솔장다리 추출물의 항산화 활성 및 세포주기조절에 의한 항암 활성 분석
오유나1, 진수정1, 박현진1, 권현주1,2, 김병우1,2*
1동의대학교블루바이오소재개발센터
2동의대학교자연과학대학생명응용학과
Received: November 27, 2013 / Revised: March 6, 2014 / Accepted: March 11, 2014
서 론
우리나라통계청에서발표한 2012년사망원인통계자료 에의하면 3대사망원인은암, 심장질환, 뇌혈관질환으로 총사망자의 47.1%를차지하고, 그중암은 30.4%로가장높 은사망률을나타낸다고보고하였다. 특히대장암의경우, 그
사망률이전년대비 5.7% 증가하였으며여성암환자의경우
폐암에이어두번째로높은발병률을보이며해마다그발 생률이점차증가하고있다. 또한전세계적으로도사망원인
의네번째를차지하고있다[40]. 대장암의발병원인은정확
히알려져있지는않으나흡연, 서구화된식습관, 비만, 스트
레스, 유전적요인등의관련성이보고되고있다[3, 8, 20].
암환자의약 70~80%가항암제치료중오심, 구토등부
작용을경험하며, 이러한부작용증상에대하여 1990년대말
부터 NK1 receptor antagonist인 aprepitant나 2세대 sertron계인 palonsetron 등을사용중이지만항우울제로분 류되는 aprepitant 성분은또다른부작용을낳을우려가보
고되고있다[7, 19]. 또한현재항암제로쓰여지고있는여러
물질들은강한독성을가져, 암세포뿐만아니라정상세포까 지영향을미치는등부작용이많은실정이다. 따라서암의 치료뿐아니라예방을위해가능한독성이적고효과적인항 암제의개발이절실히필요한현실이다. 따라서최근몇십
년간 새로운 항암제 개발을 위한 분자 마커(molecular
marker)가빠르게개발되고있으며다양한 growth factor, 신호전달관련인자, 세포사멸유도및세포주기조절관련
인자들을타겟으로하여많은연구가진행되고있다[14, 15,
Anti-oxidative and Anti-cancer Activities by Cell Cycle Regulation of Salsola collina Extract You Na Oh1, Soojung Jin1, Hyun-jin Park1, Hyun Ju Kwon1,2, and Byung Woo Kim1,2*
1Blue-Bio Industry Regional Innovation Center, Dong-Eui University, Busan 614-714, Republic of Korea
2Department of Life Science and Biotechnology, College of Natural Sciences, Dong-Eui University, Busan 614-714, Republic of Korea
Salsola collina, also known as Russian thistle, is widely distributed in and around waste facilities, roadsides, and drought and semi-drought areas, and is used as a traditional folk remedy in Chinese medicine for the treatment of hypertension. In this study, we have evaluated the anti-oxidative and anti-cancer activities of the ethanol extract of S. collina Pall. (EESC), and the molecular mechanisms of its anti-cancer effects on human colon carcinoma HT29 cells. EESC exhibited anti-oxidative activity through DPPH radical scavenging capacity and showed cytotoxic activity in a dose-dependent manner in HT29 cells. After EESC treatment, HT29 cells altered their morphology, becoming smaller and irregular in shape. EESC also induced cell accu- mulation in the G2/M phase in a dose-dependent manner, accompanied by a decrease of cell population in the G1 phase. The G2/M arrest by EESC was associated with the increased expression of cyclin-dependent kinase (CDK) inhibitor p21 and Wee1 kinase, which phosphorylates, or inactivates, Cdc2. EESC treatment induced the phosphorylation of Cdc2 and Cdc25C, and inhibited cyclin A and Cdc25C protein expression. In addition, S arrest was induced by the highest concentration of EESC treat- ment, associated with a decrease of cyclin A and Cdk2 expression. These findings suggest that EESC may possess remark- able anti-oxidative activity and exert an anti-cancer effect in HT29 cells by cell cycle regulation.
Keywords: Anti-cancer activity, anti-oxidative activity, Cdc25C, G2/M arrest, p21, Salsola collina
*Corresponding author
Tel: +82-51-890-2900, Fax: +82-51-890-2914 E-mail: [email protected]
© 2014, The Korean Society for Microbiology and Biotechnology
26]. 또한부작용및독성이적은항암제의개발을위하여천 연물, 특히식물유래천연물로부터항암성분을찾는연구가 활발히진행되고있으며, 다양한성분들이혼합되어있는천 연물의특징을이용하여천연물의조추출물또는용매분획 물을사용하는의약품개발및단일화합물이나합성된유
도체등을통한새로운의약품개발이시도되고있다[37]. 또
한이들천연물을이용한의약품개발은천연물내존재하 는다양한성분들의상호작용을통하여다수의타겟에작용 할수있어, 다양한질환에대한조합치료가가능하도록하 는장점을가진다.
