164 책임저자:최영현, 614-052, 부산시 진구 양정동 산 45번지
동의대학교 한의과대학 생화학교실 Tel: 051-850-7413, Fax: 051-853-4036 E-mail: [email protected]
접수일:2010년 4월 7일, 1차수정일: 2010년 4월 12일, 2차수정일: 2010년 4월 15일, 게재승인일:2010년 4월 19일
Correspondence to:Yung Hyun Choi
Department of Biochemistry, Dongeui University College of Oriental Medicine, San 45, Yangjung-dong, Busanjin-gu, Busan 614-052, Korea Tel: +82-51-850-7413, Fax: +82-51-853-4036
E-mail: [email protected]
톳 메탄올 추출물에 의한 인체 폐암 세포의 Apoptosis 유발
동의대학교 한의과대학 1병리학교실, 2생화학교실,
3대학원 바이오물질제어학과 및 블루바이오 소재개발센터
박신형1ㆍ신동역2ㆍ엄현섭1ㆍ지규용1ㆍ최영현2,3
Apoptosis Induction of Human Lung Cancer Cells by Methyl Alcohol Extracts of Hizikia fusiforme
Shin-Hyoung Park1, Dong Yeok Shin2, Hyun Sup Eom1, Gyoo Yong Chi1 and Yung Hyun Choi2,3 Departments of 1Pathology and 2Biochemistry, College of Oriental Medicine,
3Department of Biomaterial Control (BK21 Program), Graduate School and Blue-Bio Industry Regional Innovation Center, Dongeui University, Busan 614-052, Korea
Hizikia fusiforme, a brown alga, has been reported to possess a wide range of beneficial activities including immuno-modulating, anti-oxidants, anti-coagulants, and anti-carcinogenic effects. According to recent studies, H. fusiforme can inhibit the growth of cancer cell by inducing apoptosis in a certain cancer lines in vitro. However, the molecular mechanism how it acts as an inducer of apoptosis is not fully understood. In this study, we investigated the apoptotic effects of H. fusiforme methyl alcohol extracts (MEHF) in A549 and NCI-H460 human lung carcinoma cell lines. It was shown that MEHF induced the cell growth inhibition in a dose-dependant manner and it was more significant in NCI-H460 cells.
There also appeared increased sub-G1 phase DNA content, chromatin condensation, as well as DNA fragmentation in MEHF-treated cells, which are indicators of apoptosis. MEHF activated the caspase-8 via up-regulating Fas, FasL and DR5 expression and cleavage of Bid protein. MEHF also up-regulated the pro-apoptotic Bax levels and down-regulated anti-apoptotic Bcl-xL expression, which was associated with the loss of mitochondrial membrane potential (MMP) and activation of caspase-9 and caspase-3.
Taken together, these findings suggest that H. fusiforme might be useful for clinical application to treat human lung cancer. (Cancer Prev Res 15, 164-171, 2010)
Key Words: Hizikia fusiforme, Human lung cancer, Apoptosis, Caspase
서 론
톳(Hizikia fusiforme)은 갈조식물인 모자반과의 해조류로 서 남해안과 제주도에 주로 서식하는 다년생 해조류이 다. 톳은 칼슘, 요오드, 철 등의 무기질과 식이섬유소 중 알긴산을 다량 포함하고 있는 저열량 식품으로서, 고지
혈증이나 당뇨병에 효과적인 것으로 알려져 있다.1) 최근 연구에 따르면 톳은 항암, 항산화, 항염증, 항돌연변이, 항혈액응고 효과가 있으며, 그밖에 대사촉진, 면역활성 효과도 보고되고 있다.2∼5) 특히 에탄올이나 메탄올 등의 유기용매로 추출한 톳 시료는 탁월한 항산화 및 항암 효 과를 보였지만,6∼10) 항암효과에 대한 분자생물학적 기전 에 관한 구체적인 연구는 여전히 미비한 실정이다.
