274 책임저자:최영현, 614-052, 부산시 진구 양정2동
동의대학교 한의과대학 생화학교실 Tel: 051-850-7413, Fax: 051-853-4306 E-mail: [email protected]
접수일:2009년 8월 13일, 게재승인일:2009년 8월 27일
Correspondence to:Yung Hyun Choi
Department of Biochemistry, Dongeui University College of Oriental Medicine, Yangjung-2dong, Busanjin-gu, Busan 614-052, Korea Tel: +82-51-850-7413, Fax: +82-51-853-4306
E-mail: [email protected]
불등가사리 메탄올 추출물에 의한 U937 인체백혈병세포의 Apoptosis 유발
동의대학교 1블루바이오 소재개발센터, 2자연과학대학 생명응용학과, 3대학원 바이오물질제어학과,
4생활과학대학 식품영양학과, 5한의과대학 생화학교실
박 철1ㆍ현숙경1ㆍ이은우1,2ㆍ권현주1,2,3ㆍ황혜진1,4ㆍ김병우1,2,3ㆍ최영현1,3,5
Apoptosis Induction of Human Leukemia U937 Cells by Methanol Extract of Gloiopeltis furcata
Cheol Park1, Sook Kyung Hyun1, Eun-Woo Lee1,2, Hyun Ju Kwon1,2,3, Hye Jin Hwang1,4, Byung-Woo Kim1,2,3 and Yung Hyun Choi1,3,5
1Blue-Bio Industry RIC, 2Department of Life Science and Biotechnology, College of Natural Sciences, 3Department of Biomaterial Control (BK21 program), Graduate School, 4Department of Food and Nutrition, College of Human Ecology,
5Department of Biochemistry, College of Oriental Medicine, Dongeui University, Busan 614-052, Korea
Gloiopeltis furcata is a kind of edible seaweed that mainly grows in the temperate seaside areas of the northwest pacific including Korea, Japan and China, and has been known to exert pharmacological activities including antioxidant, antimutagenic and anticancer activity. However, the molecular mechanisms of G. furcata in malignant cells have not been clearly elucidated yet. In this study, it was investigated the effects of ethyl methanol extract of G. furcata (MEGF) on the anti-proliferative effects of U937 human leukemia cells. MEGF treatment resulted in a concentration-dependent growth inhibition by including apoptosis, which could be proved by MTT assay, DAPI staining, agarose gel electrophoresis and flow cytometry analysis. The increase in apoptosis induced by MEGF treatment was correlated with up-regulation of pro-apoptotic Bax expression and down-regulation of anti-apoptotic Bcl-2 and Bcl-xL expression. MEGF treatment inhibited the levels of IAP family members such as XIAP and cIAP-1 and induced the proteolytic activation of caspase-3 and caspase-9, and a concomitant degradation of poly (ADP-ribose) polymerase and β-catenin. Taken together, these findings provide important new insights into the possible molecular mechanisms of the anti-cancer activity of G. furcata. (Cancer Prev Res 14, 274-281, 2009)
Key Words: Gloiopeltis furcata, U937, Apoptosis, Bax, Caspase
서 론
우리의 식생활과 밀접한 연관이 있는 많은 천연물들 이 암예방 및 항암작용이 있는 것으로 알려져 있지만 최 근 육상자원의 한계를 극복하기 위하여 해양생물 유래
천연 물질에 대한 관심이 매우 높아지고 있다.1) 그 중에 서도 식용이 가능한 해조류는 강력한 항산화 효과가 있 다고 잘 알려져 있으며, 혈중 콜레스테롤을 내려주고, 고 혈압과 혈액의 점도를 낮출 뿐 아니라 궤양을 예방하고, 항균작용을 가지는 등의 다양한 효과가 있는 것으로 보 고되어지고 있다.2∼4) 아울러 최근 해조류 추출물들의 암
세포주기 주기 교란과 연관된 세포사멸(apoptosis) 유도를 통한 항암작용 가능성에 관한 다양한 보고들이 이루어 지고 있다.5∼8)
암의 효과적인 예방과 치료에 있어서 비정상적인 세 포나 암세포의 apoptosis 유발은 많은 치료제의 표적이 되 고 있으며, apoptosis는 개체보존의 수준에서 손상된 세포 들의 제거를 위한 세포의 사멸 현상으로서 중요한 수단 이기도 하다.9) Apoptosis의 유발에 다양한 유전자의 활성 변화가 관여한다는 사실이 알려지면서 apoptosis와 연관 된 분자적 기전이 최근 많이 밝혀지고 있으며, apoptosis 는 특히 암세포의 생장, 증식의 억제와 암세포 파괴의 한 방법으로써 널리 연구되고 있다.10,11)
본 연구에서는 선행연구를 통하여 항돌연변이 효능이 우수하여 항암활성효능이 높을 것으로 예상되는 해조류 의 한 종류인 홍조식물 풀가사리과의 불등가사리(G.loio- peltis furcata)3,12,13)에 함유된 항암/암예방 후보물질의 도출 을 위하여 불등가사리의 메탄올 추출물이 인체 백혈병 세포의 증식에 미치는 영향을 조사하였으며, 백혈병세 포의 증식 억제 효과가 apoptosis 유발과 연관성이 있는지 의 여부를 조사하였다.
