www.earticle.net
Adiponectin에 의한 유선상피세포의 글루코스 흡수 및 성장조절작용
여인서* ∙ 박병성 ∙ 고용균 ∙ 양부근 강원대학교 동물생명과학대학
Adiponectin Regulation of Glucose Uptake and Growth in Mammary Epithelial Cells
In-Suh Yuh*, Byung-Sung Park, Yong-Gyun Goh and Boo-Keun Yang College of Animal Life Sciences, Kangwon National University, Chuncheon 24341, Korea
ABSTRACT1)
The objective of the research was to identify the presence of adiponectin receptors and to study adiponectin action on glucose uptake and growth in mouse mammary epithelial cells. These cells expressed adiponectin receptors, AdipoR1 and AdipoR2. Insulin (10 ng/ml) or insulin like growth factor-I (IGF-I, 10 ng/ml) alone did not alter the degree of AdipoR1 and AdipoR2 genes expression from 0 to 4 h incubation.
Prolactin (10 ng/ml) or epidermal growth factor (EGF, 10 ng/ml) alone also did not induce the two genes’
mRNA in the incubation time. Adiponectin (1 μg/ml) alone or pre-incubation of insulin alone (100 ng/ml) for 2 h prior to adiponectin stimulation did not increase 2-deoxy-D-glucose,[1,2-3H] uptake but adiponectin+pre-incubation of insulin significantly increased glucose uptake compare to control (p<0.05). In a similar way, insulin alone or pre-incubation of adiponectin alone (2 h) did not increase glucose uptake but insulin+pre-incubation of adiponectin increased glucose uptake compare to control (p<0.05). Insulin sensitization for 2 h prior to adiponectin stimulation tended to increase glucose uptake response by the following adiponectin stimulation showing small interaction effect between insulin and adiponectin (p<0.1).
However, adiponectin sensitization for 2 hours prior to insulin stimulation did not shown interaction effect between adiponectin and insulin (p>0.1). The glucose uptake by both of hormones seems to be not interactive but additive (p<0.05). Adiponectin in the presence of 2% FBS decreased DNA synthesis of mammary epithelia (p<0.05). AICAR (100 or 200 μM), AMPK activator, decreased mammary epithelial cell growth in the presence of 2% FBS. These results indicate that adiponectin pathway has inhibitory effect on mammary epithelial cell growth.
(Key words: Adiponectin, AdipoR1, AdipoR2, Mammary, Glucose, Growth)
Ⅰ. INTRODUCTION
The substantial mammary growth occurs during postnatal periods. During the pregnancy, mammary ducts are branched
and elongated, and lobules-alveoli are formed and functionally differentiated to synthesize milk. The mammary glands finally undergo lactogenesis, galactopoiesis and involution. These whole biological processes are
* Corresponding author: In-Suh Yuh, Division of Applied Animal Science, Animal Biotechnology Program, Kangwon National University, Chuncheon 24341, Korea. Tel: +82-33-250-8626, E-mail: [email protected]
This is an Open Access journal distributed under the teams of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses(by-nc/3.0) which permits unrestricted non-commercial use, and reproduction in any medium, provided the original work is properly cited.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM
www.earticle.net
regulated by plasma hormones or local physiological regulators. Parenchymal and stromal interaction in mammary gland is necessary to potentiate hormone- induced biological action. The mammary fibrous stroma highly promoted tumorigenesis but adipose pad had relatively inhibitory effect to the fibrose stroma (Sakakura, 1981). The controversy might be arisen from types of stored fat, fatty acid composition and the differences of a variety of fat tissue secretion (Neville et al., 1998).
Fat tissue plays as an active role on regulating body metabolism by secreting various bioactive molecules termed adipokines (Wong et al., 2003; Kadowaki and Yamauchi, 2005). Adiponectin is an adipocyte-specific secreted protein and produced during adipocyte differentiation. It was known as adipocyte complement- related protein (acrp30), adipoQ (Scherer et al., 1995;
Maeda et al., 1996). Adiponectin is composed of an N-terminal collagen-like domain and a C-terminal globular domain with significant sequence and structural similarities to the complement factor C1q. Adiponectin inhibited gluconeogenesis and glycogenesis but increased fatty acids oxidation, glucose uptake and insulin sensitivity (Fruebis et al., 2001; Thomas et al., 2002; Wu et al., 2003). These effects were mediated through two distinct adiponectin receptors, AdipoR1 and AdipoR2 (Yamauchi et al., 2003). AdipoR1 abundantly produced in skeletal muscle, whereas AdipoR2 mostly expressed in the liver. These two distinct receptors are predicted to contain seven transmembrane domains, but functionally different from G-protein coupled receptors (Scheer et al., 1996; Wess, 1997).
