311 서 론
세포 성장의 기본적 진행에 해당되는 세포주기는 G1, S, G2 및 M기로 구분이 되며 cyclin dependent kinases (Cdks)와 cyclin의 catalytic complex의 활성 여부에 의하여
조절된다.1,2) G1기는 cyclin D/Cdk4 또는 Cdk6, 그리고 cyclin E/Cdk2에 의하여 조절되고 S기 동안은 cyclin A/
Cdk2에 의하여 활성화된 DNA polymerase α에 의해 염색 체가 복제된다. 또한 G2/M기는 cyclin A/Cdc2 및 cyclin B/Cdc2에 의해 조절된다.3,4) 이들 중 G2/M기에 관여하는 Cdc2는 Schizosaccharomyces pombe에서 최초로 발견되었으며
인체위암 및 흑색종세포에서 Genistein에 의한 세포주기 G2/M Arrest 유발에 관한 연구
1부산대학교 자연과학대학 생물학과, 2동의대학교 한의과대학 생화학교실 및 대학원 바이오물질제어학과
최우영1․박 철1,2․김경미1․한민호1․최영현2․이원호1
Induction of G2/M Arrest by Genistein in Human AGS Gastric Carcinoma and SK-MEL-2 Melanoma Cells
Woo Yung Choi1, Cheol Park1,2, Kyung Mi Kim1, Min Ho Han1, Yung Hyun Choi2 and Won Ho Lee1
1Department of Biology (BK21 Program), College of Natural Science, Pusan National University, Buan 609-735,
2Department of Biochemistry, Dongeui University College of Oriental Medicine and Department of Biomaterial Control (BK21 Program), Dongeui University Graduate School, Busan 614-052, Korea
Genistein, a natural isoflavonoid phytoestrogen, is a strong inhibitor of protein tyrosine kinase and DNA topoisomerase II activities. Genistein has been shown to have anticancer proliferation, dif- ferentiation and chemopreventive effects. In the present study, we investigated the mechanism of action by which genistein suppressed the proliferation of AGS human gastric carcinoma and SK-MEL-2 melanoma cells. Treatment of AGS and SK-MEL-2 cells to genistein resulted in the growth inhibition and morphological change in a dose-dependent manner. Flow cytometric analysis revealed that genistein treatment caused G2/M phase arrest of the cell cycle, which was associated with a down-regulation of cyclin A, cyclin B1 and cyclin-dependent kinase (Cdk) 2. And we also observed down-regulation of Cdc25C, which was a marker of cell proliferation, and plays important role in cyclin B-Cdc2 complex activation, however Wee1 kinase expression was not affected. Furthermore, genistein induced Cdk inhibitor p21 (WAF1/CIP1) expression through p53-independent manner in AGS cells, but not affect in SK-MEL-2 cells. Taken together, these data provide strong molecular evidence for the anti-tumor activity of genistein in AGS human gastric carcinoma and SK-MEL-2 melanoma cells. (Cancer Prev Res 11, 311-320, 2006)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: Genistein, AGS, SK-MEL-2, G2/M arrest, p21
책임저자:이원호, ꂕ 609-735, 부산광역시 금정구 장전동 30 부산대학교 자연과학대학 생물학과
Tel: 051-510-2257, Fax: 051-517-2980 E-mail: [email protected]
접수일:2006년 10월 2일, 게재승인일:2006년 10월 25일
Correspondence to:Won Ho Lee
Department of Biology, College of Natural Science, Pusan National University, 30, Jangjeon-dong, Geumjeong-gu, Buan 609-735, Korea Tel: +82-51-510-2257, Fax: +82-51-517-2980
E-mail: [email protected]
cyclin B1과 heterodimeric complex를 형성하는데 Wee1 kinase에 의하여 Cdc2의 ATP-binding domain인 Thr-14 및 Tyr-15 잔기의 인산화에 의해 비활성 형태로 유지되고5,6) Cdc25C에 의하여 이들 잔기의 탈인산화가 일어나면 활 성형태로 전환된다.7) 이러한 탈인산화와 활성화는 유사 분열의 개시에 매우 중요하다.