한편 apoptosis는 해당 세포가 준비된 프로그램에 따라 능동적으로 자살하는 과정으로서, 1970년대 초반 처음 제시된 후 20세기 후반부터 폭발적인 연구가 이루어져 왔다. 영양소의 결핍과 독물, 외상 등 외적 요인에 의해 세포가 급팽창해 파괴되는 준비되지 않은 죽음의 형태 인 necrosis와는 달리 apoptosis는 세포질의 응축, 세포막의 수포화, DNA의 절단을 특징으로 하는 질서정연한 프로 그램에 의해 세포가 죽음에 이르게 된다.11) 현재까지 밝혀 진 apoptosis의 기전은 크게 미토콘드리아 경로와 죽음 수 용체 경로로 나누어진다. 죽음 수용체 경로는 death-in- ducing signaling complex (DISC)를 형성하여 caspase-8을 활 성화시키며,12,13) 미토콘드리아 경로는 Bax와 Bak이 미토 콘드리아 외막으로 삽입되어 미토콘드리아 외막 투과성 이 증가하고 이로 인해 미토콘드리아 외막과 내막 사이에 존재하는 cytochrome c가 세포질로 방출되면서 caspase-9 및 apoptosis activating factor-1 (Apaf-1)과 apoptosome complex를 형성한다.14∼16) Caspase-8 및 -9와 같은 죽음개시 caspase가 활성화되면, 대표적인 죽음 실행 caspase인 caspase-3이 절 단 및 활성화되고, 그 기질인 poly (ADP-ribose) polymerase (PARP) 및 inhibitor of caspase activated DNase (ICAD) 등과 같은 단백질의 단편화를 동반한 apoptosis를 유도하게 된 다.17) 또한 caspase-8이 Bcl-2 family 단백질인 Bid를 잘라 tBid로 만든 결과 미토콘드리아의 외막투과성을 증가시 키면서 두 경로가 연결되며,18) 한 경로를 구성하는 요소 들이 다른 경로에 영향을 미치기도 한다.19) 따라서 특정 조건에서의 암세포 apoptosis 유발은 암세포의 증식억제 에 의한 암 치료법의 주요 방법으로 특정 항암활성을 가 지는 물질들의 항암기전 해석을 위한 기본적인 과정이 라 할 수 있다.
본 연구에서는 그동안 알려진 톳의 항암활성에 대한 부가적인 자료의 제시를 위하여 톳의 메탄올 추출물에 의한 암세포의 apoptosis 유발 기전에 관한 연구를 시도하 였다. 이를 위하여 인체 폐암세포를 대상으로 조사한 결 과, 톳 메탄올 추출물은 폐암세포의 증식을 강력히 억제 하였으며, 이는 apoptosis 유발에 의한 것임을 확인하였 다. 아울러 톳 메탄올 추출물은 apoptosis 유발에서 나타 나는 다양한 유전자들의 발현 및 활성 변화에 대한 유의 적인 결과를 얻었기에 이를 보고하고자 한다.
재료 및 방법 1. 실험 재료
본 연구에서 사용한 톳은 울산시 정자동 해변에서 채 집하여 하루 동안 흐르는 물에 담궈 염분을 제거하고 그
늘에서 일주일간 말린 후 미세분말로 만들었다. 톳의 메 탄올 추출물(methanol extract of H. fusiforme, MEHF)은 메탄 올 1 l에 톳 분말 50 g을 첨가하여 추출한 것으로, 60oC, 140 rpm으로 5일 동안 incubation 시켰다. 그 후 침전물을 제거하고 상층액만 분리해내어 다시 5일 동안 60oC, 140 rpm으로 건조시킨 후 생긴 결정을 Dimethyl sulfoxide (DMSO, Sigma Chemical Co., St. Louis, MO, USA)에 용해시 켰다. Western blot에서 사용된 1차 항체와 2차 항체는 모 두 Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA)에서 구입하였다.
2. 세포 배양
본 연구에 사용된 A549 및 NCI-H460 인체 폐암세포는 한국생명공학연구소(Daejeon, Korea)에서 분양받았다. 세 포의 배양을 위해 90%의 RPMI-1640 (Gibco BRL, Grand Island, NY, USA)에 10% fetal bovine serum (FBS)와 1%
penicillin streptomycin (Biofluids, Rockville, MD, USA)을 첨 가하여 사용하였으며, 세포는 37oC 및 5% CO2 조건 하에 서 배양하였다.
3. MTT assay를 통한 세포 생존율 측정
A549 및 NCI-H460 세포에 MEHF를 적정 농도로 48시 간 동안 처리한 후, 배지를 모두 제거하고 tetrazolium bromide salt (MTT, Sigma)를 0.5 mg/ml 농도가 되게 배지 로 희석하여 분주하고 2시간 동안 다시 incubation 시켰 다. 그 후 MTT 시약을 제거하고 DMSO로 각 well의 formazan을 녹인 후 96 well plate에 분주하여 ELISA reader (Molecular Devices, Sunnyvale, CA, USA)에서 540 nm로 흡 광도를 측정하였다.