PPARgamma agonists have been shown to increase adiponectin levels in mice (Yamauchi et al., 2001) and humans (Hiros et al., 2002) as well as in 3T3-L1 adipocytes in vitro(Yamauchi et al., 2001; Kadowaki et al., 2005). These effects are associated with adipocytes differentiation (Das et al., 2001) and direct transcriptional activation of genes via peroxisome proliferator response element (Combs et al., 2002; Bouskila et al., 2005). Both of adiponectin and PPARγ agonists have been shown to increase insulin sensitivity and ameliorate atherosclerosis (Bouskila et al., 2005).
Glucose is primary source for the synthesis of lactose which determines milk volume in mammary tissue by regulating osmorality in mammary gland. Therefore, glucose uptake in the mammary gland plays a major role in milk production (Bell et al., 1997). In addition, glucose is utilized for production of the chemical energies by catabolic pathway and the cofactor, NADPH, by pentose phosphate cycle for the synthesis of lactose, fat and milk proteins during lactation as well as for the synthesis of various macromolecules required for tissue remodeling and cell proliferation (Scott et al., 1976;
Threadogold et al., 1979). Mammary glands do not synthesize glucose from other metabolites by gluconeogenesis because of the deficiency of glucose–6 phosphatase activity in mammary tissue (Scott et al., 1976; Threadogold et al., 1979). Thus, glucose transport regulation through the plasma membrane of mammary epithelial cells provides a primary role in maintaining glucose homeostasis during mammary development and milking period.
The objective of this research is to identify the presence and types of adiponectin receptors and to examine adiponectin receptor regulation by some lactogenic hormones and growth factors. In addition, this study is to suggest the possible roles of adiponectin in glucose uptake and growth of normal mammary epithelial cells.
Ⅱ. MATERIALS and METHODS
1. Cell culture
Normal mouse mammary epithelial cells (NMuMG) were obtained from ATCC (Rockville, MD, USA).
Cultures were maintained at 37℃ in an atmosphere of 95% air/5% CO2. Log-phase cells were routinely cultured in sterile polystyrene T-75 and T-150 tissue culture flasks (Corning, New York, USA), containing Dulbecco’s Modified Eagle’s Medium (DMEM, Sigma-Aldrich, USA), supplemented with 10% fetal bovine serum (FBS, Gibco, USA), 50 U penicillin and 50 mg/ml streptomycin
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM
www.earticle.net
(Sigma-Aldrich).
2. DNA synthesis
Basal cell culture methods are same as described above cell culture methods. Mouse recombinant globular adiponectin (Alexis, San Diego, USA) was added at 0, 0.001, 0.01, 0.1 or 1 µg/ml concentration in the presence of 2% FBS after 24 hour serum starvation entering G0/G1. At 18 hour after treatment, 1 μCi/ml
3H-thymidine (50-90 Ci/mmol, DuPont) was added and incubated for 1 hour, and then rinsed three times with tris-buffered saline (TBS, 150 mM NaCl, 50 mM Tris, pH 7.5), three times with ice-cold 10% trichloroacetic acid (TCA), and three times with 100% ethanol. Cells were dried, dissolved in 0.5 M NaOH containing 0.1% triton X-100, neutralized with HCl, and counted by a scintillation counter.
3. Counting of cell numbers
NMuMG cells were plated in 24-well culture plate (105 cells/well) in 10% FBS for 24 h and then serum-starved for 24 h. Aminoimidazole carboxamide ribonucleotide (AICAR, Calbiochem, LaJolla, USA) was added at the indicated concentrations in the presence of 2% FBS supplementation in which cells were maintained with basal survivor for experimental days. Two days later from AICAR treatment, 2% FBS and AICAR treatment were refreshed and cell numbers were counted at 2 or 4 days later from treatment. Cell numbers were manually measured with hemocytometer.