세포성장의 억제는 oxidative stress 또는 DNA damaging agent 등과 같은 세포 내․외부 환경 변화에 의하여 일어 날 수 있는데, 이러한 stress는 G1 또는 G2/M기 특이적인 arrest를 유도할 수 있다. 특히 세포주기의 진행을 억제하 는 것으로 알려진 tumor suppressor protein인 p53은 tran- scription factor로서 cyclin과 Cdk의 발현을 억제하고 Cdk inhibitor인 p21의 발현을 직접적으로 유도한다. 하지만 어떤 경우에는 p53은 anti-apoptotic protein인 Bcl-2의 억제 및 pro-apoptotic protein인 Bax와 Bid의 증가에 의해서 유 도되는 apoptotic death를 유발하기도 한다.8,9)
Genistein (4',5,7-trihydroxyisoflavone)은 대두에 풍부하게 존재하는 phytoestrogen isoflavone이다.10∼12) Genistein은 Pseudomonas sp.에서 최초로 분리된 estrogen과 비슷한 heterocyclic diphenolic 구조이고13∼15) protein tyrosine kinase (PTK), DNA topoisomerase I과 II, protein histidine kinase 및 ribosomal S6 kinase를 억제하며16∼18) anti-estrogenicity, anti- oxidant activity, anti-angiogenesis activity 등을 가지는 것으 로 알려져 있다.19∼21) Genistein은 또한 생쥐의 배 발생 단 계의 성장 및 세포증식을 억제하거나 세포분화를 유도 하고22) 유방암, 폐암, 전립선암 및 혈구암 등과 같은 여 러 종류의 인체 암세포에서 세포의 증식을 억제하고 apoptosis를 유발하는 것으로 보고되고 있다.23∼28) 하지만
인체위암세포 및 흑색종세포에서의 증식억제기전 및 분 자적 표적인자에 대해서는 아직 잘 알려져 있지 않다.
본 연구에서는 genistein의 효능에 관하여 비교적 조사 가 이루어지지 않은 AGS 인체위암세포 및 SK-MEL-2 흑 색종세포를 대상으로 세포의 증식억제에 관여하는 주요 유전자들의 발현 변화를 조사하였다.
재료 및 방법 1. 실험재료
본 실험에 사용된 genistein은 Sigma Chemical Co. (St.
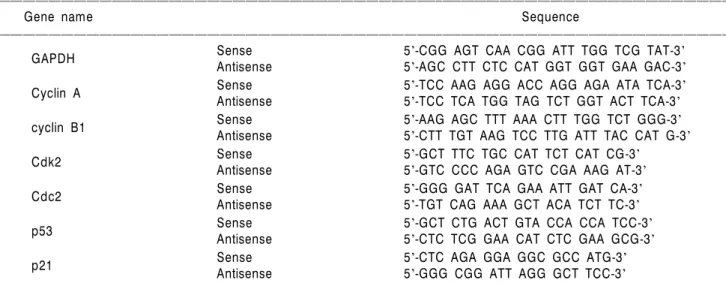
Louis, MO, USA)에서 구입하였고 세포주기 분석을 위하 여 사용된 CycleTEST PLUS DNA REAGENT Kit는 Becton Dickinson (Mississauga, Ontario, CANADA)에서 구입하였 다. mRNA 분석을 위하여 Bioneer (Taejeon, Korea)에서 구 입한 primer는 Table 1에 나타내었고 단백질 분석을 위하 여 사용된 cyclin A, p53 및 p21 항체는 CalBiochem (San Diego, CA, USA), cyclin B1, Cdk2, Cdc2, Cdc25C 및 Wee1 항체는 Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA), phospho-Cdc2 및 phospho-Cdc25C 항체는 Cell Signalling Technology Inc. (Danvers, MA, USA)에서 구입하였다. Im- munoblotting을 위해 2차 항체로 사용된 peroxidase-labeled donkey anti-rabbit 및 peroxidase-labeled sheep anti-mouse immunoglobulin은 Amersham Life Science Corp. (Arlington Heights, IL, USA)에서 구입하였다.
2. 암세포의 배양 및 시료의 처리
실험에 사용한 AGS 인체위암세포 및 SK-MEL-2 흑색
Table 1. Sequence of primers used for RT-PCR
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Gene name Sequence
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Sense 5'-CGG AGT CAA CGG ATT TGG TCG TAT-3'
GAPDH Antisense 5'-AGC CTT CTC CAT GGT GGT GAA GAC-3'
Sense 5'-TCC AAG AGG ACC AGG AGA ATA TCA-3'
Cyclin A Antisense 5'-TCC TCA TGG TAG TCT GGT ACT TCA-3'
Sense 5'-AAG AGC TTT AAA CTT TGG TCT GGG-3'
cyclin B1 Antisense 5'-CTT TGT AAG TCC TTG ATT TAC CAT G-3'
Sense 5'-GCT TTC TGC CAT TCT CAT CG-3'
Cdk2 Antisense 5'-GTC CCC AGA GTC CGA AAG AT-3'
Sense 5'-GGG GAT TCA GAA ATT GAT CA-3'
Cdc2 Antisense 5'-TGT CAG AAA GCT ACA TCT TC-3'
Sense 5'-GCT CTG ACT GTA CCA CCA TCC-3'
p53 Antisense 5'-CTC TCG GAA CAT CTC GAA GCG-3'
Sense 5'-CTC AGA GGA GGC GCC ATG-3'
p21 Antisense 5'-GGG CGG ATT AGG GCT TCC-3'
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
종세포는 생명공학연구소(KRIBB, Taejeon, Korea)에서 분 양 받았으며 90%의 RPMI-1640 배지(Gibco BRL, Grand Island, NY, USA), 10% fetal bovine serum (FBS, Gibco BRL) 에 1%의 penicillin 및 streptomycin (Gibco BRL)이 포함된 성장배지를 사용하여 5% CO2, 37oC의 조건하에서 배양 하였다. Genistein은 dimethyl sulfoxide (DMSO)에 녹여 100 mM의 stock 용액으로 제조한 뒤 -20oC에 보관하였고 적 정 농도로 배지에 희석하여 처리하였다.
