한 경향을 나타내었다(Table 1, P
⁄0.05).
GOT 및 GPT의 변화는 실험기간 중에 유의적인 차이 를 나타내지 않고, 각각 10.96~11.81 및 11.79~12.00 Karmen의 범위를 나타내었다(data not shown).
수온의 변화에 의해 발생하는 혈장 코티졸의 농도는 15�C에서는 20�C (대조구) 4.92±2.46 ng/ml보다 전반적 Fig. 1. Variations of gill respiration number (No./min.) of olive flounder (Paralichthys olivaceus) during ex- posure to temperature range from 10 to 30�C at various times. Values represent mean±SD (n = 4).
*P⁄0.05 as compared to control (20�C).
0 1 2 4 6 8
Exposure time (hr) 100
80
60
40
20
0
*
*
*
* *
*
*
*
*
*
*
* *
* *
Gill respiration number(No./min.)
10�C 25�C 15�C 30�C
으로 감소하는 경향을 나타내었으며, 10 및 30�C 극심한 온도 스트레스에서는 1시간 후 12.08±0.70 및 13.04±
4.30 ng/ml 증가하였다가 회복되는 경향을 나타내었다
(Fig. 2, P⁄0.05). 그러나25�C 사육조건에서는 실험개시 후 6시간까지 지속적으로 상승하여 8시간째에 회복되는 경향을 나타내어, 코티졸의 농도를 가장 장시간 높게 유 지하는 결과를 나타내었다.
3. 항산화효소 변화
신장에서의 SOD는 25�C, 1시간째 가장 높은 87.3± 15.27 unit/mg protein/min을 나타내었고, 실험 종료시까 지 유의하게 높게 나타났다(Fig. 3, P⁄0.05). 하지만, 30�C 에서는 1시간째 52.3±7.07로 감소하였다가 이후 4시간 째74.77±3.02로 증가하는 경향을 보였다. 그러나 그 외 의 실험구에서는 대조구에 비해 감소하는 경향을 나타내 Table 1. Variations of hematological factors of olive flounder (Paralichthys olivaceus) to exposure of acute water
temperature
Time (hr) Experimental group
10�C 15�C 25�C 30�C
Ht (%) 0 (control) 28.02±2.24 28.02±2.24 28.02±2.24 28.02±2.24
1 29.46±0.18 21.78±2.69* 31.40±1.50* 34.88±1.47*
2 24.23±1.01* 25.89±4.97 30.02±1.05 36.79±0.12*
4 24.44±1.40 23.69±2.52 28.61±2.25 32.55±1.30*
6 25.83±0.80 21.91±1.40* 24.01±1.60* 40.12±3.13*
8 20.28±0.55* 16.74±2.02* 31.80±2.25* 30.47±1.09
RBC 0 (control) 3.40±0.31 3.40±0.31 3.40±0.31 3.40±0.31
(×106 cell/µl) 1 3.85±0.87 3.06±0.47 4.10±0.59 5.64±0.89*
2 4.17±1.02 3.13±0.42 4.77±0.17* 5.19±0.56*
4 2.63±0.28* 1.71±0.41* 4.20±0.20* 4.25±0.37*
6 3.75±0.67 2.37±0.54* 4.85±0.23* 5.19±0.39*
8 5.00±0.23* 2.71±0.38 4.54±0.19* 5.02±0.29*
Hb (g/dl) 0 (control) 4.67±0.39 4.67±0.39 4.67±0.39 4.67±0.39
1 5.94±0.09 4.45±0.28 5.01±0.55 4.96±0.74
2 4.33±0.55 4.66±0.18 5.29±0.28* 5.74±0.37*
4 4.23±0.22 4.25±0.21 4.29±0.55 5.02±0.61
6 5.23±0.26* 5.56±0.60 5.05±0.40 6.80±0.26*
8 4.29±0.34 4.61±0.10 4.77±0.46 5.54±0.11*
*Asterisks indicate significant difference from the control (P⁄0.05)
The control temperature was set at 20�C. Ht: hematocrit, RBC: red blood cell, Hb: hemoglobin
Fig. 2. Variations of cortisol concentration of olive floun- der (Paralichthys olivaceus) during exposure to temperature range from 10 to 30�C at various times. Values represent mean±SD (n = 4). *P⁄
0.05 as compared to control (20�C).