정상세포는두개의세포로분열되는과정에서 G1, S, G2,
M (mitosis) 주기를가진다. G1기는세포를분열하기위한 준비단계로 checkpoint가존재하여분열에필요한인자들이 부족하거나 DNA에 damage가있을때 G1기에머무르게하 여다음단계인 S기로이행하지못하게한다. DNA 복제가이 루어지는 S기를지나면두번째 check point가존재하는 G2 기에의해 M기로의이행을조절하게되며 M기로의진행이 이어지면증폭된염색체는전기(Prophase), 중기(Metaphase), 후기(Anaphase) 및말기(Telophase)를거쳐세포분열을완 성한다[36]. 이러한세포주기는각 checkpoint 단계에서적 절히조절되지않으면세포분열을적절하게제어하지못하 고이상증식을야기하여암의원인이되기도한다[29, 32].
실제로암세포는정상세포와달리세포주기제어를벗어나 세포자멸사기전을회피하고, 무한대로분열하는능력을가 지며, 따라서암세포의세포주기를조절함으로써강력한항 암효과를가질수있고이러한세포주기제어에의한항암
활성에관하여활발하게연구되고있다[16].
세포주기의 조절에 관여하는 checkpoint에서는 Cyclin- dependent kinase (CDKs)와 cyclin complex의활성화에의
해 다음 단계로의 진행이 일어나며 이러한 CDK/cyclin
complex의활성을억제하며세포주기를조절한다[18, 35].
특히 G2/M checkpoint인 Cdc2 (CDK1)는 cyclinB1과 complex 를이루어 mitosis 단계로의진행을유도하며특정 kinase와 phosphatase에의한인산화/탈인산화작용에의해활성이조 절된다[22]. Wee1 kinase에의해 Cdc2의 Tyr-15 잔기가인 산화되어 Cdc2의 불활성화가 일어나며, 이러한 inactive form Cdc2는 Cdc25C phosphatase에의해탈인산화되면서 활성화형태로전환된다[6, 10, 33]. 또한 CDK 억제제 p21 (WAF1/CIP1)은종양억제유전자인 p53의발현증가에의해 유도되며, p21에의해 CDK/cyclin complex의활성이억제된 다[13, 31].
솔장다리(Salsola collina Pall.)는중국북동부, 남서부지 역에널리분포되어있으며중국에서는고혈압, 두통, 현기 증치료를위하여음식혹은민간요법으로사용되어지고있 으며, 현재까지는솔장다리추출물의성분규명에관한연구
만되어있을뿐, 솔장다리의생리활성에관하여는보고되어
진바가없다[23]. 따라서본연구에서는솔장다리에탄올추
출물을사용하여솔장다리의항산화능및인체대장암세포주
인 HT29에대한항암활성에관하여분석하였다.
재료 및 방법
시료 준비
솔장다리 에탄올 추출물(Ethanol extracts of Salsola collina Pall., EESC)은한국생명공학연구원, 해외생물소재 허브센터에서구입(분양번호; FBM123-061)하여사용하였으 며, ethyl alcohol 95.0% GR급을사용하여 45oC에서 sonicator 를이용하여 3일동안반복하여추출하고농축후동결건조 시킨 시료로서 적정 농도로 dimethyl sulfoxide (DMSO, Sigma, St. Louis, MO, USA)에용해하였으며−20oC에서보 관하여사용하였다.
DPPH radical 소거 활성 측정을 통한 항산화능 분석
솔장다리추출물의항산화능분석을위하여 1,1-diphenyl-
2-picrylhydrazyl (DPPH) radical 소거법을이용한전자공여 능측정을수행하였다[25]. EESC를농도별(2.56-320 µg/ml) 로메탄올에녹여준비한후 96 well plate에메탄올에용해 된 1.5×10−4 M DPPH 40 µl와각시료 160 µl를분주한혼 합액을 실온에서 30분간 반응시킨후, microplate reader (Molecular Devices, Sunnyvale, CA, USA)를 이용하여
520 nm에서흡광도를측정하였다. 시료를첨가하지않은음
성대조군과비교하여 free radical 소거활성을백분율로나 타내고, 50% 저해농도(IC50)를계산하였다. 양성대조군으 로는대표적인항산화제인 ascorbic acid를사용하였으며측 정값은 3회반복실험의평균값으로나타내었다. 억제능의 백분율공식은다음과같다.
DPPH radical scavenging activity (%) = {1−(A−B)/C}×100
A: sample absorbance 520 nm
B: color control absorbance 520 nm (without DPPH) C: negative control absorbance 520 nm (without sample) 세포배양
본연구에사용한인체대장암세포주 HT29, 폐암세포주
A549 및 간암 세포주 HepG2는 American Type Culture Collection (ATCC, Manassas, VA, USA)로부터 구입하여 사용하였으며, 10% fetal bovine serum (FBS) 및 1% penicillin/
streptomycin이포함된 Dulbecco’s modified Eagle’s medium (DMEM)을사용하여 37oC, 5% CO2조건하에서배양하였다.
암세포 사멸 효과분석
수용성의 tetrazolium salt (WST, water soluble tetrazolium salt)를이용한 dehydrogenase assay 이용하여 EESC에대 한세포독성을측정하였다. 3-5×104 cell의 HT29, A549 및 HepG2 세포를 24-well plate에분주하여 24시간배양한다 음, EESC를농도별로처리하고 37oC에서 48시간동안배양 하였다. 이때대조군은 0.1% DMSO를처리하였다. 48시간 배양 후 WST assay reagent (Daeillab, Seoul, Korea)를 well당 500 µl 첨가하여 37oC에서 30분동안 반응시킨후, microplate reader로 450 nm에서흡광도를측정하였다. 측 정값은 3회반복실험을하여그에대한평균값으로나타내 었으며, 본실험결과를바탕으로이후적정처리농도를결 정하였다.
세포 형태 변화 관찰
EESC 처리에따른 HT29 세포의형태변화관찰을위하
여 6-well plate에 1×105 cell의 HT29 세포를분주하여 24시 간동안배양후, 적정농도의 EESC를처리하여 48시간동 안배양하였다. 이후 HT29 세포의형태변화를위상차현미 경을 이용하여 200배의 배율로 관찰한 다음 Axio Vision
program을사용하여사진촬영을하였다.
세포주기변화 분석
HT29 세포에서 EESC의세포주기에미치는영향을알아
보기위하여, HT29 세포에 EESC를농도별로처리한다음 CycleTEST™ PLUS DNA Reagent Kit (Becton Dickinson,
San Jose, CA, USA)를사용하여세포주기변화를분석하였
다. 먼저 6-well plate에 HT29 세포를 5×105 cell의농도로
분주하여부착시킨후, EESC를다양한농도로처리하였다.
24시간 배양 후 세포를 회수하고 PBS로 세척한 다음,
ribonuclease A를 실온에서 10분간 처리하고 propidium iodide (PI) 용액을첨가하여 4oC에서 10분간염색하였다. 염 색된세포는 Flow cytometry (Cell Lab Quanta SC, Beckman Coulter, Fullerton, CA, USA)를이용하여분석하였다.
DAPI 염색에 의한 핵의 형태변화 관찰
EESC를처리한 HT29 세포를회수하여 cytospin (Thermo scientific, Waltham, MA, USA)을사용하여슬라이드위에 부착시키고 4% formaldehyde 용액으로상온에서 10분간고 정한다음, PBS로세척후 0.5% triton X-100으로상온에서 10분간 permeabilization시켰다. PBS로 3회 세척한 다음 1µg/ml의 4', 6-diamidino-2-phenylindole (DAPI; Sigma, St. Louis, MO, USA) 용액을이용하여염색하였다. 차광한 상태로상온에서 20분간염색한다음, PBS로세척후 Anti- fade mounting 용액을사용하여 mounting하고, 형광현미경
(Carl Zeiss, Jena, Germany)을이용하여 400배배율로핵 의형태변화를관찰하고 Axio Vision Program을이용하여 촬영하였다.
세포주기 관련 단백질 발현 분석
EESC의항암활성기전을밝히기위해세포주기중 G2/
M기 조절에 관여하는 단백질 발현을 Western blot
hybridization으로 분석하였다. 농도별로 EESC를 처리한 HT29 세포를회수하여 PBS로세척한후, lysis buffer [20 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Triton X-100, 1 µg/ml leupeptin, 1 mM PMSF]를 첨가하여 4oC에서 20분간반응시킨다음 15,000 ×g로 20분 간원심분리하여상층액을취하였다. Bradford assay로추 출한 단백질의 농도를 결정한 후 30 µg의 단백질을 10%
SDS-PAGE로 전기영동하고 nitrocellulose membrane에 blotting한후대상단백질의항체와 hybridization하였다. Membrane washing 후 Horse radish peroxidase (HRP)가 tagging된이차항체로한시간동안반응시킨후 Western blotting luminol reagent (Santa Cruz Biotechnology, Dallas, TX, USA)를 적용시킨 다음 chemiluminescence detection system (FluoChem® FC2, AlphaInnotech, San Leandro, CA, USA)을사용하여분석하였다. p53, Cyclin A, Cyclin B1, Cdc2, phospho-Cdc2 (Tyr15), Cdc25C, Wee1 및 actin의일차항체와 HRP-conjugated anti-rabbit, anti- goat, anti-mouse 등 이차항체는 Santa Cruz Biotechnology (Dallas, TX, USA)에서구입하였으며, 대상단백질인 p21, phospho-Cdc25C의일차항체는 Cell Signaling Technology (Danvers, MA, USA)에서구입하여사용하였다.
통계분석
모든결과는 mean±SD (Standard deviation)로나타내 었으며, 분석된실험데이터의통계적유의성은대조군과각 시료처리군의실험데이터로부터 Student’s t test를통하여 검증하였다.
결과 및 고찰
EESC의 항산화능 분석
활성산소는노화, 암, 관절염등에직간접적으로생체장 애를일으키는원인으로잘알려져있으며따라서항암활성 을포함한다양한생리활성후보소재의항산화능의보유유
무의확인은매우중요하다[39]. 이러한항산화작용의지표
로전자공여능분석이널리사용되고있으며, 전자공여작용 은인체내에서생성되는프리라디칼의전자를공여하여프 리라디칼에의한노화와질병을억제하는작용으로, 식물추
출물의항산화능측정에많이사용되고있다. 특히 DPPH는 비교적안정한 free radical로써 ascorbic acid, tocopherol, polyhydroxy 방향족화합물, 방향족아민류등에의해환원 되어짙은자색이탈색이되며, 이러한원리를이용하여식 물이나식품추출물또는단일화합물의항산화능측정법으 로 이용된다[42]. EESC의 항산화능을 DPPH radical scavenging activity 측정을통해분석한결과, Table 1에제
시한 바와 같이 EESC의 농도가 증가함에 따라 DPPH
radical 억제능이증가하여 2.56 µg/ml에서는 DPPH radical 억제능이 39.4%, 12.8 µg/ml에서는 87.84% 그리고 64 µg/
ml에서는 95%까지억제시키는것을확인하였다. 또한억제 능 50%일때의 EESC의농도(IC50)는 4.82 µg/ml로양성대 조군인 ascorbic acid의 IC50인 1.31 µg/ml보다는다소높았 으나, 추출물인것을감안하면매우강력한항산화활성을 지니는것을알수있었다.
EESC의 암세포 사멸 효과
EESC가강력한항산화활성을지니는것을확인하였으므 로다음으로암세포에대한사멸효과를보이는지알아보았
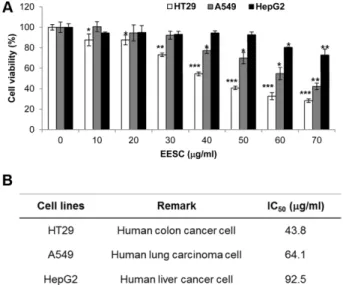
다. EESC의다양한암세포의생존율에미치는영향을알아
보기위하여인체대장암세포 HT29, 폐암세포 A549 및간 암세포 HepG2에 EESC를처리하여 WST assay를사용하여 세포사멸효과를확인하였다. 그결과 Fig. 1A에제시한바 와같이암세포주의종류에따라암세포사멸효과의정도가 다른것을확인할수있었다. HT29 세포의경우, 30 µg/ml 의 EESC 농도에서부터세포증식이억제되었으며, 43.8 µg/
ml의 EESC 농도에서는 50% 사멸효과(IC50)를보여세종류 의암세포주중가장강력한세포사멸효과를보였다. 또한 A549 세포의경우 40 µg/ml의 EESC처리에의해사멸효과 를보이기 시작했으며 64.1µg/ml의 IC50를보였다. 반면, HepG2의경우는 60 µg/ml의 EESC 농도에서부터세포증식 억제를보였으며 EESC 처리최고농도인 70 µg/ml에서도 70% 정도의세포사멸효과를보여세종류의암세포중가장
효과가약한것을알수있었다. 각암세포에대한세포사멸 효과의 IC50는 Fig. 1B에나타내었다. 이상의결과를바탕으
로이후실험은인체대장암세포주 HT29를사용하여진행
하였다.
HT29의 세포형태에 미치는 EESC의 영향
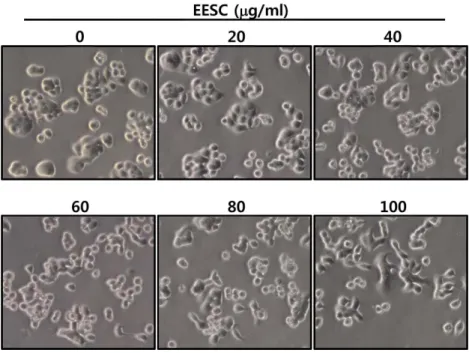
다음으로 EESC 처리가 HT29 세포의형태변화를유도하
는지관찰하기위하여 HT29 세포에 EESC를농도별로처
리한후세포의형태변화를위상차현미경으로관찰하였다. 그결과, Fig. 2에서와같이 EESC의처리농도에따라세포 밀도가점차감소하여세포증식이억제됨을알수있었다. 또한 40 µg/ml의 EESC를처리하였을때부터점차적으로세 포형태가불규칙적으로변하고, 무리를이루어자라는형태 가단일세포형태로변하는것을관찰할수있었다. 이러한 결과로부터 EESC의처리에의해 HT29의세포형태에변 화가일어남을확인하였다.
EESC에 의한 세포주기 변화 분석
현재까지다양한암세포에서정상세포와는달리 G1기및 G2기의 checkpoint에의한정상적인세포주기조절에서벗 어나비정상적으로무한증식이일어난다는것이보고되어
있다[17]. 또한최근에는이러한암세포의세포주기제어에서
Table 1. DPPH radical scavenging activity by ethanol extract of S. collina Pall.
Concentration (µg/ml)
Inhibition rate (%)
IC50 (µg/ml) EESC
2.56 39.40± 2.85 12.8 87.84± 0.92 4.82 64 95.07± 0.54 320 95.80± 0.23 Ascorbic acid
0.51 24.16± 1.98
1.31 2.56 89.83± 2.98
12.8 97.04± 1.49
Fig. 1. Effects of EESC on cell growth in various cancer cell lines. (A) Human colon carcinoma HT29 cells, human lung carci- noma A549 cells, and human hepatocellular carcinoma HepG2 cells were treated with various concentrations of EESC for 48 h.
Cytotoxic effect of EESC was determined by WST assay. Results are expressed as percentage of the vehicle treated control± SD of three independent experiments. *, p < 0.05, **, p < 0.005 and ***, p < 0.001 compared with DMSO treated cells. (B) IC50 values of human cancer cells are shown.
Fig. 2. Morphological changes by EESC in HT29 cells. Cells were treated with indicated concentration of EESC for 48 h, and then visualized by light microscopy. Magnification, ×200
Fig. 3. Induction of G2/M arrest in EESC-treated HT29 cells. HT29 cells were cultured and treated with EESC for 24 h. Cells were harvested, and then stained with propidium iodide for 30 min and analyzed by flow cytometry. DNA-flourescence histogram (A) and quan- titative data of cell distribution (B) are shown. M1, SubG1 phase; M2, G1 phase; M3, S phase; M4, G2/M phase.
벗어나는분자적기전에관하여활발히연구되고있으며, 새 로운항암제연구를위하여암세포의세포주기를제어하는
신물질을찾는노력들이활발히진행되고있다[37]. 따라서
본연구에서는 EESC에의한 HT29 세포의증식억제현상 이세포주기조절에의한것인지살펴보았다. HT29 세포에 EESC를농도별로처리한후 PI 염색을하여세포주기의변 화를 Flow cytometry로분석하였다. 그결과, Fig. 3A에서
보여지는바와같이, EESC 농도의존적으로 G2/M기의세포
비율이증가되는것을확인하였으며, 최고농도인 60 µg/ml 의 EESC를처리하였을때 G2/M기뿐아니라 S기의세포비 율도증가되는것을알수있었다. 또한 G2/M arrest가유도 되면서상대적으로 G1기의세포분포는점차감소됨을확인
하였다. 이러한 EESC 처리에따른세포주기별세포비율은
정량화하여 Fig. 3B에나타내었다. 대조군에서 16.86%였던 G2/M기 세포분포는 EESC 최고 농도인 60 µg/ml에서는 24.9%의세포가 G2/M기에정체되어있음을확인하였다. 또 한대조군에서 26.15%였던 S기의세포분포는최고농도인 60 µg/ml에서는 34.23%로증가되었다. 반면, G1기의세포
분포는 51.87%에서 36.93%로감소하였다. 따라서이와같 은결과는 EESC에의해 HT29 세포의 G2/M arrest가유도 됨을시사하며최고농도에서는 S기및 G2/M기에서세포주
기가정지되는것으로사료된다[4, 45].
다음으로 EESC에의한세포주기정지에따른핵내의변
화를관찰하기위하여 HT29 세포에 EESC를농도별로처
리한다음 DAPI 염색을수행하였다. 그결과, Fig. 4에서와
같이 EESC 처리농도가증가할수록핵내농축된염색체
가 관찰되었으며, 특히 M (mitosis)기의 단계 중 전중기 (Prometaphase, Arrow in Fig. 4), 및 후기(Anaphase, Arrowhead in Fig. 4) 단계에있는세포들이증가하는것을
알수있었다. 따라서이러한결과들은 EESC 처리에의해
G2/M arrest가일어나 mitosis가완전히일어나기전단계
의세포가많이관찰되는것을의미한다[9].
EESC가 세포주기 관련 단백질 발현에 미치는 영향 세포주기를조절하는 checkpoint는 G1/S기및 G2/M기에 존재하며, 이때 CDK/cyclin complex가 checkpoint의중요 한인자로작용하게되고, 이러한 CDK/cyclin complex의활 성여부에따라세포증식이촉진되고억제되기도한다[1]. G1/
S기 checkpoint 중에서 S기에서세포주기진행을조절하는 인자는 cyclin A/CDK2 complex이며, G2/M기에서는 Cdc2 (CDK1)와 cyclin A 및 cyclin B가중요한인자로작용한다 [28, 34, 41]. 특히 G2 후기와 초기 M기에서는 활성화된 cyclin A/Cdc2 complex에의해 M기로의진행이촉진되며, M기에서는 cyclin B/Cdc2 complex에의해조절된다[5, 27].
따라서 EESC에의한 HT29의세포주기조절관련단백질
의발현에미치는영향을확인하기위하여, S기및 G2/M기
checkpoint 관련 인자들의항체를 사용하여 Western blot analysis를수행하였다. 그결과, EESC 농도의존적으로 G2/
M checkpoint 관련단백질인 Cdc2의인산화가증가되었으며, cyclin A의발현양이감소하였다(Fig. 5A). 그에반해 cyclin B1의발현은증가되었으나, inactive Cdc2와 complex를이 루어 M기의진행을저해할것으로사료된다[24, 43]. 또한 CDK inhibitor인 p21의발현도 EESC 농도의존적으로증가 하였다(Fig. 5B). phospho-Cdc2는 Cdc2의불성활성화형태 로 Cdc2의 ATP binding site에인산기가결합하여 cyclin A/
Cdc2 complex의활성을억제하고, p21은 cyclin/CDK complex
에결합하여활성을억제하는단백질로알려져있다[44]. 따
라서 이러한 결과들로부터 EESC를 처리에 의한 G2/M arrest는 Cdc2의불활성화에의한 G2/M checkpoint인 cyclin A/Cdc2 complex의활성화저해에의한것이라사료된다. 반
면, p21의 발현을 유도하는것으로 알려진 p53의 발현은
EESC처리에의해 아무런변화가관찰되지않았으며(Fig.
5B), 이는 EESC에의한 p21의발현유도는 p53-independent 한 Fig. 4. Conformational changes in nuclear chromatin of HT29
cells by EESC treatment. EESC-treated cells were fixed with 4%
formaldehyde, permeabilized with 0.5% Triton X-100 and stained with DAPI for 20 min. Stained cells were observed by fluores- cence microscopic analysis and imaged using Axio Vision Pro- gram. EESC-treated HT29 cells showed the dense nuclear chromatin in the prometaphase (Arrow) and in the anaphase (Arrowhead) of mitosis. Scale bar, 20 µm.
것임을시사한다[21, 46]. 또한 S기 checkpoint 관련단백질 인 Cdk2의발현또한감소하였으므로(Fig. 5A), 이전의세 포주기분석결과와같이 S기의세포분포가증가하는것은 cyclin A/Cdk2 complex의감소에의해 S기에서 G2기로의 진행이억제되어나타난결과일것으로사료되나자세한기 전은추후연구가필요할것이다[4, 45].
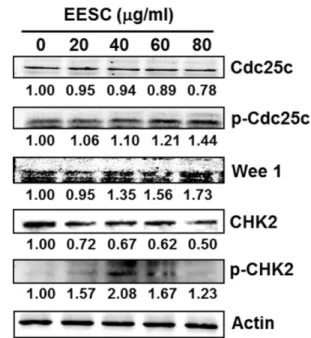
EESC의 G2/M arrest 유도 기전 분석
G2/M checkpoint의중요한단백질인 Cdc2는인산화/탈인 산화에의해그활성이조절되는데, Wee1 인산화효소에의
해인산화되어불활성화된 Cdc2는 Cdc25C 탈인산화효소에 의해 탈인산화되면서 활성화 형태가 된다[10, 33]. 또한 Cdc25C는 DNA damage에의해활성화되는 CHK1/CHK2 인산화효소에 의해 인산화될 때 불활성화된다. 이러한 Cdc25C는 Cdc25 family 중하나로, 식도암, 직장암, 유방암 등다양한 종류의 암세포에서 과발현이관찰되며 이러한
Cdc25C의발현억제는 M기로의진행을저해시켜암세포의
성장을억제한다는보고가있다[2, 30]. 앞서 EESC 처리에
의해세포주기관련단백질인 Cdc2의인산화가증가되는것
을확인하였으므로, EESC의정확한 G2/M arrest 유도기전
을분석하기위하여 Cdc2의활성을조절하는분자들의단백
질발현양상을확인하였다. Fig. 6에서와같이 Cdc2의인산 화를일으키는 Wee1 kinase의발현이증가되고, Cdc25C의 발현은감소하였다. 또한 Cdc25C의불활성화를일으키는인 산화는증가되었으며, Cdc25C의인산화를유도하는 CHK2 의활성화형태인 phospho-CHK2 발현도증가되었다(Fig.
6). 이러한결과들은 EESC 처리에의해 CHK2가활성화되 고, 활성화된 CHK2가 Cdc25C를인산화시켜불활성화시키
고, 따라서 Cdc2의탈인산화에의한활성화가억제되는것
을시사한다. 반면 EESC 처리에의해증가된 Wee1 kinase 는 Cdc2를인산화시켜 Cdc2를불활성화시키고, 최종적으 로 G2기에서 M기로의세포주기진행을저해하는것이라사 Fig. 5. Effects of EESC on the expression of cell cycle-related
proteins in HT29 cells. HT29 cells were treated with various con- centrations of EESC for 24 h. The cells were lysed and cellular proteins were then separated by SDS-PAGE, followed by Western blot analysis using antibodies against S-G2/M transition-related proteins (A) and p53 and p21 (B). Actin was used as an internal control. Numerical bottom panel of each band indicates the fold change in the band intensity compared with that of control.
Fig. 6. Effects of EESC on the expression of upstream signal proteins leading to G2/M arrest in HT29 cells. EESC-treated cells were harvested and lysed, and the proteins were subjected to Western blot analysis using antibodies against Cdc25C, p- Cdc25C, Wee1, CHK2, and p-CHK2. Actin was used as an inter- nal control. Numerical bottom panel of each band indicates the fold change in the band intensity compared with that of control.
료된다[11, 12, 38].
현재전세계적으로증가하고있는암의발병률및사망률 에비해효과적이고부작용이적은암치료제의개발은아 직도부족한실정이며, 따라서기존의항암제에비해효과적 이며암세포에대한특이성이높은새로운작용기전의항암 제개발의요구가대두되고있다. 특히세포주기제어가비 정상적으로작동되어무한증식능력을가지는암의특성을 고려하여, 세포주기조절인자를타겟으로하여세포주기조 절인자들의활성을제어함으로써암세포화를막는치료법 에관해많은관심이집중되고있으며, 그를위한다양한기 전의항암활성을가지는소재에관한연구개발이활발하게 진행중이다. 따라서본연구는솔장다리추출물의뛰어난 항산화및항암활성을확인하고대장암세포의세포주기조 절에의한항암기전을해석하여, 새로운기전의항암활성보 유소재의개발을위한중요한기초자료가될것이라사료 된다. 또한본연구결과를바탕으로하여나아가동물실험 등을통한실질적인솔장다리추출물의항암활성에관한연 구도지속되어야할것이다.
요 약
본 연구에서는 명아주과 수송나물속 솔장다리(Salsola collina Pall.) 추출물의항산화및항암활성을분석하였다. 먼저솔장다리의에탄올추출물의 DPPH radical scavenging activity를분석한결과, IC50가 4.82 µg/ml로나타나강한항 산화능을 보유하였음을 확인하였다. 또한 대장암 세포주 (HT29), 폐암세포주(A549), 간암세포주(HepG2)를사용하 여솔장다리추출물의암세포사멸효과를분석한결과, IC50
가각각 43.8, 64.1, 92.5 µg/ml로강력한세포사멸효과를나
타냈으며특히 HT29에대한강한사멸효과를보였다. 솔장
다리추출물의항암활성기전분석을위해세포주기를분 석한결과, 대장암세포인 HT29의 G2/M arrest를유도하였 으며최고농도인 60 µg/ml까지 S기세포수가증가하였다. 세포주기관련단백질의발현분석결과, 솔장다리추출물을 처리한경우, G2기에서 M기로의전이에필수적인단백질인 Cdc25C와 cyclin A의발현이감소되었고, 반면 Cdc25C와 Cdc2의불활성화형태인 p-Cdc25C, p-Cdc2는증가하였다. 또한 p21과 Wee1의발현도증가되었다. 하지만 p53의발현 량은변화가없었다. 이러한결과는솔장다리추출물을처리 한경우, p53 비의존적으로 p21의발현이증가되어 cyclin A/
Cdc2 complex의활성이 조절되고, 이어서 G2/M phase의 check point에작용하는 Wee1의발현증가및 Cdc25C, Cdc2 의인산화에의한불활성화를통하여 G2/M arrest가유도되 는것을시사한다. 또한솔장다리추출물처리에의해 S기진 행을조절하는 Cdk2의발현량도감소하여, cyclin A/Cdk2
complex가감소되어 S기의세포수가증가한것으로보인다. 따라서본연구결과를통해솔장다리추출물이높은항산 화활성을지니며암세포의세포주기를조절하여높은항암 활성을보유함을확인하였다.
Acknowledgements
This work was supported by Blue-Bio Industry Regional Innova- tion Center (RIC08-06-07) at Dong-Eui University as a RIC pro- gram under Ministry of Trade, Industry & Energy and Busan city.
References
1. Aktas H, Cai H, Cooper GM. 1997. Ras links growth factor signaling to the cell cycle machinery via regulation of cyclin D1 and the Cdk inhibitor p27KIP1. Mol. Cell. Biol. 17: 3850-3857.
2. Albert H, Santos S, Battaglia E, Brito M, Monteiro C, Bagrel D.
2011. Differential expression of CDC25 phosphatases splice variants in human breast cancer cells. Clin. Chem. Lab. Med.
49: 1707-1714.
3. An Y. 1999. Prospective and retrospective studies on risk fac- tors of four most common cancers in Korean males: stomach, live, lung and colorectum. Seoul, Ministry of Health & Welfare.
4. Aranha O, Wood DP Jr, Sarkar FH. 2000. Ciprofloxacin medi- ated cell growth inhibition, S/G2-M cell cycle arrest, and apop- tosis in a human transitional cell carcinoma of the bladder cell line. Clin. Cancer Res. 6: 891-900.
5. Arellano M, Moreno S. 1997. Regulation of CDK/cyclin com- plexes during the cell cycle. Int. J. Biochem. Cell Biol. 29: 559- 573.
6. Booher RN, Holman PS, Fattaey A. 1997. Human Myt1 is a cell cycle regulated kinase that inhibits Cdc2 but not Cdk2 activity. J. Biol. Chem. 272: 22300-22306.
7. Byar KL, Berger AM, Bakken SL, Cetak MA. 2006. Impact of adjuvant breast cancer chemotherapy on fatigue, other symp- toms, and quality of life. Oncol. Nurs. Forum 33: E18-26.
8. Cha MR, Yoon MY, Kim JY, Hwang JH, Park HR. 2006. The cytotoxic effect of the Gleditsiae semen extracts oh human colon carcinoma cells. J. Korean Soc. Appl. Biol. Chem. 49:
248-253.
9. Chang CH, Yu FY, Wu TS, Wang LT, Liu BH. 2011. Mycotoxin citrinin induced cell cycle G2/M arrest and numerical chromo- somal aberration associated with disruption of microtubule for- mation in human cells. Toxicol. Sci. 119: 84-92.
10. Donzelli M, Draetta GF. 2003. Regulating mammalian check- points through Cdc25 inactivation. EMBO Reports 4: 671-677.
11. Dorai T, Aggarwal BB. 2004. Role of chemopreventive agents in cancer therapy. Cancer Lett. 215: 120-140.
12. Dulic V, Stein GH, Far DF, Reed SI. 1998. Nuclear accumula- tion of p21Cip1 at the onset of mitosis: a role at the G2/M-phase transition. Mol. Cell. Biol. 18: 546-557.
13. Elledge SJ, Harper JW. 1994. Cdk inhibitors: on the threshold