3. 세포형태의 관찰
세포배양용 페트리 접시에 적정 수의 세포를 분주하 여 24시간 동안 안정화시킨 다음 genistein을 농도별로 처 리하여 48시간 동안 배양한 후, 도립 현미경(Carl Zeiss, Germany)을 이용하여 200배의 배율로 각 농도에 따른 세 포의 형태변화를 관찰하였다.
4. MTT assay를 이용한 세포 성장억제 조사
세포배양용 6 well plate에 well 당 1×105개의 암세포를 분주하고 24시간 동안 안정화시킨 다음 genistein을 배지 에 희석하였다. 48시간 후 배지를 제거하고 tetrazolium bromide salt (MTT, Sigma) 시약을 0.5 mg/ml 농도로 희석 하여 200μl씩 분주하고 3시간 동안 배양하였다. 배양이 끝난 다음 MTT 시약을 제거하고 DMSO를 100μl씩 분주 하여 well에 생성된 formazin을 모두 녹인 후 ELISA reader (Molecular Devices, Sunnyvale, CA, USA)로 540 nm에서 흡 광도를 측정하였다. 측정은 모두 3회 실시하였으며, 그 에 대한 평균값과 표준 오차를 Microsoft EXCEL program 을 사용하여 분석하였다.
5. Flow cytometry를 이용한 세포주기 분석
정상 및 genistein이 함유된 배지에서 자란 암세포들을 PBS로 두세 번 씻어내고, CycleTEST PLUS DNA REA- GENT Kit를 이용하여 고정 및 염색을 하여 4oC, 암실에 서 30분 동안 반응시킨 다음 DNA flow cytometry (Becton Dickinson, San Jose, CA, USA)에 적용시켜 형광반응에 따 른 histogram을 ModiFit LT (Becton Dickinson) 프로그램으 로 분석하였다.
6. Reverse transcription-polymerase chain re- action (RT-PCR) 분석
동일한 조건에서 준비된 세포들을 대상으로 TRIzol B (Invitrogen, Carlsbad, CA, USA)를 이용하여 total RNA를 분 리하였다. 분리된 RNA를 정량한 후, ONE-STEP RT-PCR PreMix (iNtRON BIOTECHNOLOGY, Korea)를 이용하여 2
μg의 RNA에서 ss cDNA를 합성하였다. 선행연구 조건에 준하여 이 cDNA를 template로 사용하여 관찰 대상 유전 자(Table 1)를 polymerase chain reaction (PCR) 방법으로 증 폭하였다.29) 이때 housekeeping 유전자인 glyceraldehyde- 3-phosphate dehydrogenase (GAPDH) 유전자를 internal con- trol로 사용하였다. 각 PCR 산물들을 1% agarose gel을 이 용하여 전기영동하고 ethidium bromide (EtBr)로 염색한 후 UV 하에서 확인하였다.
7. 단백질의 분리, 전기영동 및 Western blotting
정상 및 genistein이 처리된 배지에서 자란 세포들을 lysis buffer로 용해한 후, 고속원심분리로 세포 내 잔사물 을 분리시킨 후 동량의 단백질을 SDS-polyacrylamide gel 전기영동으로 분리하였다. 분리된 단백질을 nitrocellulose membrane (Schleicher and Schuell, Keene, NH, USA)으로 전 이시킨 후, 특정 단백질에 대한 항체와 그에 대한 이차 항체 반응을 실시한 후 enhanced chemiluminoesence (ECL) 용액(Amersham Life Science)을 적용시킨 다음 X-ray film에 감광시켜 특정 단백질의 양을 분석하였다.
결과 및 고찰
1. 암세포의 형태에 미치는 genistein의 영향
Genistein 처리에 의한 AGS 인체위암세포 및 SK-MEL-2 흑색종세포의 형태 변화 관찰을 위하여 다양한 농도 범 위로 48시간 동안 처리한 후 위상차 현미경을 이용하여 정상 및 genistein이 처리된 조건에서 배양된 폐암세포의 모양을 관찰하였다. Fig. 1A 및 B에서 볼 수 있듯이 genistein의 농도 증가에 따라 두 세포주 모두에서 세포의 심한 형태적 변형이 관찰되었는데, 처리된 genistein의 농 도가 증가될수록 전체적으로 세포질이 응축되면서 배양 접시의 바닥에 낮게 부착되는 듯한 모습을 보였다. 또한 세포의 모양이 길어지면서 돌기와 같은 형태가 분지를 이루기 시작하였고, 고농도 처리 군에서는 많은 암세포 들이 부착 능력이 상실되어 배지 위로 부유하기 시작하 였으며 배양 접시에 부착된 세포들도 심한 형태적 변이 를 수반하였다.