Fig. 3. Variations of superoxide dismutase (SOD) activity in kidney of olive flounder (Paralichthys olivaceus) during exposure to temperature range from 10 to 30�C at various times. Values represent mean±
SD (n = 4). *P⁄0.05 as compared to control (20�C).
0 1 2 4 6 8
Exposure time (hr)
0 1 2 4 6 8
Exposure time (hr) 10�C 25�C
15�C 30�C 25
20 15
10 5
0
Cortisol(ng/ml)
*
*
*
*
*
*
*
*
*
*
*
*
10�C 25�C 15�C 30�C 120
100 80 60 40 20 0
* *
*
* ** *
* *
SOD(unit/mg protein/min)
었으며, 10�C의 실험구에서 가장 낮은 수치를 나타내었 다. 간장에서는 유의한 차이를 나타내지 않았다(data not shown).
한편, 간장에서의 CAT의 활성은 10�C의 실험구를 제 외한 실험구에서 유의하게 높은 수치를 나타내었으며, 10�C에서는 전반적으로 대조구와 유사한 수치를 나타내 었다(Fig. 4, P⁄0.05). 신장에서는 유의한 차이를 나타내 지 않았다(data not shown).
고 찰
일반적으로, 혈액성상에 있어서 Ht, RBC 및 Hb 등은 생체의 산소운반능력을 나타낸다(Chang et al., 1999). 해 산어류에 있어 수온상승 자극은 Ht, RBC 및 Hb는 증가 시키고, 수온하강 자극은 이와 반대의 경향을 나타내는 것으로 보고되고 있다(Ishioka, 1980; Davis et al., 1990;
Chang et al., 1999, 2001). 또한 어체는 스트레스의 작용 으로 혈액 산성화 현상(acidosis)에 의해 적혈구의 산소 운반능력이 떨어지는 것으로 알려져 있다(Perry and Reid, 1993). 이상의 연구 결과에서와 같이 본 연구에서 도 수온상승 시에는 증가되고 수온하강 시에는 감소되는 경향을 나타내었다. 이러한 경향들은 온도가 높아질수록 사육수의 용존산소포화도가 낮아져 이에 따른 산소 부족 으로 어류의 적혈구 수 및Ht수치가 증가하고 활발한 아 가미운동을 통하여 혈액으로의 산소운반을 능동적으로 취하고 있음을 나타내는 것으로 추정된다.
한편, 일반 인체 진단에서 널리 이용되는 혈청 전이 효 소인 GOT와 GPT의 활성은 조직손상 등 어류질병의 진
단에 이용되고 있으며(Shich, 1978; Smith and Ramos, 1980), 일반적으로 장시간의 오염 물질 노출에 의한 스트 레스 작용으로부터 변화하는 것으로 알려져 있다(Casillas and Ames, 1985; Rao et al., 1990). 그러나, 본 연구에서의 급격한 수온 변화 스트레스에서의GOT 및GPT는 전 실 험구간에 있어서 유의한 차이를 나타내지 않았다(data not shown). 이러한 결과는 단기간의 스트레스 조건에서 는 이들 효소의 활성에 큰 영향을 미치지 않을 가능성이 시사되나 보다 세부적인 연구가 진행되어져야 할 것으로 판단된다.