4. DAPI staining을 통한 핵의 관찰
MEHF를 48시간 처리한 세포들을 모아 3.7% parafor- maldehyd (Sigma)로 상온에서 10분 동안 고정하였다. 그 후 상층액을 제거하고 적정량의 PBS로 suspension하여 cytospin으로 세포를 slide glass 위에 부착시킨 후, 4,6-dia- midino-2-phenylindole (DAPI) solution (Sigma)으로 약 30분 간 염색시켜 형광현미경(Carl Zeiss, Germany)을 이용하여 400배의 배율로 핵의 형태를 관찰하였다.
5. Flow cytometry을 통한 세포주기 및 MMP 변화의 정량적 분석
적정농도의 MEHF가 48시간 처리한 세포들을 모아 Cycle TEST PLUS (Becton Dickinson, San Jose, CA, USA) solution A 및 B를 상온에서 각각 10분씩 처리한 후, Cycle
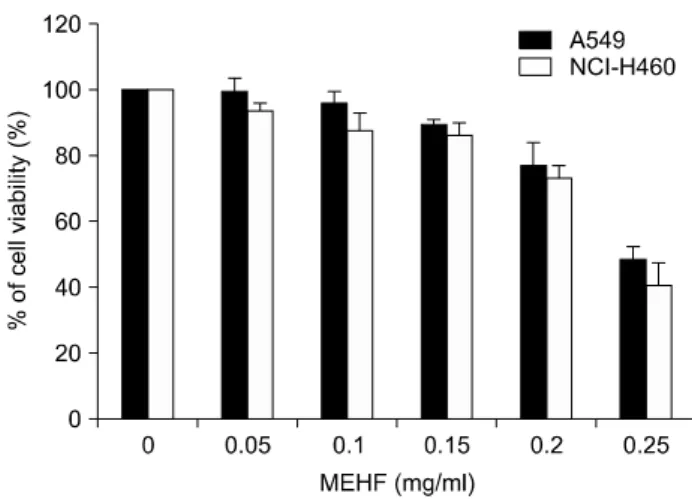
Fig. 1. Anti-proliferative effects of H. fusiforme methyl alcohol extracts (MEHF) in A549 and NCI-H460 human lung cancer cells. Cells were seeded in the 6 well plate at an initial density of 1×105 cells per well. After 24 h of stabilization, the cells were treated with EMHF with the indicated concentrations for 48 h and then MTT assay was performed. Each point represents the mean±SD of three independent experiments.
TEST PLUS solution C를 처리하여 4oC에서 10분간 염색하 였다. 그 후 DNA flow cytometer (BD FACSCalibur)를 사용 하여 세포주기의 histogram을 분석하였다. Mitochondrial membrane potential (MMP) 변화의 분석은 MEHF가 처리된 암세포들을 대상으로 10μM로 희석한 5,5',6,6'-tetrachloro- 1,1',3,3'-tetraethylbenzimidazolylcarbocyanine iodide (JC-1) dye (Calbiochem, San Diego, CA, USA)로 30분간 염색한 후 역 시 DNA flow cytometer를 이용하여 MMP의 변화 정도를 정량적으로 비교하였다.
6. DNA fragmentation 분석
MEHF가 처리된 세포들에 lysis buffer [5 mM Tris-HCl (pH 7.5), 5 mM EDTA, 0.5% Triton X-100] 800μl를 넣고 4oC에서 1시간 동안 lysis시켰다. 그 후 상층액 750μl를 분리하여 proteinase K (Sigma)를 0.5 mg/ml가 되도록 처리 하고 phenol:chloroform:isoamyl alcohol 혼합 용액(Sigma) 을 750μl 넣은 후 30분간 교반시켰다. 다시 원심분리하 여 상층액만 분리한 후 isopropanol (Sigma) 300μl과 5 M NaCl 100μl를 첨가하여 4oC에서 overnight 하였다. 이것 을 원심분리시켜 남은 pellet을 RNase A가 포함된 DEPC 로 녹인 후 6× gel loading dye (Bioneer, Daejeon, Korea)를 섞어 2% agarose gel에 loading하여 50 V에서 1시간 정도 전기 영동하였다. 그 후 ethidium bromide (EtBr, Sigma) 용 액에 15분간 염색하여 UV 하에서 DNA ladder를 관찰하 였다.