재료 및 방법 1. 실험 재료 및 세포배양
본 실험을 위하여 생명공학연구소(KRIBB, Taejeon, Korea)에서 분양받은 백혈병 U937세포는 10%의 우태아 혈청(fetal bovine serum, FBS, Gibco BRL, Grand Island, NY, USA)과 1%의 penicillin-streptomycin 등이 포함된 RPMI- 1640 배지(Gibco BRL)를 사용하여 37oC, 5% CO2 조건하 의 CO2 incubator에서 배양하였다. 불등가사리는 전라남 도 무안군에 위치한 (주)삼일물산에서 제공받았으며, 메 탄올 추출물(methanol extracts of G. furcata, MEGF)을 얻기 위하여 건조 후 메탄올을 첨가하고 37oC에서 진탕한 다 음 4시간 동안 3회 반복 추출하고 회전식 진공 농축기로 감압 농축시킨 후 동결 건조하였다.13)
2. MTT assay에 의한 세포 성장억제 조사
세포 배양용 6 well plate에 U937 세포를 적정량 분주하 여 24시간 동안 안정화시킨 다음 MEGF를 적정량 처리 하여 48시간 동안 배양한 후 각 well의 배지를 깨끗하게 제거하고 tetrazolium bromide salt (MTT, Amresco, Solon, OH, USA)를 0.5 mg/ml 농도가 되게 희석하여 2 ml씩 분 주하였다. 3시간 동안 CO2 incubator에서 빛을 차단하여 반응시킨 다음 MTT 시약을 제거하고 DMSO로 각 well에
생성된 formazin을 각각 동량으로 녹인 후 ELISA reader (Molecular Devices, Sunnyvale, CA, USA)를 이용하여 540 nm에서의 흡광도를 측정하였다. 측정은 모두 세 번을 하 였으며, 그에 대한 평균값과 표준 오차를 Sigma Plot 4.0 프로그램(SPSS Ins.)으로 구하였다.
3. DNA flow cytometry 분석
Apoptosis 유발의 정량적 분석을 위하여 정상 및 MEGF 을 처리한 배지에서 48시간 동안 배양시킨 U937 세포를 2,000 rpm에 5분간 원심분리하여 상층의 배지를 버리고 세포 pallet만 모아서 PBS 1 ml로 충분히 재부유 시킨 다 음 2,000 rpm으로 5분간 원심분리 하였다. 상층의 PBS만 버리고 남은 세포 pallet에 CycleTEST PLUS DNA REA- GENT KIT (Becton Dickinson, San Jose, CA, USA)를 사용하 여 고정 및 propidium iodide (PI, concentration, 50 μg/ml;
Sigma) 염색액을 처리하여 암실, 4oC에서 15분 동안 염색 하였다. 염색 후 DNA flow cytometry (Becton Dickinson, San Jose, CA, USA)에 대조군 및 MEGF 처리군을 적용시 켜 형광반응에 따른 histogram을 ModiFit LT (Becton Dick- inson) program으로 분석하였다.
4. DAPI staining에 의한 세포핵의 형태 관찰
MEGF 처리에 의한 U937 세포의 apoptosis 유발 여부 확인을 위한 핵의 형태적 변화를 관찰하기 위하여 정상 및 MEGF가 처리된 배지에서 48시간 동안 배양된 세포 를 모은 다음 37% formaldehyde 용액과 PBS를 1:9의 비 율로 섞은 fixing solution을 모아진 세포에 500 μl 첨가하 여 잘 섞어준 후, 실온에서 10분 동안 고정하였다. 2,000 rpm으로 5분간 원심 분리한 후 상층액을 제거하고 PBS 200 μl를 넣어서 충분히 섞은 다음 slide glass 위에 120 μl 정도 떨어뜨려 1,200 rpm에서 5분간 cytospin하였다.