스트레스에 의한 생체내 호르몬 분비는 대부분 시상하 부-뇌하수체-부신피질계의 조절기전에 의존하여 이루어 지는데(Choi and Kim, 1990), 이 기전의 말초 호르몬인 코티졸 함량은 어체가 받는 스트레스 반응의 지표로 널리 이용되고 있다(Donaldson, 1981). 본 연구에서의 코티졸 농도는 20�C보다 15�C에서 낮은 농도를 나타내었으며, 10�C에서는 높아지는 결과를 초래하였다. 그리고25�C 및 30�C에서도10�C와 같은 현상을 보였다. 수온 상승에 의 한 코티졸 농도의 증가는 수온상승에 따른 어체 내로의 불충분한 산소 공급에 기인하는 것으로 보고되고 있다 (Chang et al., 2001). 본 연구에서 30�C 및 25�C에서의 코티졸 농도의 증가는 아가미 운동 횟수 및 혈액성상의 결과로 볼 때 Chang et al (2001) 등과 같이 산소수급의 문제에 기인한 것으로 추정된다. 또한, 15�C 결과는 낮은 수온일수록 낮아진 대사량 및 낮은 에너지 요구량에 기 인한 결과로 Ht, RBC 및 Hb와 동일한 결과를 나타내었 다. 그러나 10�C에서의 코티졸 농도는 수온 하강폭이 커 짐으로 인해 어체에 과도한 스트레스가 작용하여 그 농 도가 증가했을 것으로 생각되어진다.
한편, 스트레스에 의해 발생하는 활성산소는 어떤 유해 물질이나 약물 등에 폭로되었을 때 병적 상태에서는 이 들이 과다하게 생성되어 조직에 치명적인 손상을 입힐 수 있다(Goldberg and Stern, 1977; Simon et al., 1981;
Moody and Hassan, 1982). 이런 생체에서 발생하는 활성 산소에 의한 단백질, 핵산 및 생체막 손상은 SOD 및 CAT 등 항산화 효소에 의해 방어된다. 항산화 효소의 기 작을 보면SOD 및 CAT는 활성산소를 보다 안정된 물질 로 환원시키는데, 그 중 SOD는 superoxide radical (O2-) 을 H2O2와 O2(2O2-±2H±→H2O2±O2)로 전환시키며, 이 전환된 H2O2를 CAT가 O2와 H2O (2H2O2→2H2O±
O2)로 분해하여 활성산소를 형성하기 전에 생체에 무해 한 분자들로 전환시킨다(Forman and Fridovich, 1973).
일반적으로 이러한 항산화 효소는 어류 종, 조직 및 계절 에 따라 항산화 효소 활성이 변화하는 것으로 알려져 있 다(Aksnes and Njaa, 1981; Wdzieczak et al., 1981;
Fig. 4. Variations of catalase (CAT) activity in liver of olive flounder (Paralichthys olivaceus) during expo- sure to temperature range from 10 to 30�C at vari- ous times. Values represent mean±SD (n = 4). *P
⁄0.05 as compared to control (20�C).
0 1 2 4 6 8
Exposure time (hr) 9
8 7 6 5 CAT(nmol/mg protein/min) 4
10�C 25�C 15�C 30�C (×105)
*
*
* *
* *
*
Gabryelak et al., 1983). 또한, Parihar et al. (1996, 1997) 는 온도 증가에 의한 스트레스로 실험 후 4시간까지 SOD 효소활성 등이 증가한다는 연구 보고가 있다. 본 실 험에서도 상기의 연구 결과와 유사한 경향을 나타내었는 데, 수온 상승시2시간 또는4시간 이후까지 항산화 효소 인 SOD 및 CAT가 증가하였다가 이후 실험 종료시까지 낮아지는 경향을 나타내었다. 이러한 결과는 급격한 수온 상승과 수온하강 스트레스에 기인하여 발생하는 활성산 소에 대한 어체의 생리학적 방어 기작으로 인해 SOD 및 CAT가 증가한 결과라고 생각되어지며, 그 후 감소되는 것은 어체내 생리학적 방어기작의 한계에 도달함으로써 어체내 기초 생리학적인 변화를 초래한 결과에 기인하여 항산화 효소 활성이 감소한 것으로 여겨진다.