7. 단백질의 분리, 전기영동 및 Western blotting
동일 조건에서 준비된 세포들을 모아 lysis buffer [25 mM Tris-Cl (pH 7.5), 250 mM NaCl, 5 mM EDTA, 1%
NP-40, 1 mM phenymethylsulfonyl fluoride (PMSF), 5 mM dithiothreitol (DTT)]에 1시간 가량 lysis시킨 후 Laemmli sample buffer를 동량으로 섞어 SDS-polyacrylamide gel에 loading하여 전기영동시켰다. 이 acrylamide gel을 electro- blotting을 통해 2시간 동안 nitrocellulose membrane (Schleicher and Schuell, Keene, NH, USA)에 옮긴 후 5%
skim milk로 1시간 동안 blocking하고 1차 항체를 붙여 4oC 에서 overnight하였다. 그 후 PBS-T로 세척하고 상온에서 1시간가량 2차 항체를 붙인 후 다시 세척하고 Enhanced Chemilunimoecence (ECL) 용액(Amersham Life Science Corp., Arlington Height, IL, USA)을 처리한 후 암실에서 Kodak X-ray에 감광시켰다.
8. Caspase activity 측정
준비된 암세포들을 대상으로 caspase-3, -8 및 -9 Colori-
metric Assay Kits (R&D Systems, Minneapolis, MN, USA)의 사용방법에 따라 단백질을 추출 및 정량하여 150μg의 단백질을 96 well plate에 주입하였다. 그 후 caspase activity kit 2× reaction buffer 50μl에 1 M DTT를 10μl/ml이 되도 록 섞어 96 well plate에 첨가하고, 여기에 4 mM 기질을 5μl씩 첨가하였다. 이 때 caspase-3의 기질은 Asp-Glu-Val- Asp (DEVD)-p-nitroaniline (pNA), caspase-8의 기질은 Ile- Glu-His-Asp (IETD)-pNA, caspase-9의 기질은 Leu-Glu-His- Asp (LEHD)-pNA였다. 그 후 37oC에서 2시간 동안 incubation시킨 후 ELISA reader를 이용하여 405 nm에서 흡광도를 측정하였다.
결과 및 고찰
1. MEHF가 A549 및 NCI-H460 폐암세포의 생존율 및 형태에 미치는 영향
A549 및 NCI-H460 폐암세포의 증식에 미치는 MEHF 의 영향을 조사하기 위하여 적정 농도를 48시간 동안 처 리한 후 MTT assay를 실시하였다. Fig. 1의 결과에 제시한 바와 같이 MEHF의 처리 농도가 증가할수록 생존율의 감소를 확인하였는데, 0.2 mg/ml MEHF가 처리된 경우 A549 세포는 76%, NCI-H460 세포는 72% 정도로 나타났 으며, 0.25 mg/ml MEHF 처리군에서는 각각 48% 및 40%
의 생존율을 보였다. 다음은 A549 및 NCI-H460 세포의 형태에 MEHF가 어떤 영향을 미치는지 알아보기 위해
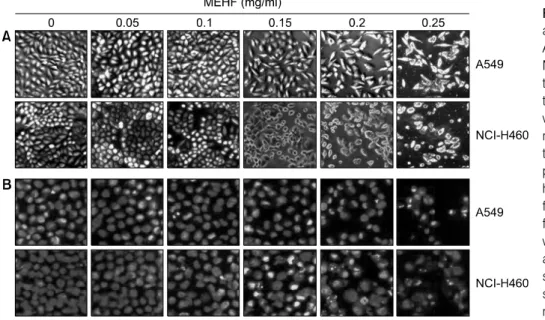
Fig. 2. Morphological changes and apoptotic body formation of A549 and NCI-H460 cells by MEHF treatment. (A) After treating the cells with various concen- trations of MEHF for 48 h, they were observed using an inverted microscope (original magnifica- tion, 200×). (B) For nuclear mor- phology, MEHF treated cells were harvested and spin down. After fixing with 3.7% paraformaldehyde for 10 min, the cells were stained with a DAPI solution for 10 min and stained nuclei were then ob- served under a fluorescent micro- scope using a blue filter (original magnification, ×400).
MEHF를 농도별로 처리하여 48시간 배양한 후 현미경으 로 관찰하였다. Fig. 2A에 나타난 바와 같이 MEHF의 처 리 농도가 증가될수록 부착된 세포의 밀집도가 처리 농 도 의존적으로 감소하였고, 배지 상에 부유하는 세포의 수가 증가하여 부착력 상실에 의한 세포의 죽음 유도 효 과가 있을 것으로 기대되었다. 이러한 세포의 죽음이 apoptosis에 의한 것인지 확인하기 위하여 핵을 특이적으 로 염색하는 DAPI 염색을 시행한 결과 전형적인 apoptosis가 유발된 세포에서 관찰되는 DNA 단편화에 의 한 apoptotic body가 MEHF 처리 농도 의존적으로 증가하 였다. 특히 A549 세포에서는 0.25 mg/ml 처리군에 이르 러서야 apoptotic body가 부분적으로 나타나는 반면, NCI-H460 세포에서는 0.15 mg/ml 처리군에서부터 apop- totic body가 출현하여 0.25 mg/ml 처리군에서는 매우 증 가되었다. 이들 결과는 MEHF가 A549 세포보다 NCI-H460 세포에 대해 더욱 탁월한 세포사멸 효과를 가진다는 것 을 의미한다.