PBS로 2∼3회 washing하고 slide glass를 건조시킨 후 4’, 6-diamidino-2-phenylindole (DAPI, Sigma, St. Louis, MO, USA) 용액을 세포가 고정된 slide glass 위에 적당량을 떨 어뜨린 후 빛을 차단하고 실온에서 염색시켰다. 약 10분 가량 염색시킨 후, 형광 현미경(Carl Zeiss)을 이용하여 400배의 배율로 각 MEGF 처리 농도에 따른 핵의 형태 변화를 관찰하였다.
5. Agarose gel 전기영동에 의한 DNA fragmenta- tion의 분석
특정 세포에 apoptosis가 유발되었을 때 관찰될 수 있는 DNA fragmentation의 분석을 위하여 정상 및 MEGF가 처 리된 배지에서 48시간 동안 배양된 U937 세포를 모아
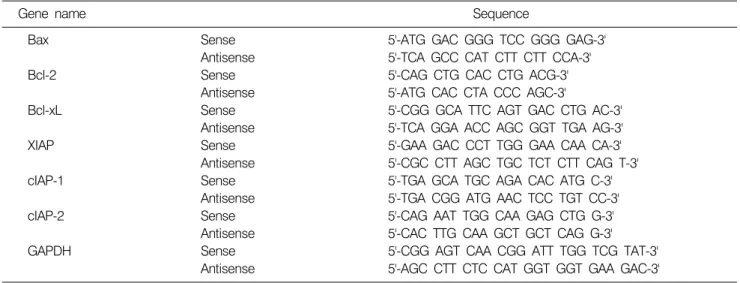
Table 1. Gene-specific primers for RT-PCR
Gene name Sequence
Bax Sense 5'-ATG GAC GGG TCC GGG GAG-3'
Antisense 5'-TCA GCC CAT CTT CTT CCA-3'
Bcl-2 Sense 5'-CAG CTG CAC CTG ACG-3'
Antisense 5'-ATG CAC CTA CCC AGC-3'
Bcl-xL Sense 5'-CGG GCA TTC AGT GAC CTG AC-3'
Antisense 5'-TCA GGA ACC AGC GGT TGA AG-3'
XIAP Sense 5'-GAA GAC CCT TGG GAA CAA CA-3'
Antisense 5'-CGC CTT AGC TGC TCT CTT CAG T-3'
cIAP-1 Sense 5'-TGA GCA TGC AGA CAC ATG C-3'
Antisense 5'-TGA CGG ATG AAC TCC TGT CC-3'
cIAP-2 Sense 5'-CAG AAT TGG CAA GAG CTG G-3'
Antisense 5'-CAC TTG CAA GCT GCT CAG G-3'
GAPDH Sense 5'-CGG AGT CAA CGG ATT TGG TCG TAT-3'
Antisense 5'-AGC CTT CTC CAT GGT GGT GAA GAC-3' lysis buffer [5 mM Tris-HCl (pH 7.5), 5 mM EDTA, 0.5%
Triton X-100]를 4oC에서 30분간 처리한 다음, 그 상층액 에 proteinase K (Sigma)를 0.5 mg/ml의 농도로 처리하여 50oC에서 3시간 동안 반응시켰다. 그 후 phenol:chlo- roform:isoamyl alcohol 혼합 용액(25:24:1, Sigma)을 첨 가하고 30분간 반응시킨 후 원심 분리하여 얻어진 상층 액에 적정량의 isopropanol (Sigma)과 5 M NaCl를 첨가한 다음 24시간 정도 4oC에서 반응시켰다. 분리된 DNA pellet에 RNase A가 적당량 들어있는 TE buffer를 첨가하 여 녹이고 gel loading dye (Bioneer, Daejeon, Korea)를 혼합 한 후 1.5% agarose gel을 이용하여 1시간 정도 50 V로 전 기영동하여 ethidium bromide (EtBr, Sigma)로 염색하고 ultra vilolet (UV) 하에서 관찰하였다.