2. 암세포의 증식에 미치는 genistein의 영향
AGS 인체위암세포 및 SK-MEL-2 흑색종세포의 증식에 미치는 genistein의 영향을 알아보기 위하여 genistein을 48 시간 동안 처리한 후 MTT assay를 이용하여 조사한 결과, Fig. 2A 및 B에서 나타난 바와 같이 두 세포주 모두에서 genistein의 농도가 증가함에 따라 암세포의 증식이 감소
하는 것을 알 수 있었다. 특히 SK-MEL-2 흑색종세포에서 는 genistein을 40μM 처리하였을 경우 약 60%의 증식억 제 효과가 나타난 반면 AGS 인체위암세포에서 약 70%
의 증식억제 효과가 나타나 AGS 세포가 genistein에 대해 조금 더 민감하게 반응하는 것을 알 수 있었다.
3. 세포주기 분포에 미치는 genistein의 영향
세포주기 조절의 관점에서 암세포는 세포주기의 비정 상화에 따른 질병으로 정의될 수 있으며, 특정 시기의
세포주기 억제는 세포주기 조절인자의 발현 및 활성여 부에 따라 조절될 수 있다.30,31) 따라서 genistein의 처리에 의한 AGS 인체위암세포 및 SK-MEL-2 흑색종세포의 증 식억제가 세포주기 특정 시기의 진행억제와 연관성이 있는지를 알아보기 위하여 flow cytometry를 이용하여 분 석하였다. Fig. 3 및 Table 2에 나타난 바와 같이 AGS 인 체위암세포의 경우 정상배지에서 자란 암세포의 경우 G1, S 및 G2/M기에 해당하는 세포의 빈도는 약 38.69%, 28.50% 및 32.81% 정도였으나 40μM 처리군의 경우 G1 Fig. 1. Effects of genistein on the morphology of (A) AGS human gastric carcinoma and (B) SK-MEL-2 melanoma cells. Cells were incubated with variable concentrations of genistein. After 48 h incubation, cells were sampled and examined under light microscopy (original magnification, ×200).
0
A
10 20 30 40
AGS
Genistein ( M)µ
0 10 20 30 40
Genistein ( M)µ
B SK-MEL-2
Fig. 2. Anti-proliferative effects of genistein treatment in (A) AGS human gastric carcinoma and (B) SK-MEL-2 melanoma cells.
Cells were treated with variable concentrations of genistein. After 48 h incubation, MTT assay was performed. Results are expressed as average from two separate experiments.
및 S기에 해당하는 세포의 빈도는 약 33.13% 및 8.95%로 감소한 반면 G2/M기에 해당하는 세포의 빈도는 약 57.91%로 증가하였다. SK-MEL-2 흑색종세포의 경우도
G1, S 및 G2/M기에 해당하는 세포의 빈도는 약 48.60%, 20.95% 및 30.76% 정도였으나 genistein의 농도가 증가할 수록 G1 및 S기에 해당하는 세포의 빈도가 감소하고 G2/M기에 해당하는 세포의 빈도가 증가하여 40μM 처 리군의 경우 G1 및 S기는 약 4.61% 및 6.98%로 나타난 반면 G2/M기의 경우는 약 88.40%로 나타났다. 이상의 결과를 살펴볼 때 두 세포주 모두에서 genistein에 의한 증식 억제는 G2/M기에서의 세포주기 억제와 연관성이 있었다. 특히 SK-MEL-2 흑색종세포가 AGS 인체위암세 포에 비해서 G2/M기에서의 세포주기 억제가 더욱 더 강 하게 나타나는 것을 알 수 있었다.
4. Cyclin 및 Cdk의 발현에 미치는 genistein의 영향
일반적으로 진핵세포에서 세포주기는 protein kinase인 Cdks의 활성화에 의하여 조절된다.32∼34) 활성화된 Cdks 는 특정 cyclin과 특이적인 결합에 의하여 cyclin/Cdk com- plex를 형성하여 세포주기의 진행을 조절한다.35) S기에서 G2기로의 진입은 cyclin A/Cdk2 complex가 관여하고 G2기 에서 M기로의 진입에는 B-type cyclin과 Cdc2가 결합하여 Fig. 3. Effects of genistein on cell cycle distribution of (A) AGS human gastric carcinoma and (B) SK-MEL-2 melanoma cells.
Exponentially growing cells have been grown in variable concentrations of genistein for 48 h, then analyzed by flow cytometry as described in Materials and Methods. The data representative example for duplicate tests.