2. MEHF 처리에 의한 NCI-H460 세포의 sub-G1 상 승 및 DNA 단편화
MEHF에 의한 apoptosis 유도 효과가 NCI-H460 세포에 서 높게 나타났으므로, apoptosis 유발의 또 다른 직접적 인 증거가 되는 DNA의 단편화 유도 여부를 NCI-H460 세포를 대상으로 확인하였다. Fig. 3A에서 나타낸 바와 같이 MEHF 처리 농도의 증가에 따라 DNA 단편화의 정 도가 매우 증가되었으며, 이러한 apoptosis 유발의 정도를 정량적으로 비교하기 위하여 MEHF를 농도별로 처리하 여 48시간 배양한 후 DNA flow cytometry로 cell cycle 분포
도를 분석한 결과 Fig. 3B에 나타난 바와 같이 apoptosis를 일으킨 세포군을 의미하는 sub-G1기의 빈도가 약 20%까 지 증가하였다. 이상의 결과는 MEHF에 의한 NCI-H460 세포의 죽음이 apoptosis에 의한 것임을 의미하며, MEHF 에 의한 암세포의 증식억제는 apoptosis 유도와 밀접하게 관련되어 있음을 알 수 있었다.
3. 죽음 수용체 경로 유전자의 발현에 미치는 MEHF의 영향
죽음 수용체 경로, 즉 외인성 경로는 죽음 신호를 전달 하는 ligand와 막수용체 사이의 상호작용에 의한 것으로 서 지금까지 잘 알려진 ligand/수용체에는 FasL/FasR, Apo2L/DR4, Apo2L/DR5, TRAIL/DR4, TRAIL/DR5 등이 있
다.12,13) Ligand가 수용체와 결합하면 세포질 내의 FADD
나 TRADD와 같은 adapter protein이 결합하여 죽음 유도 신호 복합체(DISC. death induciing signaling complex)를 형 성하고 그 결과 procaspase-8을 자가분해 및 활성화시킨
다.12,20) MEHF에 의한 NCI-H460 폐암세포의 apoptosis 유
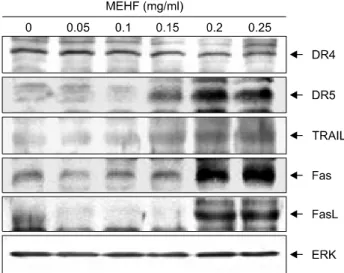
도에 이와 같은 죽음 수용체 관련 인자들의 관련성을 확 인하기 위하여 다양한 농도의 MEHF가 처리된 NCI-H460 세포를 대상으로 몇 가지 주요 죽음 수용체 연관 인자들 의 발현 변화 정도를 조사하였다. Fig. 4에 나타낸 결과에 서 알 수 있듯이 FasL 및 TRAIL과 같은 ligand의 발현이 MEHF 처리 농도의 증가에 따라 발현이 매우 증가하였 고, Fas 및 DR5와 같은 수용체의 발현도 증가하였다. 이 러한 결과는 MEHF에 의한 NCI-H460 세포의 apoptosis 유 도에는 죽음수용체 경로 활성화와 밀접한 관련이 있음 을 의미하는 것이다.
Fig. 4. Effects of MEHF on the levels of death receptor-related gene products in NCI-H460 cells. After incubation with MEHF for 48 h, the cells were lysed and the cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. The proteins were visualized using an ECL detection system. Actin was used as the internal control.
Fig. 3. Induction of DNA fragmentation and increased sub-G1 cell population by MEHF treatment in NCI-H460 cells. (A) Cells were harvested and centrifuged after treatment with the indicated concentrations of MEHF for 48 h. For the analysis of DNA fragmentation, genomic DNA was extracted and analyzed on 2% agarose gel containing EtBr. Marker indicates a size marker of the DNA ladder. (B) To quantify the degree of apoptosis induced by MEHF, the cells grown under the same conditions as (A) were evaluated for the sub-G1 DNA content using a flow cytometer.