6. Reverse transcription-polymerase chain re- action 분석
상기와 동일한 조건에서 준비된 U937 세포를 PBS로 세척하고 TRIzol reagent (Invitrogen Co., Carlsbad, CA, USA) 를 4oC에서 1시간 동안 처리하여 total RNA를 각 처리 농도 별로 분리하였다. 분리된 RNA를 정량한 후, 관련된 primer, DEPC water 그리고 ONE-STEP RT-PCR PreMix Kit (Intron, Korea)를 넣고 Mastercycler gradient (Eppendorf, Hamburg, Germany)를 이용하여 증폭하였다. 각 PCR 산물 들을 양적 차이를 확인하기 위하여 1× TAE buffer로 1%
agarose gel을 만들고 well 당 각각의 primer에 해당하는 PCR 산물에 6× agarose gel loading buffer (BIONEER, Korea) 를 섞어서 20 μl씩 well에 loading 한 후 100 V로 전기영 동을 하였다. 전기영동으로 DNA 분리가 끝난 gel을 EtBr 을 이용하여 염색한 후 UV 하에서 확인하고 Picture
works' photo enhancer를 이용하여 사진 촬영을 하였다.
RT-PCR을 이용하여 분석을 시도한 유전자의 종류는 Table 1에 나타낸 바와 같으며, 이때 housekeeping 유전자 인 glyceraldehyde-3-phosphate dehydrogenase (GAPDH) 유전 자를 internal control로 사용하였다.
7. Western blotting 분석
정상 및 MEGF가 처리된 배지에서 자란 U937 세포를 적당량의 lysis buffer [25 mM Tris-Cl (pH 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenymethylsulfonyl fluoride (PMSF), 5 mM dithiothreitol (DTT)]로 용해한 후, 고속원심분리기를 이용하여 14,000 rpm에 15분간 세포 내 잔사물을 분리시킨 다음 동량의 단백질을 sodium dodecyl sulphate (SDS)-polyacrylamide gel을 이용하여 전기 영동으로 분리하였다. 분리된 단백질을 nitrocellulose membrane (Schleicher and Schuell, Keene, NH, USA)으로 electroblotting에 의해 전이시킨 후, 특정 단백질에 대한 항체와 그에 대한 이차 항체 반응을 실시한 후 암실에서 Enhanced Chemiluminoesence (ECL) 용액(Amersham Life Sci- ence Corp., Arlington Heights, IL, USA)을 처리시킨 다음 X-ray film에 감광시켜 특정단백질의 양을 분석하였다.
본 실험에 사용된 항체들은 Santa Cruz Biotechnology Inc.
(Santa Cruz, CA, USA) 및 Calbiochem (Cambridge, MA, USA) 에서 구입하였으며, 2차 항체로 사용된 peroxidase-labeled donkey anti-rabbit 및 peroxidase-labeled sheep anti-mouse immunoglobulin은 Amersham Life Science에서 구입하였다.
8. In vitro caspase-3, -8 및 -9 활성의 측정 MEGF가 처리된 U937 세포의 caspase 활성 측정을 위한
Fig. 1. Inhibition of cell viability in human leukemia U937 cells after treatment with methanol extracts of Gloiopeltis furcata (MEGF). U937 cells were seeded at 1×105/ml in a 6-well plate and treated with variable concentrations of MEGF for 48 h. The growth inhibition was measured by the metabolic-dye-based MTT assay. The data shown are means±SD of three independent experiments.
Fig. 2. Apoptosis induction of U937 human leukemia cells by MEGF. (A) U937 cells were treated with MEGF for 48 h and then stained with DAPI solution. After 10 min incubation at room temperature, the cells were washed with PBS and nuclear morphology was photographed with a fluorescent microscope using blue filter. Magnification, ×400. (B) U937 cells were incubated with variable concentrations of MEGF for 48 h, then collected and DNA was extracted. The DNA fragmentations were separated on 1.5% agarose gel electrophoresis and visualized under UV light after staining with EtBr. (C) The cells grown under same condition as (B) were stained with PI for flow cytometry analysis. The percentages of cells with hypodiploid DNA (sub-G1 phase) contents represent the fractions undergoing apoptotic DNA degradation. Data are expressed as mean±SD of three independent experiments.
colorimetric assay kits는 R&D Systems (Minneapolis, MN, USA)에서 구입하였으며, 제시된 방법에 준하여 활성의 증감 여부를 조사하였다. 이를 위하여 사용된 기질은 caspase-3의 경우에는 Asp-Glu-Val-Asp (DEVD)-p-nitroani- line (pNA)이었고 caspase-8의 경우에는 Ile-Glu-Thr-Asp (IETD)-pNA이었으며, caspase-9은 Leu-Glu-His-Asp (LEHD)- pNA였다. 준비된 plate를 37oC에서 2시간동안 incubation 시킨 후 ELISA reader를 이용하여 405 nm의 흡광도를 이 용하여 반응의 정도를 측정하였다.