Cell number
DNA content G1
S G2/M
0 10 20 20 40
Sub-G1
A.AGS
Genistein ( M)µ
B.SK-MEL-2
Table 2. Fractions of each cell cycle phase of AGS human gastric carcinoma and SK-MEL-2 melanoma cells cultured in the presence or absence of various concentrations of genistein ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
% of cells Cell Genistein
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
line (μM) G1 S G2/M
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
AGS 0 38.69 28.50 32.81
10 46.09 27.31 26.59
20 48.88 23.35 27.76
30 50.47 15.53 34.01
40 33.13 8.95 57.91
SK-MEL-2 0 48.60 20.65 30.76
10 47.50 18.86 33.65
20 29.60 14.75 55.65
30 11.24 8.59 80.17
40 4.61 6.98 88.40
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Fig. 4. Effects of genistein treatment on the cyclin A and cyclin B1 expression in AGS human gastric carcinoma and SK-MEL-2 melanoma cells. (A) After 48 h incubation with genistein, total RNAs were isolated and reverse- transcribed. The resulting cDNAs were subjected to PCR with indicated primers and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
0
A
10 20 30 40
Genistein ( M)µ
0 10 20 30 40
Genistein ( M)µ
Cyclin A
Cyclin B1
GAPDH
0
B
10 20 30 40
Genistein ( M)µ
0 10 20 30 40
Genistein ( M)µ
Cyclin A
Cyclin B1
Actin
AGS SK-MEL-2
Fig. 5. Effects of genistein treatment on the Cdk2 and Cdc2 expression in AGS human gastric carcinoma and SK-MEL-2 melanoma cells. (A) After 48 h incubation with genistein, total RNAs were isolated and reverse-transcribed. The resulting cDNAs were subjected to PCR with indicated primers and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies.
Proteins were visualized using an ECL detection system. Actin was used as an internal control.
0
A
10 20 30 40
Genistein ( M)µ
0 10 20 30 40
Genistein ( M)µ
Cdk2
Cdc2
GAPDH
0
B
10 20 30 40
Genistein ( M)µ
0 10 20 30 40
Genistein ( M)µ
AGS SK-MEL-2
Cdk2
Cdc2
Actin
야 한다. 이들 중 cyclin B/Cdc2 compex는 maturation-pro- moting factor 또는 M phase-promoting factor이며 세포분열 에 관여하는 것으로 알려져 있다.36,37) G2/M기로의 진행 에 있어서 cyclin A 및 cyclin B의 합성이 없을 경우 Cdk2 및 Cdc2는 불활성 상태로 존재하며 그 세포는 G2/M기로 진입을 할 수 없게 된다. 이와 같은 세포주기에 관여하는 유전자들의 조절은 암을 치료하는 표적으로 유용하다.32) 상기 결과에서 알 수 있듯이 genistein 처리에 의한 세 포증식 억제효과는 세포주기 G2/M arrest와 연관이 있었 기에, G2/M기에 관여하는 cyclins 및 Cdks의 발현에 미치 는 genistein의 영향을 조사하였다. Fig. 4 및 5에서 나타난 바와 같이 genistein의 농도 증가에 따른 유전자들의 변화 가 관찰되었는데 먼저 AGS 인체위암세포의 경우 mRNA 및 단백질 수준에서 cyclin A 및 cyclin B1의 발현감소가 관찰된 반면 Cdk2 및 Cdc2는 아무런 변화가 관찰되지 않 았다. 하지만 SK-MEL-2 흑색종세포는 cyclins의 변화는 관찰되지 않은 반면에 Cdk2의 발현이 현저하게 감소하 는 것으로 나타났다. 이상의 결과에서 genistein 처리에 따라 G2/M기에서의 세포주기 억제는 AGS 인체위암세포 의 경우 cyclins의 발현감소 및 SK-MEL-2 흑색종세포의 경우 Cdk2의 발현감소가 중요한 요인으로 작용한 것으 로 생각된다.
5. Cdc25C 및 Wee1의 발현에 미치는 genistein의 영향
Cdc2의 활성은 cyclin B와의 결합에 의해서뿐만 아니라 Cdc2 자체의 인산화 정도에 의해서도 영향을 받는다.
Cdc25C와 Wee1은 Cdc2의 Try-15 잔기의 인산화를 조절
하여 결과적으로 Cdc2 kinase activity와 mitosis로의 진입을 조절한다. 먼저 Cdc25C의 경우는 Cdc2의 Try-15 잔기를 탈인산화하여 Cdc2를 활성화시키고 mitosis를 촉진한
다.38,39) 이와 반대로 Wee1의 경우는 세포가 mitosis로 들
어갈 수 있는 조건이 될 때까지 Cdc2의 Try-15 잔기를 인산화하여 그 활성을 억제하는 것으로 알려져 있다.40,41) 즉 late G2에서 Cdc25C의 활성은 증가되고 Wee1의 활성 은 억제되어 Cdc2가 탈인산화된 상태로 활성화되어 세 포주기가 mitosis로 진행하게 된다.