4. Glucose uptake
Glucose transport was assayed on the base of uptake of 2-deoxy-D-glucose, [1, 2-3H]. Cell cultures for glucose uptake experiment were performed in the glucose free and phenol red free DMEM. Briefly, 3x105 NMuMG cells/well (24 well culture plate) were cultured in the presence of 10% FBS+DMEM for 24 h and then changed to serum free DMEM for 15 h. Half of treatment groups
were pre-incubated with insulin (100 ng/ml) and half of them were not for 2 hours and then culture media were changed to fresh DMEM. Adiponectin (1 μg/ml) and 2-deoxy-D-glucose, [1, 2-3H] (1 µCi/ml, 60 Ci/mmole), or 2-deoxy-D-glucose, [1, 2-3H] alone were added to each half of insulin pre-treatment groups for 30 min. In a similar ways, cells were treated by pre-incubation of adiponectin (1 μg/ml) in one group and not treated by adiponectin in the other group for 2 h. Cells were refreshed with DMEM and then added 2-deoxy-D-glucose, [1, 2-3H] (1 µCi/ml, 60 Ci/mmole) with or without insulin 100 ng/ml in both groups. Cells were washed with ice cold phosphate buffered saline (PBS, pH 7.4) 3 times and then air-dried. Cells were solubilized with 1 ml of 0.5 M NaOH-0.1% Triton X-100, neutralized with 1 N HCl, and then radioactivity was measured.
5. Total RNA isolation
Total RNAs were isolated using QIA shredder, RNeasy kit (Qiagen, Hilden, Germany), and RNA later RNA stabilization reagents(Ambion, Austin, TX). Upon isolation, total RNA was frozen at –80℃ until RT-PCR analysis or real time PCR.
6. RT-PCR
Two microgram of mRNA was reverse transcribed in a volume of 20 μl 50 mM tris-Cl (pH 8.3), 75 mM KCl, 5 mM MgCl2, 10 mM dithiothreitol, 0.5 mM each deoxy-NTP (dNTP), 0.5 μg random hexamer primer (Promega, Madison, WI), and 1 U avian myeloblastosis virus (AMV) reverse transcriptase (Promega). Reactions were performed at 37℃ for 1.5 h, 95℃ for 10 min, and quenched on ice immediately. PCR was performed in a DNA cycler (PTC 100, MJ Research, Watertown, MA) for 30 cycles with each cycle consisting of 30 seconds of denaturation at 95℃, 30 seconds of annealing at 60℃, and 30 seconds of extension at 72℃. Oligonucleotide primers (Integrated DNA technologies, Coralville, IA) were used to amplify adiponectin receptor cDNA. The
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM
www.earticle.net
sequences of primers of used were as follows: AdipoR1 (242 bp): sense, 5’-CTTGACGATGCTGAGACCAA-3’;
antisense, 5’-CGTGTGGGGAGCAGTAGAAG-3’, AdipoR2 (175 bp): sense, 5’-GCCCAGCTTAGAGACACCTG-3’;
antisense, 5’-GCCTTCCCACACCTTACAAA-3’, GAPDH (222 bp): sense, 5’-GATGACATCAAGAAGGTGGTGAA- 3’; antisense, 5’-GTCTTACTCCTTGGAGGCCATGT-3’.
7. Real time PCR
RNA samples were reverse transcribed as described above. Samples were then analyzed by real time PCR using a GeneAmp® 5700 system (PE Biosystems, Foster City, CA). cDNA 5 µl was diluted with 25 µl of SYBR Green®master mix (PE Biosystems) and amplified for 30 cycles using the AdipoR1 and AdipoR2 receptor primers described above (final concentration of 20 µM). As normalizing control, mouse glyceraldehydes-3-phosphate dehydrogenase (mGAPDH) primers (sequences were described at above RT-PCR method) were used. There action was carried in a final volume of 50 µl and for 40 cycles with each cycle consisting of 1 min denaturation at 95℃, 2 min of annealing at 56℃, and 1 min of extension at 72℃. Amplified PCR product was analyzed using GeneAMP®5700 software (PE Biosystems). At the end of the PCR run, a melting curve was performed to verify the presence of a single amplicon, and some samples were also analyzed by 5% polyacryl-amide gel electrophoresis (PAGE). Real time PCR data were analyzed by the ΔΔCT method (PE Biosystems GeneAmp Manual, Foster City, CA). Fold change detected in mRNA is calculated by 2-ΔΔCT, ΔΔ CT=(CTeffector:treated-CTGAPDH:treated)–(CTeffector:control– CTGAPDH:control), where subscripts refer to mRNA amplified (effector or GAPDH) and whether sample was treated or control.