일반적인 수온 스트레스 조건에서는 수온 상승과 함께 비례적으로 코티졸 및 항산화 효소 활성의 상승한다고 알려져 있다(Davis et al., 1990; Parihar et al., 1996, 1997). 그러나, 본 실험에서는 10 및 30�C에서는 코티졸 및 항산화 효소가 상승한 후 단시간 내에 회복되는 경향 을 나타내었으나, 25�C에서 가장 지속적으로 높은 농도 가 유지되는 결과를 나타내었다. 이상의 결과에서와 같 이, 적정수온 이상의 과도한 수온 변화보다 오히려 적은 수온 변화에 있어서의 스트레스 관련 혈액성상의 변화가 크게 나타나, 생체내의 방어 기작이 적은 수온 변화에 있 어서 그 작용능력이 낮아지게 되고 이에 따른 회복력이 비교적 낮아져 지속적인 스트레스로 작용하는 것으로 판 단된다. 그러나, 이러한 변화에 대한 종합적인 검토를 위 해서는 장기간 또는 단계별 수온 변화에 대한 연구가 앞 으로 검토되어져야 할 것으로 판단된다.
적 요
본 연구에서는 넙치(Paralichthys olivaceus)를 이용하 여 급격한 수온 변화 스트레스에 의한 혈액 및 항산화효 소 변화에 대하여 조사하였다. 실험구는 각각 10, 15, 20 (대조구), 25 및 30�C로 설정하였으며, 측정 시간은 1, 2, 4, 6 및 8시간 후에 측정하였다. 그 결과, 혈액 중에 He- matocrit (Ht), Hemoglobin (Hb) 및 Red blood cell (RBC) 농도는 수온상승 시 증가하였다. 또한, 혈장에서의 코티 졸 농도는 20�C와 비교하여 15�C에서는 감소하였고, 10 및 30�C에서는 상승 후 일정시간이 경과함에 따라 회복 되었으나, 25�C에서는 장시간 코티졸 농도가 높게 유지 되었다. 신장에서의 SOD 활성은 25�C에서 높게 나타났 으며, 10 및 15�C에서는 그 활성이 감소하였다. 간장에서 의 CAT 활성은 10�C를 제외한 실험구에서 유의하게 높
은 수치를 나타내었다. 이러한 결과들로 볼 때, 25�C의 수온에서 사육하였을 때 스트레스 반응이 높게 장시간 지속되는 것으로 판단된다. 하지만, 장기간 또는 단계별 온도의 영향에 대해서 수행되어져야 할 것으로 생각된다.
사 사
이 논문은2003년도 제주대학교 두뇌한국21 사업에 의 하여 연구되었으며, 연구비를 지원하여 주신 데 대하여 깊이 감사드립니다.
인 용 문 헌
Aksnes, A. and L.R. Njaa. 1981. Catalase, Glutathione peroxidase and superoxide dismutase in different fish species. Comp. Biochem. Physiol., 69B : 893~896.
Avtalion, R.R., E. Weiss and T. Moalem. 1976. Regulatory effects of temperature upon immunity in ectothermic veterbrate. In comparative immunology. Marchalonis, J., Blachwell Scientific Publication, Oxford, pp. 227~ 238.
Casillas, E. and W. Ames. 1985. Serum chemistry of diseased English sole, Parophrys vetulus Girard, from polluted areas of Puget Sound, Washington. J. Fish Dis., 8(5) : 437~449.
Chance, B., H. Siec and A. Boveris. 1979. Hydroperoxide metabolism in mammalian organs. Physiol, Rev., 59 : 527~605.
Chang, Y.J., J.W. Hur, H.K. Lim and J.K. Lee. 2001. Stress in olive flounder (Paralichthys olivaceus) and fat cod (Hexagrammos otakii) by the sudden drop and rise of water temperature. J. Korean Fish. Soc., 34 : 91~97 (in Korean).
Chang, Y.J., M.R. Park, D.Y. Kang and B.K. Lee. 1999.
Physiological responses of cultured olive flounder (Par- alichthys olivaceus) on series of lowering seawater temperature sharply and continuously. J. Korean Fish.