4. MEHF가 미토콘드리아 경로에 미치는 영향
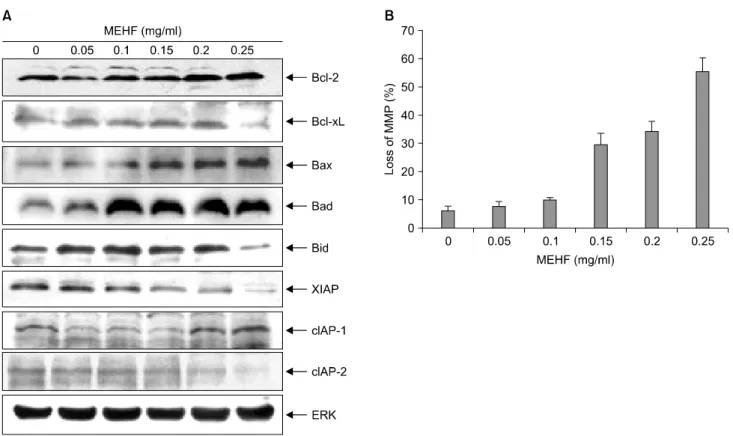
다음으로 미토콘드리아 경로, 즉 내인성 경로를 이루 는 단백질들의 발현에 미치는 MEHF의 영향을 조사하였 다. 미토콘드리아 경로는 성장인자, 호르몬, 산소 등의 결핍이나 방사선, 바이러스, free radicals 등의 자극으로 인해 유발되는 것으로, 이러한 자극이 미토콘드리아 막 전위(MMP)를 저하시켜 막 사이 공간에 존재하는 apoptosis 전구 단백질을 세포질로 방출시키게 된다.14,15) 이 때 방출된 cytochrome c가 Apaf-1 및 procaspase-9와 결합 하여 caspase-9를 활성화시켜 apoptosis를 유발하게 되는 데, 이 과정에는 inhibitors of apoptosis proteins (IAP) family 의 발현 저하 및 Bcl-2 family의 발현 변화가 밀접하게 연 관된다.16) Bcl-2 family는 미토콘드리아 막전위를 조절하 는 데 중요한 역할을 하며,21) Bcl-2와 Bcl-xL과 같은 anti-apoptotic 및 Bax와 Bid와 같은 pro-apoptotic 단백질의 발현 변화가 관여된다. 특히 Bid는 caspase-8이 활성화되 면서 tBid로 잘려 미토콘드리아 막의 투과성을 증가시키 는 등 죽음수용체 경로와 미토콘드리아 경로를 연결시 키는 핵심적인 역할을 한다.18) Fig. 5A에서 나타내었듯이 NCI-H460 세포에 MEHF를 48시간 처리한 결과 apoptosis 저해 단백질인 Bcl-xL의 발현은 감소되었고, apoptosis 전
구 단백질인 Bax 및 Bad의 발현은 증가하였으며, Bid가 발현이 감소되었음을 알 수 있었다. 또한 apoptosis를 저
Fig. 5. Effects of MEHF on the levels of mitochondrial pathway related-gene products and mitochondria membrane potential (MMP) in NCI-H460 cells. (A) After incubation with MEHF for 48 h, the cells were lysed and the cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. The proteins were visualized using an ECL detection system. Actin was used as the internal control. (B) To quantify the loss of MMP by MEHF, the cells grown under the same conditions as (A) were harvested and stained with JC-1 dye for 30 min, and MMP levels were measured by a flow cytometer.
해하는 IAP family인 xIAP 및 cIAP-2의 발현은 모두 감소 하였으며, 이와 함께 미토콘드리아 막전위(MMP) 수준도 MEHF 처리에 따라 농도 의존적으로 감소하였다(Fig.
5B). 이들 결과를 통해 MEHF가 NCI-H460 세포에 유도하 는 apoptosis는 미토콘드리아 경로를 거치며, 특히 Bid의 cleavage를 통해 죽음수용체 경로와 밀접하게 연관되어 있음을 유추할 수 있었다.
5. MEHF가 caspases 활성에 미치는 영향
이상의 결과에서 MEHF에 의한 NCI-H460 세포의 apoptosis 유발에는 죽음수용체 경로와 미토콘드리아 경 로 모두가 관련이 있을 것으로 나타났으므로, 이들 죽음 신호 경로에 관련된 caspase의 활성화 여부를 조사하였 다. 그 결과 caspase-8과 caspase-9의 비활성형의 발현이 MEHF 처리 농도의 증가에 따라 점차 감소하였으며, caspase-3 역시 MEHF 처리에 따라 비활성형의 발현이 현 저히 감소되었다(Fig. 6A). 이러한 단백질 수준에서의 결 과를 재확인하기 위하여 in vitro caspase 활성을 정량적으
로 조사한 결과, Fig. 6B에서 알 수 있듯이 MEHF의 처리 농도 증가에 따라 조사된 3종류의 caspase 모두가 처리 농도 의존적으로 활성이 점차 증가되었다. 즉 죽음수용 체 및 미토콘드리아 경로에 핵심적인 역할을 하는 caspase 의 활성이 모두 감소되어 MEHF에 의한 apoptosis 유발에 는 두 경로가 동시에 관여하고 있음을 알 수 있었다.