따라서 G2/M기 진행을 조절하는 또 다른 인자인 Cdc 25C 및 Wee1의 발현에 미치는 genistein의 영향을 알아본 결과는 다음과 같다. Fig. 6에서 나타난 바와 같이 Wee1 의 경우는 두 세포주 모두에서 큰 변화를 관찰할 수 없 었지만 Cdc25C의 경우는 genistein의 처리농도가 증가할 수록 발현정도가 감소하는 것으로 나타났다. 즉 AGS 인 체위암세포 및 SK-MEL-2 흑색종세포에서 genistein의 처 리에 따라 Wee1의 발현과는 무관하게 Cdc25C의 발현이 감소한 것은 Cdc2가 활성화될 수 없었기 때문에 mitosis 로의 진행이 억제되어 G2/M기에서 세포주기가 억제된 것으로 추측된다.
6. p53 및 Cdk inhibitor p21의 발현에 미치는 genistein의 영향
세포주기는 Cdk inhibitors (CKIs)의 세포 내 농도에 의 하여 조절된다. CKIs는 CIP/KIP family 및 INK family로 나 누어지는데 이들은 cyclin/Cdk 복합체와 결합하여 세포주 기를 억제한다.35,42) CKIs 중 CIP/KIP family에 속하는 p21 은 종양 억제 유전자인 p53에 의하여 활성화되어 G1, S
Fig. 6. Effects of genistein treatment on the p-Cdk2, Cdc25C, p-Cdc25C and Wee1 expression in AGS human gastric carcinoma and SK-MEL-2 melanoma cells. The cells were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
0 10 20 30 40
Genistein ( M)µ
0 10 20 30 40
Genistein ( M)µ
p-Cd2 (Tyr15)
Cdc25C
P-Cdc25C (Ser216)
Wee1
Actin
AGS SK-MEL-2
및 G2/M기 전체에 걸쳐서 세포주기의 진행을 억제한 다.43) p21의 활성화에는 p53이 관여하는 것이 일반적이 지만 p53 비의존적으로 활성화된다고도 보고되어지고 있다.44∼48)
본 연구에서는 genistein에 의한 G2/M기의 세포주기 억 제에 종양 억제 유전자 p53 및 Cdk inhibitor p21이 어떠한 영향을 미치는지를 조사하였다. Fig. 7의 결과에서 알 수 있듯이 SK-MEL-2 흑색종세포에서는 두 유전자 모두 아 무런 변화가 관찰되지 않았지만 AGS 인체위암세포에서 는 p53 비의존적인 경로로 p21 유전자의 발현이 증가됨 을 알 수 있었다.
결 론
본 연구에서는 genistein이 AGS 인체위암세포 및 SK- MEL-2 흑색종세포의 증식에 미치는 영향에 대해서 조사 하였다. Genistein은 두 세포주 모두에서 처리 농도 의존 적으로 형태적인 변형을 유발하였고 세포의 증식을 억 제하였다. 이러한 형태적인 변형 및 증식억제는 G2/M기 의 세포주기 억제와 연관이 있음을 flow cytometry를 이용 한 세포주기 분석을 통하여 알 수 있었다. 따라서 이러한
G2/M기의 세포주기 억제에 관여하는 유전자들을 RT- PCR 및 western blotting을 이용하여 조사한 결과 두 세포 주 사이의 유전자 발현의 정도가 다르게 나타났다. 먼저 AGS 인체위암세포의 경우는 cyclin A 및 cyclin B1의 발현 저하, p53 비의존적인 p21의 선택적인 발현 증가 및 Cdc25C의 번역 수준에서의 발현저하에 의하여 genistein 에 의한 G2/M기 세포주기 억제가 나타났으며 SK-MEL-2 흑색종세포의 경우는 cyclins, p53 및 p21의 발현변화 없 이 Cdk2의 전사 및 번역 수준에서의 발현저하 및 Cdc25C 의 번역 수준에서의 발현저하에 의하여 G2/M기에서 세 포주기가 억제되는 것으로 추정된다. 이상의 결과들을 살펴볼 때 genistein이 AGS 인체위암세포 및 SK- MEL-2 흑색종세포에서 효과적인 화학요법제로 사용할 수 있을 가능성을 제시할 수 있었으나, 보다 다양한 접근을 통한 항암효과의 기전에 관한 부가적인 조사가 필요할 것으 로 생각한다.
감사의 글
이 논문은 부산대학교 자유과제 학술연구비(2년)에 의 하여 연구되었습니다.
Fig. 7. Induction of Cdk inhibitor p21 by genistein treatment in AGS human gastric carcinoma and SK-MEL-2 melanoma cells. (A) Cells were cultured for 48 h in the absence or in the presence of genistein and total RNAs were isolated and reverse-transcribed.
The resulting cDNAs were subjected to PCR with indicated primers and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) After 48 h incubation with genistein, the cells were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system.
Actin was used as an internal control.