Soc., 32(5) : 601~606 (in Korean).
Chen, Y., X.D. Cao, Y. Lu and X.R. Wang. 2000. Effects of rare earth metal ions and their EDTA complexes on antioxidant enzymes of fish liver. Environ. Contam.
Toxicol., 65 : 357~365.
Cho, J.K. and H.C. Yang. 1996. Determination of formal- dehyde residue and histopathological observation in formalin and neutral-formalin treated Korean rockfish (Sebastes schlegeli). J. Fish Pathol., 9 (2) : 157~168.
Choi, J.H. and H.R. Kim. 1990. The effects of tryptophan
and tyrosine-enriched diet on the serum cortisol, glu- cose and free fatty acid levels of stressed rats. Korean J Nutrition., 23(4) : 229~236.
Davis, K.B., P. Torrance and N.C. Parker. 1990. Physio- logical stress in striped bass: effect of acclimation temperature. Aqaculture, 91 : 349~358.
Donaldson, E.M. 1981. In Stress in Fish A. D. Pickering (ed.), Academic Press, London, 11 pp.
Forman, H.J. and I. Fridovich. 1973. Superoxide dismutase:
A comparison of rate constant. Arch. Biochem. Bio- phys., 158 : 396.
Fryer, J.N. 1975. Stress and adrenocorticosteroid dynamics in the goldfish, Carassiu auratus. Can. J. Zool., 53 : 1011~1020.
Gabryelak, T., M. Piatrowska, W. Leyko and G. Peres.
1983. Seasonal variation in the activities of peroxide metabolism enzymes in erythrocytes of freshwater fish species. Comp. Biochem. Physiol., 75C : 383~385.
Goldberg, B. and A. Stern. 1977. The role of the superoxide anion as a toxic species in the erythrocyte. Arch. Bio- chem. Biophys., 178 : 218~225.
Horning, W.B.I. and R.E. Pearson. 1973. Growth tempera- ture requirement And lower lethal temperature for juvenile smallmouth bass (Micropterus dolomieui). J.
Fish. Res. Bd. Can., 30 : 1226~1230.
Ishioka, H. 1980. Stress reactions in the marine fish-I Stress reactions induced by temperature change. Bull Jpn Soc. Sci. Fish., 46 : 523~532 (in Japanese).
Kim, J.K. and J.H. Kim. 2001. Diffusion modelling of cold discharge on coastal waters. Jour. Fish. Mar. Sci. Edu., 13(2) : 132~145 (in Korean).
Livingstone, D.R., S. Archibald, J.K. Chipman and J.W.
Marsh. 1992. Antioxidant enzymes in liver of dab Lim- anda limanda from the North Sea. Mar. Ecol. Prog.
Ser., 91 : 97~104.
Lowry, O.H., N.J. Rosenbrought, A.L. Farr and R.J. Randall.
1951. Protein measurement with the Folin-phenol reagent. J. Biol. Chem., 193 : 265~275.
Marklund, S. and G. Marklund. 1974. Involvement of the superoxide anion radical in the antioxidation of pyro- gallol and a convenient assay for superoxide dismutase.
Eur. J. Biochem., 47 : 469~474.
Moody, C.S. and H.M. Hassan. 1982. Mutagenicity of oxy- gen free radicals. Proc. Natl. Acad. Sci., 79 : 2855~2859.
Nelson, D.P. and L.A. Kiesow. 1972. Enthalpy of decompo- sition of hydrogen peroxide by catalase at 25�C (with molar extinction coefficients of H2O2solutions in the UV). Anal Biochem., 49 : 474~478.
Park, C.K. 1978. Chemical oceanographic aspect of the cold water mass in offshore of the east coast of Korea.
Korean Fish. Soc., 11(2) : 49~54 (in Korean).
Parihar, M.S., A.K. Dubey, T. Javeri and P. Prakash. 1996.
