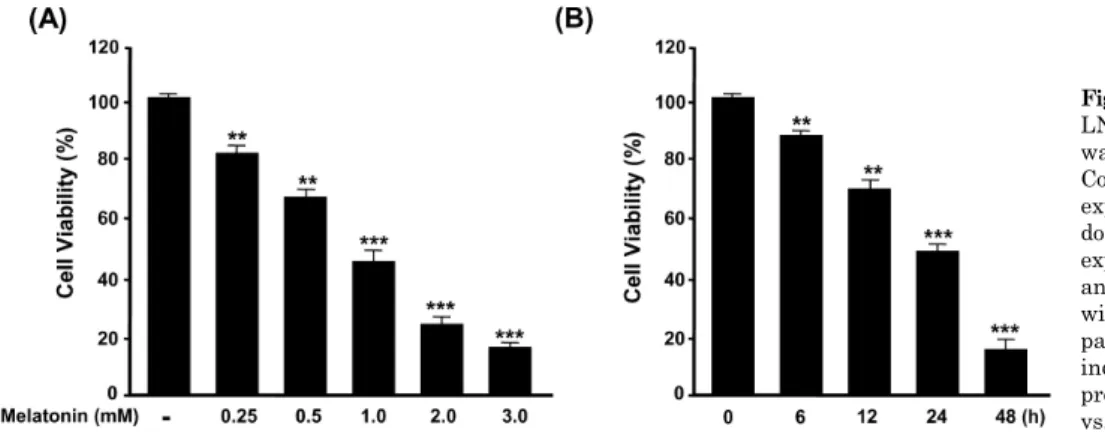
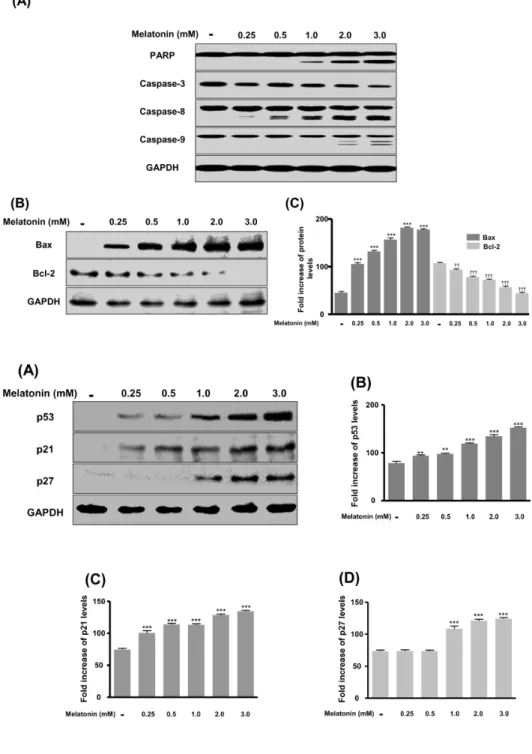
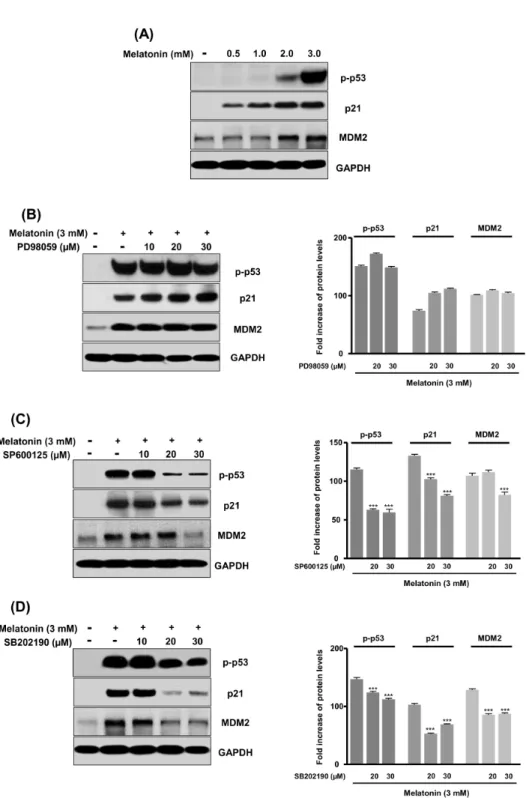
Melatonin Induces Apoptotic Cell Death via p53 in LNCaP Cells
5
0
0
전체 글
(2)
(3)
(4)
(5)
수치
관련 문서
In conclusion, PD induces the cell death via the inhibition of MAPK/ERK pathway in the human cervical cancer cell line, HeLa S 3 cells.. The inhibition of MAPK/ERK pathway
Arylquin 1 induces morphological changes (cell body shrinkage and cell detachment) and decreases cell viability in various cancer cells.. Arylquin 1-induced cell death is
We demonstrated that ionizing radiation utilizes PI3K-JNK signaling pathway for amplifying mitochondrial dysfunction and susequent apoptotic cell death: We showed
We found that [6]-gingerol inhibited the cell growth, disrupted the cell cycle progression in both HPac cells (wild type p53) and BXPC-3 cells (mutant p53 protein), and also
Our results indicate that the activation of p38 and JNK, but not ERK, is involved in the induction of pip92 during NMDA-mediated neuronal cell death, suggesting that different
Conclusions: The study suggests possibilities that the blue-light promotes retinal damage and causes apoptotic cell death via ERK and SRC pathway in mouse retina, and blue-light
In addition, to examine whether HoxB4 affects cell death induced by cell cycle arrest, we treated Ba/F3-GFP and Ba/F3-HoxB4 cells with 100 nM DOX for 24 h, and then
Taken together, we found curcumin-induced ROS down-regulation of E2F4 expression and modulation of E2F4 target genes which finally lead to the apoptotic cell death in HCT116
