265 http://dx.doi.org/10.4196/kjpp.2012.16.4.265
ABBREVIATIONS: HSCs, hematopoietic stem cells; DOX, doxoru- bicin; IL-3, interleukin-3; GFP, green fluorescent protein; FACS, fluorescence-activated cell sorting; TNF-α, tumor necrosis factor-α.
Received May 12, 2012, Revised July 4, 2012, Accepted July 19, 2012
*Corresponding to: Dong-Ku Kim, Department of Animal Biotechno- logy, Konkuk University, 1, Hwayang-dong, Gwangjin-gu, Seoul 143-701, Korea. (Tel) 82-2-450-3743, (Fax) 82-2-455-1044, (E-mail) [email protected]
†
Co-corresponding to: Jin-Hoi Kim, Department of Animal Biotech- nology, Konkuk University, 1, Hwayang-dong, Gwangjin-gu, Seoul 143-701, Korea. (Tel) 82-2-450-3687, (Fax) 82-2-455-1044, (E-mail) [email protected]
‡
These authors contributed equally to this work.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://
creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
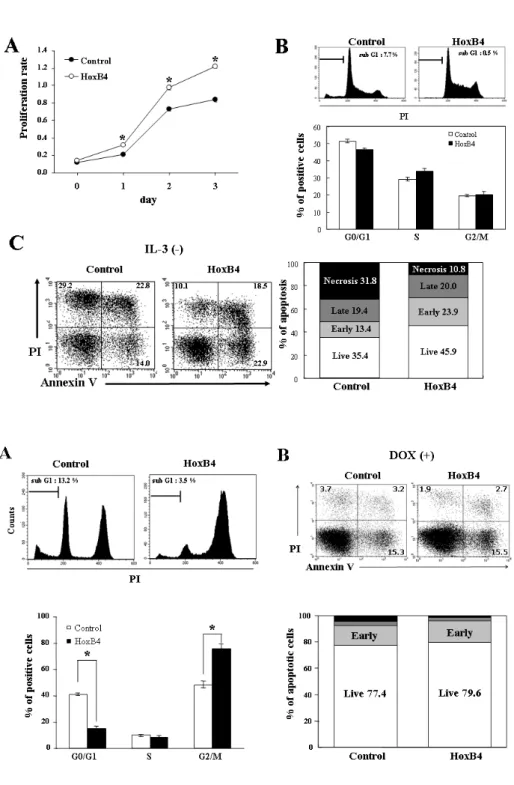
Increased HoxB4 Inhibits Apoptotic Cell Death in Pro-B Cells
Sung-Won Park
1,‡, Kyung-Jong Won
2,‡, Yong-Soo Lee
3, Hye Sun Kim
4, Yu-Kyung Kim
1, Hyeon-Woo Lee
4, Bokyung Kim
2, Byeong Han Lee
5, Jin-Hoi Kim
6,†, and Dong-Ku Kim
6,*
1
Department of Biomedical Science, College of Life Science, CHA University, Seongnam 463-712,
2Department of Physiology and Medical Science, School of Medicine, Konkuk University, Chungju 380-701,
3Transplantation Research Center, Samsung Biomedical Research Institute, Seoul 135-710,
4Department of Pharmacology, Institute of Oral Biology, School of Dentistry, Kyung Hee University, Seoul 130-701,
5