Changs in lipid peroxidation, superoxide dismutase activity, ascorbic acid and phospholipid content in liver of freshwater catfish heteropneustes fossilis exposed to elevated temperature. J. Therm. Biol., 21(5/6) : 323~ 330.
Parihar, M.S., T. Javeri, T. Hemnani, A.K. Dubey and P.
Prakash. 1997. Responses of superoxide dismutase, glutathione peroxidase and reduced glutathione antio- xidant defenses in gills of the freshwater catfish (Het- eropneustes fossilis) to short-term elevated tempera- ture. J. Therm. Biol., 22 : 151~156.
Perry, S.F and S.D. Reid. 1993. β-adrenergic signal trans- duction in fish interactive effects of catecholamines and cortisol. Fish. Physiol. Biochem., 11 : 195~203.
Rao, P.P., K.V. Joseph and K.J. Rao. 1990. Histopathologi- cal and biochemical changes in the liver of a fresh water fish exposed to heptachlor. J. Nat. Conserv., 2(2) : 133~137.
Reitman, S. and S. Frankel. 1957. A colorimetric method for the determination of serum glutamic oxaloacetic and glutamic pyruvic transaminase. Am. J. Clin. Pathol., 28 : 56.
Salati, F., K. Kawai and R. Kusuda. 1988. Immune res- ponse of ayu against Vibro anguillarum lipopolysac- charide. Nippon Suisan Gakkaishi., 55(1) : 45~49.
Schreck, C.B. 1981. Stress and compensation in teleostean fishes: responses to social and physical factors. In Stress in fish, A.S. Pickering. Ed. Academic Press, London., 295~321 pp.
Shich, M.S. 1978. Changes of blood enzymes in brook trout induced by infection with Aeromonas salmonicida. J.
Fish Biol., 11 : 13~18.
Simon, R.H., C.H. Scoggin and D. Patterson. 1981. Hydro- gen peroxide causes the fatal injury to human fibro- blasts exposed to oxygen radicals. J. Biol. Chem., 256 : 7181~7186.
Singley, J.A. and W. chavin. 1971. Cortisol levels of normal goldfish, Carassius auratus L. and response to osmotic change. Am. Zool., 11 : 653.
Smith, A.C. and F. Ramos. 1980. Automated chemical an- alysis in fish health assessment. J. Fish Biol., 17 : 445
~450.
Varanka, Z., I. Rojik. I. Varanka, J. Nemcsok and M.
Abraham. 2001. Biochemical and morphological chang- es in carp (Cyprinus carpio L.) liver following exposure to copper sulfate and tannic acid. Comp. Biochem.
Physiol., 128C : 467~478.
Wdzieczak, J., G. Zalesna., A. Bartowiak, H. Witas and W.
Leyko. 1981. Comparative studies on superoxide dis- mutase, catalase and peroxidase level in erythrocytes of different fish species. Comp. Biochem. Physiol., 68B : 357~358.
Wedemeyer, G.A. and D.J. Mcleay. 1981. Methods for determining The tolerance of fishes to environmental stressors. In Stress and Fish A.D. Pickering (ed.), Academic Press, London, pp. 247~275.
Wendel, A. and S. Feuerstein. 1981. Drug-induced lipid peroxidation in mice-1. Modulation by monoxygenase
activity, glutathione and selenium status. Biochem Pharmacol., 30 : 2513~2520.
Zikic, R.V., A.S. Stajn, S.Z. Pavlovic, B.I. Ognjanovic and Z.
S. Saicic. 2001. Activities of superoxide dismutase and catalase in erythrocytes and plasma transaminases of goldfish (Carassius auratus gibelio Bloch.) exposed to cadmium. Physiol. Res., 50 : 105~111.
방종득ㆍ박수일. 1994. 넙치의 면역반응에 수온이 미치는 영 향. 한국어병학회, 7(2) : 119~126.
정 용ㆍ옥치상. 1996. 인간과 환경. 지구문화사.
Received : January 10, 2004 Accepted : March 5, 2004