한편 활성화된 caspase는 세포의 정상적인 생존에 필수 적인 세포 내 주요 단백질들을 분해할 수 있으며, 단편화 가 일어난 이들 단백질들은 apoptosis가 유발되었다는 표 지자로서 활용이 된다. 본 연구에서는 대표적인 caspase-3 의 기질 단백질에 해당되는 CAD/ICAD 및 PARP의 발현 변화를 조사하였는데, ICAD는 caspase-3에 의해 절단되면 서 복합체를 이루고 있던 CAD를 자유롭게 분리시키고 그 결과 DNA 절단과 염색질 응축을 일으키는 역할을 한다.22) PARP는 정상세포의 DNA 수복이나 유전자 안정 성 유지에 중요한 역할을 하며, apoptosis 시에 caspase-3에 의해 분해가 일어나면 이러한 회복기능이 상실된다.23) Fig. 6A에 나타낸 것처럼 ICAD 및 PARP 단백질은 발현
Fig. 6. Activation of caspases and degradation of ICAD and PARP by MEHF treatment in NCI-H460 cells. (A) After incubation with MEHF for 48 h, the cells were lysed and the cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. The proteins were visualized using an ECL detection system. Actin was used as the internal control. (B) The cell lysates from the cells grown under the same conditions as (A) were assayed for in vitro caspase-3, caspase-8 and caspase-9 activity using DEVD-pNA, IETD-pNA, and LEHD-pNA, respectively, as substrates. The released fluorescent products were measured. The data represents the average±SD of three independent experiments.
이 MEHF 처리 농도의 증가에 따라 단편화의 정도가 증 가하였음을 알 수 있었는데, 이러한 결과는 MEHF 처리 에 따른 NCI-H460 세포의 apoptosis 유발은 caspase-3의 활 성을 통한 표적단백질의 단편화에 의하여 이루어지고 있음을 보여 주는 것이다.
결 론
본 연구에서는 항산화 및 항염증작용 등 다양한 약리 적 효능을 가진 것으로 보고되고 있는 톳의 항암작용에 관한 기전연구를 위하여 인체 폐암세포의 증식에 미치 는 톳 메탄올 추출물(MEHF)의 영향을 조사하였다.
MEHF가 처리된 폐암세포는 처리 농도 의존적으로 생존 률이 현저히 감소하였으며, 이러한 증식억제는 apoptosis 유도에 의한 것임을 전형적인 apoptosis가 유발된 세포에 서 관찰 가능한 세포막의 특이적인 blebbing 현상, apop- totic body 형성과 sub-G1기 세포의 증가, DNA의 단편화 등을 통하여 확인하였다. MEHF에 의한 apoptosis 유발 경 로 해석을 위한 다양한 실험의 결과, MEHF는 죽음의 수 용체 경로 활성을 통하여 caspase-8을 활성화시켰으며, Bid의 cleavage를 유발하여 미토콘드리아 경로가 활성화 되었음을 caspase-9의 활성 증가로 확인하였고, 이로 인해 caspase-3의 활성도 증가시켰으며, 증가된 caspase-3의 활
성에 의하여 caspase-3 표적 단백질들의 단편화가 유발되 었음을 알 수 있었다. 이상의 결과들은 인체 폐암세포에 서 톳의 항암작용을 이해하는 중요한 자료가 될 것이며, 향후 톳 메탄올 추출물의 어떤 성분이 apoptosis를 직접적 으로 일으키는지 여부와 그 자세한 기전 연구를 위한 기 초자료로서 활용될 것이다.
감사의 글
본 연구는 지식경제부·부산광역시 지원 지역혁신센터 사업 동의대학교 블루바이오 소재 개발 및 실용화 지원 센터(RIC08-06-07) 지원에 의하여 이루어진 결과임.
참 고 문 헌
1) Watanabe T, Hirayama T, Takahashi T, Kokubo T, Ikeda M.
Toxicological evaluation of arsenic in edible seaweed, Hizikia species. Toxicology 14, 1-22, 1979.
2) Yan X, Chuda Y, Suzuki M, Nagata T. Fucoxanthin as the major antioxidant in Hijikia fusiformis, a common edible seaweed. Biosci Biotechnol Biochem 63, 605-607, 1999.