0
A
10 20 30 40
Genistein ( M)µ
0 10 20 30 40
Genistein ( M)µ
p53
p21
GAPDH
0
B
10 20 30 40
Genistein ( M)µ
0 10 20 30 40
Genistein ( M)µ
AGS SK-MEL-2
p53
p21
Actin
참 고 문 헌
1) Pines J. Protein kinases and cell cycle control. Semin Cell Biol 5, 399-408, 1994.
2) Li A, Blow JJ. The origin of CDK regulation. Nat Cell Biol 3, 182-184, 2001.
3) Oren M. Decision making by p53: life, death and cancer. Cell Death Differ 10, 431-442, 2003.
4) Swanton C. Cell-cycle targeted therapies. Lancet Oncol 5, 27-36, 2004.
5) McGowan CH, Russell P. Human Wee1 kinase inhibits cell division by phosphorylating p34cdc2 exclusively on Tyr15.
EMBO J 12, 75-85, 1993.
6) Booher RN, Holman PS, Fattaey A. Human Myt1 is a cell cycle-regulated kinase that inhibits Cdc2 but not Cdk2 activity. J Biol Chem 272, 22300-22306, 1997.
7) Jin P, Gu Y, Morgan DO. Role of inhibitory CDC2 phos- phorylation in radiation-induced G2 arrest in human cells. J Cell Biol 134, 963-970, 1996.
8) Vidal A, Koff A. Cell-cycle inhibitors: three families united by a common cause. Gene 247, 1-15, 2000.
9) Lowe SW, Cepero E, Evan G. Intrinsic tumour suppression.
Nature 432, 307-315, 2004.
10) Messina M, Barnes S. The role of soy products in reducing risk of cancer. J Natl Cancer Inst 83, 541-546, 1991.
11) Knight DC, Eden JA. A review of the clinical effects of phytoestrogens. Obstet Gynecol 87, 897-904, 1996.
12) Mills PK, Beeson WL, Phillips RL, Fraser GE. Cohort study of diet, lifestyle, and prostate cancer in Adventist men. Cancer 64, 598-604, 1989.
13) Adlercreutz CH, Goldin BR, Gorbach SL, Hockerstedt KA, Watanabe S, Hamalainen EK, Markkanen MH, Makela TH, Wahala KT, Adlercreutz T. Soybean phytoestrogen intake and cancer risk. J Nutr 125, 757-770, 1995.
14) Mukhopadhyay D, Tsiokas L, Zhou XM, Foster D, Brugge JS, Sukhatme VP. Hypoxic induction of human vascular endothelial growth factor expression through c-Src activation.
Nature 375, 577-581, 1995.
15) Tatsuta M, Iishi H, Baba M, Yano H, Uehara H, Nakaizumi A. Attenuation by genistein of sodium-chloride-enhanced gastric carcinogenesis induced by N-methyl-N'-nitro-N-nitro- soguanidine in Wistar rats. Int J Cancer 80, 396-399, 1999.
16) Okura A, Arakawa H, Oka H, Yoshinari T, Monden Y. Effect of genistein on topoisomerase activity and on the growth of [Val 12]Ha-ras-transformed NIH 3T3 cells. Biochem Biophys Res Commun 157, 183-189, 1988.
17) Adlercreutz H. Western diet and Western diseases: some hormonal and biochemical mechanisms and associations. Scand J Clin Lab Invest 201, 3-23, 1990.
18) Huang J, Nasr M, Kim Y, Matthews HR. Genistein inhibits protein histidine kinase. J Biol Chem 267, 15511-15515, 1992.
19) Wei H, Bowen R, Cai Q, Barnes S, Wang Y. Antioxidant and antipromotional effects of the soybean isoflavone genistein. Proc Soc Exp Biol Med 208, 124-130, 1995.
20) Barnes S. Related effect of genistein on in vitro and in vivo models of cancer. J Nutr 125, 777-783, 1995.
21) Setchell KD. Phytoestrogens: the biochemistry, physiology, and implications for human health of soy isoflavones. Am J Clin Nutr 68, 1333-1346, 1998.
22) Peterson G, Barnes S. Genistein and biochanin A inhibit the growth of human prostate cancer cells but not epidermal growth factor receptor tyrosine autophosphorylation. Prostate 22, 335-345, 1993.
23) Dave B, Eason RR, Till SR, Geng Y, Velarde MC, Badger TM, Simmen RC. The soy isoflavone genistein promotes apoptosis in mammary epithelial cells by inducing the tumor suppressor PTEN. Carcinogenesis 26, 1793-1803, 2005.
24) Hotz MA, Del Bino G, Lassota P, Traganos F, Darzynkiewicz Z. Cytostatic and cytotoxic effects of fostriecin on human promyelocytic HL-60 and lymphocytic MOLT-4 leukemic cells. Cancer Res 52, 1530-1535, 1992.