8. Statistical analysis
Statistical analysis was performed using general linear
models procedure (SAS, 1985) and analysis of variance (ANOVA). Preplanned t-test was used to compare treatment groups. The adiponectin and insulin interaction effect of glucose uptake was estimated by orthogonal contrasts. Briefly, test for interaction effect between adiponectin and insulin (A): [(adiponectin+insulin) -insulin alone] vs [(adiponectin alone-no addition)]. Test for interaction effect of (B): [(insulin+adiponectin)- adiponectin alone] vs [(insulin alone–no addition)]. Unless otherwise stated significance was set to p<0.05.
Ⅲ. RESULTS AND DISCUSSION
1. Adiponectin receptors in mammary epithelia
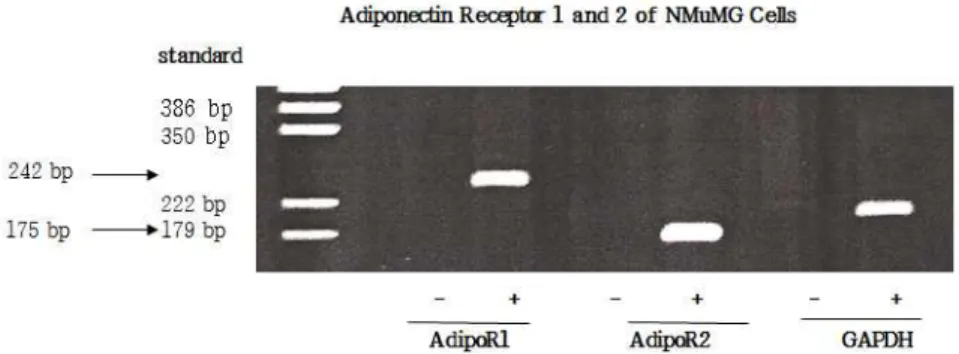
This research was to examine adiponectin and its receptor regulation as endocrine factor in glucose uptake and growth of mammary epithelial cells. To test the hypothesis that mammary epithelial cells express adiponectin receptors, total RNA was extracted and then RT-PCR reaction was performed for 30 cycles. In Fig. 1, the amplified-DNA fragments of AdipoR1 (242 bp), AdipoR2 (175 bp) and GAPDH (internal control) were detected in the presence of AMV-reverse transcriptase but were not in the absent of AMV-reverse transcriptase indicating no contamination of genomic DNA in total RNAs. Adiponectin increased the uptake of fatty acid and glucose, and modulated the metabolism of fatty acid and glucose, suggesting physiologically significant role in maintaining metabolic homeostasis for various tissue function (Yamauchi et al., 2003; Onay-Besikci et al., 2004). This result suggests the possibility that adiponectin receptors in mammary tissue would regulate mammary function such as milk synthesis as well as mammary growth by uptaking micro-nutrients for energy source and milk constituents. It should be clarified how the differential expressions of both type receptors alter differently mammary functions.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM
www.earticle.net
Fig. 1. RT-PCR analysis of adiponectin receptor 1 (AdipoR1) and receptor 2 (AdipoR2) expression. Total RNA was isolated from NMuMG cells grown 10% FBS and used as template for RT-PCR. The amplified DNA was fractioned by 5% polyacrylamide gel electrophoresis and visualized with ethidium bromide staining. +; RT-PCR reaction with avian myeloblastosis virus (AMV) reverse transcriptase, -; RT-PCR reaction without AMV reverse transcriptase. The product sizes of AdipoR1, AdipoR2 and GAPDH are 242 bp, 175 bp, and 192 bp, respectively.