3) Jung K, Ha E, Uhm Y, Park H, Kim MJ, Kim H, Baik H, Hong M, Yang J, Yim SV. Suppressive effect by Hizikia fusiforme on the production of tumor necrosis factor in BV2
murine microglial cells. Neurol Res 29, S88-92, 2007.
4) Kim SA, Woo MK, Kwak CS, Lee MS. Antimutagenic and cytotoxic effects of ethanol extracts from five kinds of seaweeds. J Korean Soc Food Sci Nutr 34, 451-459, 2005.
5) Ryu HS, Jung YH, Kim HS. Effect of Hizikia Fusiforme water extracts on mouse immune cell activation. J Korean Soc Food Sci Nutr 40, 624-629, 2007.
6) Ko MS, Shin KM, Lee MY. Effects of Hizikia fusiforme ethanol extract of antioxidative enzymes in ethanol-induced hepatotoxicity of rat liver. J Korean Soc Fooc Sic Nutr 31, 87-91, 2002.
7) Park KE, Jang MS, Lim CW, Kim YK, Seo YW, Park HY.
Antioxidant activity on ethanol extract from boiled-water of Hizikia fusiformis. J Korean Soc Appl Biol Chem 48, 435-439, 2005.
8) Shon JH, Kang DY, Oh HC, Jung BM, Kim MH, Shin MO, Bae SJ. The Effects on antimicrobial and cytotoxicity of Hijikia fusiformis fraction. J Korean Soc Food Sci Nutr 39, 444-450, 2006.
9) Kim TY, Jin CY, Kim GY, Choi IW, Jeong YK, Nam TJ, Kim SK, Choi YH. Ethyl alcohol extracts of Hizikia fusiforme sensitize AGS human gastric adenocarcinoma cells to tumor necrosis factor-related apoptosis-inducing ligand-mediated apoptosis. J Med Food 12, 782-787, 2009.
10) Kim SO, Choi YH. The ethyl alcohol extract of Hizikia fusiforme inhibits matrix metalloproteinase activity and regulates tight junction related protein expression in Hep3B human hepatocarcinoma cells. J Med Food 13, 31-38, 2010.
11) Norbury CJ, Hickson ID. Cellular responses to DNA damage.
Annu Rev Pharmacol Toxicol 41, 367-401, 2001.
12) Elmore A. Apoptosis: a review of programmed cell death.
Toxicol Pathol 35, 498-516, 2007.
13) Schulze-Osthoff K, Ferrari D, Los M, Wesselborg S, Peter ME. Apoptosis signaling by death receptors. Eur J Biochem 254, 439-459, 1998.
14) Saelens X, Festjens N, Vande Walle L, van Gurp M, van Loo G, Vandenabeele P. Toxic proteins released from mitochondria in cell death. Oncogene 23, 2861-2874, 2004.
15) Hill MM, Adrain C, Duriez PJ, Creagh EM, Martin SJ.
Analysis of the composition, assembly kinetics and activity of native Apaf-1 apoptosomes. EMBO J 23, 2134-2145, 2004.
16) Schimmer AD. Inhibition of apoptosis proteins: translating basic knowledge into clinical practice. Cancer Res 64, 7183- 7190, 2004.
17) Kim R. Recent advances in understanding the cell death pathways activated by anticancer therapy. Cancer 103, 1551- 1560, 2005.
18) Esposti MD. The roles of Bid. Apoptosis 7, 433-440, 2002.
19) Igney FH, Krammer PH. Death and anti-death: tumour resistance to apoptosis. Nat Rev Cancer 2, 277-288, 2002.
20) Kischkel FC, Hellbardt S, Behrmann I, Germer M, Pawlita M, Krammer PH, Peter ME. Cytotoxicity-dependent APO-1 (Fas/CD95)-associated proteins form a death-inducing signaling complex(DISC) with the receptor. EMBO J 14, 5579-5588, 1995.
21) Cory S, Adams JM. The Bcl-2 family: regulators of the cellular life-of-death switch. Nat Rev Cancer 2, 647-656, 2002.
22) Sakahira H, Enari M, Nahata S. Cleavage of CAD inhibitor in CAD activation and DNA degradation during apoptosis.
Nature 391, 96-99, 1998.
23) Kaufmann SH, Desnoyers S, Ottaviano Y, Davidson NE, Poirier GG. Specific proteolytic cleavage of poly (ADP-ribose) polymerase: an early marker of chemotherapy-induced apoptosis.
Cancer Res 53, 3976-3985, 1993.