결과 및 고찰
1. MEGF의 처리에 의한 U937 세포의 증식 억제 및 apoptosis의 유발
U937 백혈병세포의 증식에 미치는 불등가사리 메탄 올 추출물(MEGF)의 영향을 비교하기 위하여 다양한 농 도(0∼75 μg/ml)로 48시간동안 처리한 후 MTT assay를 실시하였다. Fig. 1에 나타낸 바와 같이 MEGF의 처리 농 도가 증가할수록 U937 세포의 생존율은 현저하게 감소 되어 30 μg/ml 이상 처리군에서 50% 이하의 생존율을 보였다. 이는 형태학적 관찰에서 세포의 밀도 감소와 함 께 세포의 사멸에 의한 잔존물들이 배양액 상에 처리 농 도 의존적으로 점차 늘어나는 것을 확인할 수 있었다.
MEGF의 처리에 의한 증식 억제 효과가 apoptosis 유발과 상관성이 있는지를 조사하기 위하여 DAPI 염색에 의한
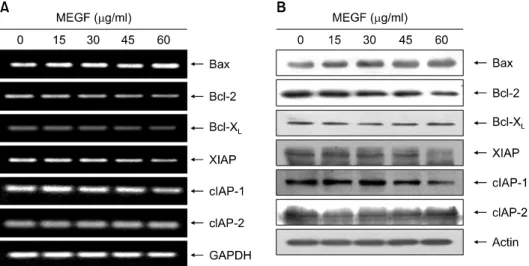
Fig. 3. Effects of MEGF on the Bcl-2 and IAP family members expression in U937 cells. (A) After 48 h incubation with MEGF, total RNAs were isolated and reverse-transcribed. The resulting cDNAs were subjected to PCR with the indicated primers, and the reaction products were subjected to electrophoresis in 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells were lysed and then cellular proteins were separated by 10% SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
핵의 형태적 변화, agarose gel 전기영동에 의한 DNA 단 편화 및 flow cytometry 분석에 의한 sub-G1기 세포의 빈 도 변화 등을 조사하였다. 먼저 Fig. 2A에 나타낸 것과 같이 apoptosis가 유발된 세포에서 관찰되는 chromatin condensation에 의한 apoptotic body의 출현이 MEGF 처리 농도 의존적으로 증가됨을 확인할 수 있었고, Fig. 2B의 agarose gel 전기영동에 의하여 MEGF의 처리 농도 증가에 따라 DNA 단편화가 증가되었음을 알 수 있었다. 또한 Fig. 2C의 flow cytometry 결과에서 나타난 것과 같이 정상 배지에서 자란 U937 세포에 비하여 MEGF가 처리된 배 지에서 배양된 U937 세포에서 처리 농도 의존적으로 apoptosis가 일어난 세포에 해당되는 sub-G1기에 속하는 세포의 빈도가 상당량 증가하는 것을 확인하였다. 이와 같은 결과는 MEGF의 처리에 의한 U937 세포의 증식 억 제 효과가 apoptosis의 유발과 직접적인 연관성이 있음을 의미하여 주는 것이다.
2. Bcl-2 family의 발현에 미치는 MEGF의 영향
Apoptosis의 조절인자 중 Bcl-2 family에 속하는 몇 가지 인자들은 apoptosis 유발 조절에 가장 대표적인 유전자로 알려져 있는데, 그중 Bcl-2 및 Bcl-xL 등은 anti-apoptotic 분자로서 apoptosis의 유발을 억제하는 기능을 가지며, Bax는 pro-apoptotic 분자로 Bax 단백질 발현의 증가는 apoptosis의 유발과 관계가 있다. 이들 두 유전자는 세포 내 소기관 중 mitochondria로부터의 cytochrome c를 유리시
켜 cysteine-related protease인 caspase, 종양억제 유전자인 p53, DNA의 단편화와 연관된 endonuclease 등의 활성을 조절하며, 이들은 서로 dimer의 형태로 존재하며 그들의 발현 수준에 변화가 초래되면 apoptosis가 유발되는 것으 로 알려져 있다.10,14) 따라서 U937 세포에서 MEGF에 의 한 apoptosis 유발에 이들 유전자가 관련되어 있는지의 여 부를 RT-PCR 및 Western blot 분석으로 조사하였다. Fig.
3에 나타낸 바와 같이 anti-apoptotic 인자인 Bcl-2 및 Bcl-xL의 발현은 mRNA 및 단백질 수준에서 모두 MEGF 처리 농도의 증가에 따라 발현의 정도가 감소되었으나, pro-apoptotic 인자인 Bax의 경우 MEGF 처리 농도 증가에 따라 전사 및 번역 수준에서 발현이 모두 증가되었다.
이는 MEGF의 처리에 의한 U937 세포의 생존율 저하에 따른 apoptosis 유발에 Bcl-2 family의 발현 변화가 중요한 역할을 하고 있음을 보여주는 결과이다.
3. IAP family의 발현에 미치는 MEGF의 영향
Apoptosis 조절에 관여하는 또 다른 인자 중 IAP family 에 속하는 여러 유전자 산물은 caspase와의 직접적인 결 합을 통하여 그들의 apoptotic 활성을 억제할 수 있을 것 으로 밝혀져 있다.15,16) 따라서 본 연구에서는 MEGF 처리 에 의한 U937 세포의 apoptosis 유발에 이들 IAP family가 관여하는지의 여부를 조사하였다. Fig. 3에 나타난 바와 같이 MEGF 처리에 따라서 조사된 IAP family의 3가지 인 자들 중, cIAP-2의 발현은 전사 및 번역 수준에서 큰 변
Fig. 4. Activation of caspases and degradation of caspase-3 target proteins by MEGF treatment in human leukemia U937 cells. (A) After 48 h incubation with MEGF, U937 cells were lysed, and cellular proteins were separated by SDS-polyacryla- mide gel and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies.
Proteins were visualized using an ECL detection system. Actin was used as an internal control. (B) U937 cells were treated with indicated concentrations of MEGF for 48 h, collected and then lysed. Aliquots (150 μg proteins) were incubated with DEVD-pNA, IETD-pNA and LEHD-pNA for caspase-3, cas- pase-8 and caspase-9 activity, respectively, at 37oC for 3 h.
The released fluorescent products were measured. The data shown are means±SD of three independent experiments.
화가 없었으나, XIAP 및 cIAP-1은 전사 및 번역 수준에서 모두 MEGF의 처리 농도 증가에 따라 발현의 정도가 현 저히 감소되었다. 이러한 결과는 MEGF의 처리에 의한 U937 세포의 apoptosis 유발에 IAP family 인자들의 발현 감소에 따른 caspase 효소들의 활성이 증가할 수 있음을 보여 주는 것이다.
4. Caspase의 활성에 미치는 MEGF의 영향
Caspase protease라는 효소는 apoptosis 유발에 가장 중요 한 조절인자로서 작용하는데, 이들 family에 속하는 단백 질들은 세포에서 핵과 mitochondria의 외막에 불활성 상 태로 존재하며, Bcl-2/Bax family 발현의 변화에 따라 이들 의 활성도가 조절될 수 있다.17,18) 이들은 proenzyme 형태 로 존재하다가 apoptosis 유도를 활성화 시키는 신호에 의 해 활성화된 protease로 전환되어 직접 또는 간접적으로 세포 내에 존재하는 많은 표적 단백질의 분해에 관여하 는 것으로 알려져 있다. 따라서 본 연구에서는 MEGF 처 리에 의한 U937 세포의 apoptosis 유발에 이들 caspase의 활성 변화가 관여하는지의 여부를 조사하였다. 이를 위 하여 3가지 종류의 caspase (caspase-3, -8 및 -9)를 조사하였 는데, 이 중 caspase-8은 death receptor와 연관된 extrinsic pathway의 과정에서, caspase-9는 미토콘드리아와 연관된 intrinsic pathway에서 initiation caspase로 작용한다. 그리고 caspase-3은 caspase-9의 활성에 의하여 활성화되거나 자가 활성에 의하여 apoptosis의 최종적인 과정에서 관여한
다.11,19) 먼저 Western blot analysis에 의한 3가지 종류의
caspase 단백질 발현의 정도를 비교한 결과는 Fig. 4에 나 타낸 바와 같으며 caspase-3의 경우 MEGF의 처리에 따라 활성형 caspase-3의 발현이 매우 증가되었다. 그리고 caspase-8의 경우도 활성형 단백질의 발현이 MEGF 처리 농도의 증가에 따라 발현의 정도가 증가되었으나 caspase-9의 활성형 단백질의 발현은 크게 증가되지 않았 다. 이러한 단백질 수준에서의 결과를 재확인하기 위하 여 in vitro caspase 활성을 정량적으로 조사한 결과는 Fig.