2. Expression of adiponectin receptor genes by growth or/and lactogenic hormones
To test if the known lactogenic hormones (insulin or prolactin) or the known mammary tissue growth factors (insulin like growth factor-I, IGF-I or epidermal growth factor, EGF) regulate adiponectin receptor gene expression or not, real time PCR analysis was performed in Fig. 2. NMuMG cells were treated with lactogenic hormones or growth factors for 0, 30 min, 1, 2, and 4 h incubation after 24 h of G0/G1 phase. When cells were treated with insulin (10 ng/ml) or IGF-I (10 ng/ml), the magnitudes of gene expression of AdipoR1 or AdipoR2 were between 0.6 and 1.5 folds for all incubation times (30 min, 1, 2 and 4 h) to that of 0 incubation time and no statistical differences were shown when the effects of incubation times were compared with that of 0 incubation time (Fig. 2(A), (B)). These results were similar to those of the cultured in 1% FBS supplemented basal medium but insulin (10 ng/ml) addition showed a moderate decrease of AdipoR2 expression at 30 min after addition (Fig. 2(D), p<0.05). This decrease of AdipoR2 by insulin was quickly recovered after additional 30 min incubation. In our previous experiments, for long term incubation (12 hrs and 24 h) with insulin and IGF-I in the presence of 1% FBS, neither insulin nor IGF-I altered AdipoR1 and AdipoR1 gene expressions. It is not clear whether the short term decrease of AdipoR2 gene expression was arbitrary or not. Tsuchida et al. (2004)
reported that messenger RNA levels of AdipoR1 and AdipoR2 in the skeletal muscle and liver increased after fasting, and re-feeding quickly restored these to levels of original fed condition suggesting negative regulatory effect of insulin on adiponectin receptor gene expression in mRNA levels. However, the inhibitory effect of insulin on AdipoR2 expression in mammary epithelial cells of our experiment was temporal and rapidly recovered within 30 min from the decrease. Thus the inhibitory effect of insulin on AdipoR2 expression would not seem to be biologically meaningful. This result is compatible to the glucose uptake experiment in Fig. 3 (A), in which insulin does not down-regulate adiponectin effect on glucose uptake. In prolactin (10 ng/ml) or EGF (10 ng/ml) treatment, the relative magnitudes of gene expression for the various incubation times to that of 0 time were ranged between 0.7 and 1.4 in both types of adiponectin receptors and no statistical differences were shown to that of 0 time incubation (Fig. 2(E), (F)). In addition, when cells were incubated with 10 ng/ml of prolactin or EGF in the presence of 1% FBS, AdipoR1 and AdipoR2 gene expressions were not altered for all incubation times (Fig. 2(G), (H)). These results suggest that the known lactogenic hormones (insulin or prolactin) or mammary tissue growth factors (IGF-I or EGF) do not regulate adiponectin receptor gene expression for short time (maximum 4 h incubation) and long term incubation (maximum 24 h, in our previous experiment) in the
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM
www.earticle.net
Fig. 2. Effects of insulin, IGF-I, EGF and prolactin on the expression of AdipoR1 and AdipoR2 genes in NMuMG cells. Real time PCR results of AdipoR1 and R2 genes for the given time of incubation with effectors were normalized to GAPDH. The data described means (fold induction of gene expression to control)±pooled SEM (n=6) *, p<0.05 compared with the 0 time incubation (no addition). (A) and (B) AdipoR1 and AdipoR2 expression in the presence of insulin (10 ng/ml) or IGF-I (10 ng/ml) for the 0, 0.5, 1, 2 and 4 h incubation times, (C) and (D) AdipoR1 and AdipoR2 expression in the presence of insulin(10 ng/ml) or IGF-I (10 ng/ml) supplemented with 1% FBS (E) and (F) AdipoR1 and AdipoR2 expression in the presence of prolactin (10 ng/ml) or EGF (10 ng/ml), (G) and (H) AdipoR1 and AdipoR2 expression in the presence of prolactin (10 ng/ml) or EGF (10 ng/ml) supplemented with 1%
FBS.
presence or in the absence of 1% FBS. FBS 1%
supplementation to maintain better cell viability than without 1% FBS also did not alter lactogenic hormone
and growth factor response in adiponectin receptor gene expression.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM
www.earticle.net
3. Glucose uptake by adiponectin and insulin
In glucose uptake experiment, adiponectin (1 μg/ml) alone (Fig. 3(A)) or pre-incubation of insulin alone (100 ng/ml) for 2 h prior to adiponectin stimulation did not increase 2-deoxy-D-glucose, [1, 2-3H]] (1 µCi/ml, 60 Ci/mmole) uptake but adiponectin+pre-incubation of insulin significantly increased glucose uptake compare to control (p<0.05). In a similar way, insulin alone (Fig.
3(B)) or pre-incubation of adiponectin alone (2 h) did not increase glucose uptake but insulin+pre-incubation of adiponectin increased glucose uptake compare to control (p<0.05). To clarify if the pre-incubation of insulin or adiponectin sensitizes the other hormone effect on glucose uptake or not, interaction effect was analyzed by orthogonal contrast. When insulin sensitized cells for 2 h prior to adiponectin stimulation, glucose uptake response by adiponectin tended to increase to that of adiponectin without insulin pre-treatment (Fig. 3(A), p<0.1). This result cautiously suggests that if insulin sensitization for 2 h prior to adiponectin stimulation is effective to increase glucose uptake response by the following
adiponectin stimulation, this effect may not due to adiponecin receptor gene regulation but due to the activation of adiponectin signal intermediates by insulin.
Because insulin did not alter AdipoR1 and AdipoR2 gene expressions in Fig 2.(A), (B), (C), and (D). In addition, when adiponectin sensitized cells for 2 hours prior to insulin stimulation, there was no interaction effect between adiponectin and insulin in glucose uptake (Fig. 3(B), p>0.1). Over all results indicate that the effect of insulin and adiponectin on glucose uptake seemed to be additive rather than interactive suggesting independent effect in glucose uptake between two hormones. Ceddia et al. (2005) reported that insulin or adiponectin alone increased glucose transport and GLUT4 translocation in skeletal muscle cells. Tomas et al. (2002) reported that 2.5 μg/ml of globular form of adiponectin induced 1.5 fold increase in 2-deoxyglucose uptake for 30 min incubation via AMPK activation without insulin in muscle tissue. It could not be ruled out the possibility that low glucose uptake and less sensitization on adiponectin and insulin in our experiment were due to the stimulation by monomer
Fig. 3. Effect of adiponectin or/and insulin on uptake of 2-deoxy-D-glucose,[1,2,3H] in the glucose and phenol red free DMEM for 30 minutes incubation in NMuMG cells. (A) The pre-incubation of insulin (100 ng/ml) for 2 hours prior to adiponectin (1 µg/ml) stimulation. Half of treatment groups were pre-incubated with insulin (100 ng/ml) for 2 hours and the others were not, and then adiponectin (1 µg/ml) plus 2-deoxy-D-glucose,[1,2-3H] (1 µCi/ml, 60 Ci/mmole) and 2-deoxy-D-glucose, [1, 2-3H] alone were added into the insulin pre-incubation groups or no insulin pre-incubation groups for 30 minutes. (B) In similar way to (A), cells were inversely incubated by pre-incubation of adiponectin (1 µg/ml) for 2 hours, and then insulin stimulated for 30 min. Insulin (100 ng/ml) and 2-deoxy-D-glucose, [1, 2-3H] (1 µCi/ml, 60 Ci/mmole) or 2-deoxy-D-glucose, [1, 2-3H] alone were added for 30 minutes after 2 hour adiponectin pre-incubation group or no adiponectin pre-incubation group. *, p<0.05 compared with control (no addition).
Test for interaction effect between adiponectin and insulin (A): [(adiponectin+ insulin)-insulin alone] vs [(adiponectin alone-no addition)]. The interaction effect between adiponectin and insulin was shown at p<0.1. Test for interaction effect of (B):
[(insulin+adiponectin)-adiponectin alone]vs[(insulin alone–no addition)]. The interaction effect between adiponectin and insulin was not shown (p>0.1).
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM
www.earticle.net
type of adiponectin and low concentration of adiponectin.
Adiponectin was known to form a characteristic homo-multimers from trimers and hexamers to high molecular weight multimer (Richards et al., 2006).
Schraw et al. (2008) suggested that adiponectin complexes had distinct biochemical characteristics and biological activity. The high molecular weight (HMW) adeponectin complexes were significantly reduced in obese, insulin-resistance individuals than lean individuals.
Thus, the amount of HMW multimers could be more effective than total molecular weight to provide full biological activity in vivo or in vitro experiments.
4. Growth response by adiponectin
To examine the effect of adiponectin in mammary epithelial cell growth, adiponectin or AICAR which is an AMP-activated protein kinase (AMPK) activator and mimics adiponectin effect (Lemieux et al., 2003), was added into NMuMG cells which were grown in basal medium (2% FBS) and measured cell growth activity.
Addition of 1 μg/ml adiponectin significantly decreased DNA synthesis of NMuMG cells in basal medium (33,835 dpm/well in control vs 16,684 dpm/well in 1 μ g/ml adiponectin, p<0.05) (Fig. 4). In addition, the number of NMuMG cells grown in 2% FBS basal medium were significantly decreased in the presence of 100 and 200 µM of AICAR for 2 or 4 day incubation period (34.4 and 39.8% reduction to control for 2 day incubation in AICAR 100 or 200 µM or 38 and 45.5%
reduction to control for 4 day incubation in AICAR 100 or 200 µM, p<0.05) (Fig. 5). The significant inhibitory effect of AICAR in mammary epithelial growth was also shown in bovine transformed-mammary epithelial cells, MAC-T, in our previous experiment. In neoplasia tissue study, adiponectin inhibited tumor cell growth(Vona- Davis et al., 2007; Grossmann et al., 2008) and had inverse relationship between serum concentration of adiponectin and breast cancer incidence (Miyosi et al., 2003; Chen et al., 2006; Jarde et al., 2009). The adiponectin action is mediated by AMP-activated protein kinase (AMPK) which increases anti-proliferative and
apoptotic responses in human MCF-7 (Dieudonne et al., 2006; Grossmann et al., 2008; Otvos et al., 2011; Surmacz, 2013). These findings support that the growth inhibitory effect of adiponectin and AICAR in normal mammary epithelial cells could be mediated via AMPK pathways in similar ways to the neoplastic cells.
Fig. 4. Effect of adiponectin on DNA synthesis in NMuMG cells. Cells were serum starved for 24 h, adiponectin was added with 2% FBS, and DNA synthesis was measured 18 h later as3H-thymidine incorporated into acid insoluble material.
* p<0.05 compared with control (0 adiponectin in 2% FBS).
Fig. 5. Effect of AICAR (AMPK activator) on NMuMG cell growth. Cells were serum starved for 24 h, AICAR was added with 2% FBS for 2 days and cell number was counted. In four day culture experiment, AICAR treatments were refreshed with 2% FBS on the 3rd day and cell numbers were counted on the 5th day after treatment. For cell number counting, cells were detached with tyrpsin-EDTA (2.5 g/L trypsin with 0.001 M EDTA in Hank’s balance salt solution without Ca2+and Mg2+) solution for 10 min and cell numbers were counted with hemocytometer.
a p<0.05 compared with control of 2 day incubation.
b p<0.05 compared with control of 4 day incubation.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM
www.earticle.net
Ⅳ. CONCLUSION
Adiponectin receptors, AdipoR1 and AdipoR2, were expressed in the growing mammary epithelial cells.
Lactogenic hormones, insulin and prolactin, and growth factors, IGF-I and EGF did not alter AdipoR1 and AdipoR2 expression. Glucose uptake effects of insulin and adiponectin seem to be independent in mammary epithelial cells. Adiponectin is an inhibitory modulator in mammary epithelial cell growth.
ACKNOWLEDGEMENT
This research was supported by Department of Dairy Science, University of Wisconsin, Madison, WI 53706, USA and by 2014 research grant from Kangwon National University (no. 120140208), Chunchon Korea.
The authors thank to Dr. LG Sheffield and Dr. M Wiltbank at the University of Wisconsin-Madison, WI, USA.
Ⅴ. REFERENCES
1. Bell, A. W. and Bauman, D. E. 1977. Adaptation of glucose metabolism during pregnancy and lactation. J.
Mamm. Gland. Biol. Neopl. 2(3):265-278.
2. Bouskila, M., Pajvani, U. B. and Scherer, P. E. 2005.
Adiponectin: a relevant player in PPAR agonist-mediated improvements in hepatic insulin sensitivity? Inter. J. Obese Relat. Metab. Dis. 29 (Supple 1):S17-3.
3. Ceddia, R. B., Somwar, R., Maida, A., Fang, X., Bikopoulos, G. and Sweeney, G. 2005. Globular adiponectin increases GLUT4 translocation and glucose uptake but reduces glycogen synthesis in rat skeletal muscle cells. Diabetologia. 48:132-139.
4. Chen, D. C., Chung, Y. F., Yeh, Y. T., Chaung, H. C., Kuo, F. C., Fu, O. Y., Hou, M. F. and Yuan, S. S. 2006.
Serum adiponectin and leptin levels in Taiwanese
breast cancer patients. Cancer Letter. 237:109-114.
5. Combs, T. P., Wagner, J. A., Berger, J., Doebber, T., Wang, W. J., Zhang, B. B., Tanen, M., Berg, A. H., O’Rahilly, S., Savage, D. B., Chatterjee, K., Weiss, S., Larson, P. J., Gottesdiener, K. M., Gertz, B. J., Charron, M. J., Scherer, P. E. and Moller, D. E. 2002.
Induction of adipocyte complement-related protein of 30 kilodaltons by PPARγ agonists: a potential mechanism of insulin sensitization. Endocrinology.
143:998-1007.
6. Das, K., Lin, Y., Widen, E., Zhang, Y. and Scherer, P.
E. 2001. Chromosomal localization, expression pattern, and promoter analysis of the mouse gene encoding adipocytes-specific secretory protein Acrp30. Biochem.
Biophys. Res. Commun. 280:1120-1129.
7. Dieudonne, M. N., Bussiere, M., Santos, E., Leneveu, M. C., Giudicelli, Y. and Pecquery, R. 2006.
Adiponectin mediates antiproliferative and apoptotic responses in human MCF7 breast cancer cells.
Biochem. Biophys. Res. Commun. 345:271-279.
8. Fruebis, J., Tsao, T. S., Javorschi, S., Ebbets-Reed, D., Erickson, M. R., Yen, F. T., Buhain, B. E. and Losish, H. F. 2001. Proteolytic cleavage product of 30 kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc. Nat. Acad. Sci. USA. 98:2005-2010.
9. Grossmann, M. E., Nkhauta, K. J., Mizuno, N. K., Ray, A. and Cleary, M. P. 2008. Effects of adiponectin on breast cancer cell growth and signaling. British J.
Cancer. 98:370-379.
10. Hirose, H., Kawai, T., Yamamoto, Y., Taniyma, M., Tomita, M., Matsubara, K., Okazaki, Y., Ishii, T., Oguma, Y., Takei, I. and Saruta, T. 2002. Effects of pioglitazone on metabolic parameters, body fat distribution, and serum adiponectin levels in Japanese male patients with type 2 diabetes.
Metabolism. 51:314-317.
11. Jarde, T., Caldefie-Chezet, F., Goncalves-Mendes, N., Mishellany, F., Buechler, C., Penault-Llorca, F. and Vasson, M. P. 2009. Involvement of adiponectin and leptin in breast cancer: Clinical and in vitro studies.
Endo-Relat. Cancer. 16(4):1197-1210.
12. Kadowaki, T. and Yamauchi, T. 2005. Adiponectin
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM
www.earticle.net
and adiponectin receptors. Endo. Rev. 26(3):439-451.
13. Lemieux, K., Konrad, D., Klip, A. and Marette, A.
2003. The AMP-activated protein kinase activator AICAR does not induce GLUT4 translocation to transverse tubles but stimulates glucose uptake and mitogen-activated protein kinases alpha and beta in skeletal muscle. FASEB J. 17(2):1658-1665.
14. Maeda, K., Okubo, K., Shimomura, I., Funahashi, T., Matsuzawa, Y. and Matsubara, K. 1996. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (Adipose most abundant gene transcript 1). Biochem. Biophys. Res. Commun.
221:286-296.
15. Miyoshi, Y., Funahashi, T., Kihara, S., Taguchi, T., Tamaki, Y. M. and Hogchi, Y. S. 2003. Association of serum adiponectin levels with breast cancer risk.
Clin. Cancer Res. 15:5699-04.
16. Neville, M. C., Medina, D., Monks, J. and Hovey, R.
C. 1998. The mammary fat pad. J. Mamm. Gland.
Biol. and Neopl. 3:109-116.
17. Onay, B. A., Altarejos, J. Y. and Lopaschuk, G. D.
2004. gAd-globular head domain of adiponectin increases fatty acid oxidation in new born rabbit hearts. J. