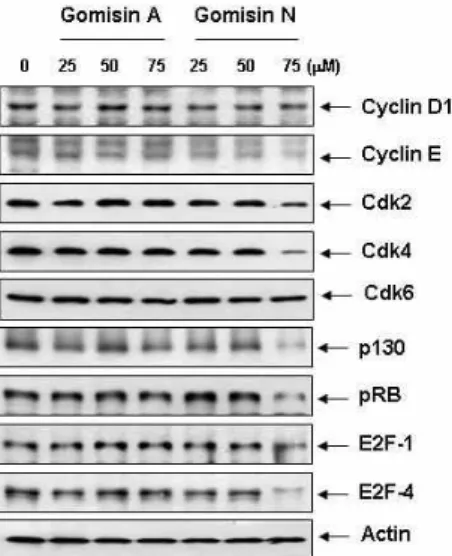
G1 Arrest of the Cell Cycle by Gomisin N, a Dibenzocyclooctadiene Lignan, Isolated from Schizandra chinensis Baill in Human Leukemia U937 Cells
6
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
수치


관련 문서
Caffeic acid phenethyl ester induces E2F-1-mediated growth inhibition and cell-cycle arrest in human cervical cancer cells.
All values were mean ± SD(n=3).. Gram staining of the isolated strains except Lb. plantarum EM from fermented fruit-vegetable juice using stored apple.. Changes in the
Concentration-Dependent inhibition of cell viability by adenosine in human oral fibroblast and FaDu human head and neck squamous cell carcinoma · · ·
It also suggest that cell proliferation inhibits by a novel signal transduction for adenosine effect in human FaDu hypopharynx squamous cell carcinoma cells....
In vitro cell migration in APE or JAG1 siRNA-treated M059K cell line Fig,11 Down-regulation of JAG1 induces S phase arrest in
Results: The human breast cancer cell subline MCF-7/MX5 cells selected in the presence of 5 µg/ml mitoxantrone (MX) were more resistant to MX (15.7... Western blot and
Particularly, after selecting a specific cycle by collecting HWC data and primary system-related data from Gori NPP Unit 1 and comparatively analyzing
tricuspidata on the production of proinflammatory cytokines in TNFα+IFNγ-stimulated HaCaT cells ...15 Fig.5: The cell viability of sub-fractions from 70% EtOH