4B에 나타낸 바와 같다. Fig. 4B의 결과에서 알 수 있듯이 MEGF의 처리 농도 증가에 따라 caspase-3의 활성이 가장 높게 나타났으며, caspase-8의 활성도 다소 증가되었지만 caspase-9의 경우 상대적으로 낮게 나타나 Western blot analysis 결과와 유사한 경향성을 보여주었다.
한편 활성화된 caspase는 세포의 정상적인 생존에 필수 적인 세포 내 주요 단백질들을 분해할 수 있으며, 단편화 가 일어난 이들 단백질들은 apoptosis가 유발되었다는 표 지자로서 활용이 된다. 본 연구에서는 대표적인 caspase-3 의 기질 단백질에 해당되는 poly (ADP-ribose) polymerase
(PARP) 및 β-catenin의 발현 변화를 조사하였는데, PARP 단백질은 eukaryotic nuclear enzyme family로서 DNA repair, gene transcription, cell cycle progression, chromatin function, genomic stability 및 cell death를 조절하는 것으로 알려져 있으며,20,21) cell-cell adhesion 및 Wnt signaling에 관여하는
E-cadherin–associated 단백질로서 세포 내 골격 유지, 다양 한 부착성 세포의 전사 조절 및 세포 유착에 관여하는 β-catenin도 caspase의 표적 단백질로서 apoptosis 조절에 관여하는 것으로 알려져 있다.22) Fig. 4A에 제시한 것처 럼 Western blotting 분석을 통하여 조사한 결과 단편화된 PARP 및 β-catenin 단백질의 발현이 MEGF 처리 농도의 증가에 따라 모두 증가되었음을 알 수 있었다. 이상의 결과는 MEGF 처리에 따른 U937 세포의 apoptosis 유발은 caspase의 활성을 통한 표적단백질의 단편화에 의하여 이 루어지고 있음을 보여 주는 것이다.
결 론
본 연구에서는 인체백혈병 U937 세포의 증식에 미치 는 불등가사리 메탄올 추출물(MEGF)의 영향을 조사하였 다. MEGF의 처리 농도의 증가에 따라 농도의존적으로 U937 세포의 생존율은 감소되었으며, 이러한 현상이 apoptosis 유발과 연관성이 있었음을 염색질 응축에 따른 apoptotic body의 출현 증가, DNA 단편화 및 flow cyto- metry 분석에 따른 sub-G1기 세포의 빈도 증가로 확인을 하였다. MEGF 처리에 의한 apoptosis 유발에 관여하는 유 전자들의 발현 변화를 RT-PCR 및 Western blot 방법으로 조사한 결과, Bcl-2 family에 속하는 anti-apoptotic 인자인 Bcl-2 및 Bcl-xL의 발현은 전사 및 번역 수준에서 감소되 었고, pro-apoptotic Bax의 발현은 증가되었으며, IAP family 단백질들의 발현도 감소되었다. 또한 caspase의 활 성 증가외 더불어 PARP 및 β-catenin과 같은 caspase-3의 기질 단백질들의 분해 등이 MEGF에 의한 U937 세포의 apoptosis 유도과정에서 관찰되었다. 이상의 결과들은 MEGF의 항암작용을 이해하는데 중요한 자료가 될 것이 고 나아가 MEGF 내 함유된 생리활성 물질의 분리 및 항 암작용 연구를 위한 중요한 기초 자료로 활용될 것이다.
감사의 글
본 연구는 지식경제부ㆍ부산광역시 지원 지역혁신센 터사업 동의대학교 블루바이오 소재 개발 및 실용화 지 원 센터(RIC08-06-07) 지원에 의하여 이루어진 결과입니다.
참 고 문 헌
1) Aneiros A, Garateix A. Bioactive peptides from marine sources: pharmacological properties and isolation procedures.
J Chromatogr B Analyt Technol Biomed Life Sci 8, 41-53, 2004.
2) Ara J, Sultana V, Qasim R, Ahmad VU. Hypolipidaemic activity of seaweed from Karachi coast. Phytother Res 16, 479- 483, 2002.
3) Heo SJ, Park EJ, Lee KW, Jeon YJ. Antioxidant activities of enzymatic extracts from brown seaweeds. Bioresour Technol 96, 1613-1623, 2005.
4) Yuan YV, Walsh NA. Antioxidant and antiproliferative acti- vities of extracts from a variety of edible seaweeds. Food Chem Toxicol 44, 1144-1150, 2006.
5) Kim JY, Yoon MY, Cha MR, Hwang JH, Park E, Choi SU, Park HR, Hwang YI. Methanolic extracts of Plocamium telfairiae induce cytotoxicity and caspase-dependent apoptosis in HT-29 human colon carcinoma cells. J Med Food 10, 587- 593, 2007.
6) Rocha FD, Soares AR, Houghton PJ, Pereira RC, Kaplan MA, Teixeira VL. Potential cytotoxic activity of some Brazilian seaweeds on human melanoma cells. Phytother Res 21, 170- 175, 2007.
7) Funahashi H, Imai T, Mase T, Sekiya M, Yokoi K, Hayashi H, Shibata A, Hayashi T, Nishikawa M, Suda N, Hibi Y, Mizuno Y, Tsukamura K, Hayakawa A, Tanuma S. Seaweed prevents breast cancer? Jpn J Cancer Res 92, 483-487, 2001.
8) Tokudome S, Kuriki K, Moore MA. Seaweed and cancer prevention. Jpn J Cancer Res 92, 1008-1009, 2004.
9) Barisic K, Petrik J, Rumora L. Biochemistry of apoptotic cell death. Acta Pharm 53, 151-164, 2003.
10) Liu S, Seidel-Dugan C. In search of p53 target genes for the therapeutic manipulation of cancer. Curr Opin Drug Discov Devel 9, 176-183, 2006.
11) Schultz DR, Harrington Jr WJ. Apoptosis: programmed cell death at a molecular level. Semin Arthritis Rheum 32, 345-369, 2003.
12) Ham SS, Lee SY, Choi M, HwangBo HJ. Antimutagenicity and cytotoxicity effect of Woorimil wheat flour extracts added with wild herb and seaweed powder. J Korean Soc Food Sci Nutr 27, 1177-1182, 1998.
13) Park SY, Jung BM, Choi YH, Bae SJ. Growth inhibition effects of cancer cell lines by Gloiopeltis furcata fractions in vitro.
J Korean Soc Food Sci Nutr 34, 771-775, 2005.
14) Bettaieb A, Dubrez-Daloz L, Launay S, Plenchette S, Rebe C, Cathelin S, Solary E. Bcl-2 proteins: targets and tools for chemosensitisation of tumor cells. Curr Med Chem Anticancer Agents 3, 307-318, 2003.
15) de Graaf AO, de Witte T, Jansen JH. Inhibitor of apoptosis proteins: new therapeutic targets in hematological cancer?
Leukemia 18, 1751-1759, 2004.
16) Wrzesien-Kus A, Smolewski P, Sobczak-Pluta A, Wierzbowska A, Robak T. The inhibitor of apoptosis protein family and its antagonists in acute leukemias. Apoptosis 9, 705-715, 2004.
17) Philchenkov A, Zavelevich M, Kroczak TJ, Los M. Caspases and cancer: mechanisms of inactivation and new treatment modalities. Exp Oncol 26, 82-97, 2004.
18) Chang HY, Yang X. Proteases for cell suicide: functions and
regulation of caspases. Microbiol Mol Biol Rev 64, 821-846, 2000.
19) Jin Z, El-Deiry WS. Overview of cell death signaling path- ways. Cancer Biol Ther 4, 139-163, 2005.
20) Kaufmann SH, Desnoyers S, Ottaviano Y, Davidson NE, Poirier GG. Specific proteolytic cleavage of poly (ADP-ribose) polymerase: an early marker of chemotherapy-induced apop-
tosis. Cancer Res 53, 3976-3985, 1993.
21) Schreiber V, Dantzer F, Ame JC, de Murcia G. Poly (ADP- ribose): novel functions for an old molecule. Nat Rev Mol Cell Biol 7, 517-528, 2006.
22) Fukuda K. Apoptosis-associated cleavage of β-catenin in human colon cancer and rat hepatoma cells. Int J Biochem Cell Biol 31, 519-529, 1999.