25) Monti E, Sinha BK. Antiproliferative effect of genistein and adriamycin against estrogen-dependent and -independent hu- man breast carcinoma cell lines. Anticancer Res 14, 1221- 1226, 1994.
26) Pagliacci MC, Smacchia M, Migliorati G, Grignani F, Riccardi C, Nicoletti I. Growth-inhibitory effects of the natural phyto- oestrogen genistein in MCF-7 human breast cancer cells. Eur J Cancer 30, 1675-1682, 1994.
27) Spinozzi F, Pagliacci MC, Migliorati G, Moraca R, Grignani F, Riccardi C, Nicoletti I. The natural tyrosine kinase inhibitor genistein produces cell cycle arrest and apoptosis in Jurkat T-leukemia cells. Leuk Res 18, 431-439, 1994.
28) Li Y, Bhuiyan M, Sarkar FH. Induction of apoptosis and inhibition of c-erbB-2 in MDA-MB-435 cells by genistein. Int J Oncol 15, 525-533, 1999.
29) Choi YH, Kong KR, Kim YA, Jung KO, Kil JH, Rhee SH, Park KY. Induction of Bax and activation of caspases during beta-sitosterol-mediated apoptosis in human colon cancer cells.
Int J Oncol 23, 1657-1662, 2003.
30) Bartek J, Bartkova J, Lukas J. The retinoblastoma protein pathway in cell cycle control and cancer. Exp Cell Res 237, 1-6, 1997.
31) Sherr CJ. The Pezcoller lecture: cancer cell cycles revisited.
Cancer Res 60, 3689-3695, 2000.
32) Stewart ZA, Westfall MD, Pietenpol JA. Cell-cycle dysregula- tion and anticancer therapy. Trends Pharmacol Sci 24, 139-145, 2003.
33) Marks PA, Richon VM, Rifkind RA. Cell cycle regulatory proteins are targets for induced differentiation of transformed cells: Molecular and clinical studies employing hybrid polar compounds. Int J Hematol 63, 1-17, 1996.
34) Yang J, Kornbluth S. All aboard the cyclin train: subcellular trafficking of cyclins and their CDK partners. Trends Cell Biol
9, 207-210, 1999.
35) Elledge SJ, Harper JW. Cdk inhibitors: on the threshold of checkpoints and development. Curr Opin Cell Biol 6, 847-852, 1994.
36) Jackman MR, Pines JN. Cyclins and the G2/M transition.
Cancer Surv 29, 47-73, 1997.
37) O'Connell MJ, Walworth NC, Carr AM. The G2-phase DNA-damage checkpoint. Trends Cell Biol 10, 296-303, 2000.
38) Strausfeld U, Labbe JC, Fesquet D, Cavadore JC, Picard A, Sadhu K, Russell P, Doree M. Dephosphorylation and activation of a p34cdc2/cyclin B complex in vitro by human CDC25 protein. Nature 351, 242-245, 1991.
39) Hoffmann I, Karsenti E. The role of cdc25 in checkpoints and feedback controls in the eukaryotic cell cycle. J Cell Sci 18, 75-79, 1994.
40) Kornbluth S, Sebastian B, Hunter T, Newport J. Membrane localization of the kinase which phosphorylates p34cdc2 on threonine 14. Mol Biol Cell 5, 273-282, 1994.
41) Fattaey A, Booher RN. Myt1: a Wee1-type kinase that phosphorylates Cdc2 on residue Thr14. Prog Cell Cycle Res 3, 233-240, 1997.
42) Harper JW. Cyclin dependent kinase inhibitors. Cancer Surv
29, 91-107, 1997.
43) Frey MR, Saxon ML, Zhao X, Rollins A, Evans SS, Black JD.
Protein kinase C isozyme-mediated cell cycle arrest involves induction of p21 (waf1/cip1) and p27 (kip1) and hypophos- phorylation of the retinoblastoma protein in intestinal epi- thelial cells. J Biol Chem 272, 9424-9435, 1997.
44) Datto MB, Yu Y, Wang XF. Functional analysis of the transforming growth factor beta responsive elements in the WAF1/Cip1/p21 promoter. J Biol Chem 270, 28623-28628, 1995.
45) Gartel AL, Tyner AL. The role of the cyclin-dependent kinase inhibitor p21 in apoptosis. Mol Cancer Ther 1, 639-649, 2002.
46) Xiong Y, Hannon GJ, Zhang H, Casso D, Kobayashi R, Beach D. p21 is a universal inhibitor of cyclin kinases. Nature 366, 701-704, 1993.
47) Zeng YX, el-Deiry WS. Regulation of p21WAF1/CIP1 ex- pression by p53-independent pathways. Oncogene 12, 1557- 1564, 1996.
48) Choi YH, Lee WH, Park KY, Zhang L. p53-independent induction of p21 (WAF1/CIP1), reduction of cyclin B1 and G2/M arrest by the isoflavone genistein in human prostate carcinoma cells. Jpn J Cancer Res 91, 164-173, 2000.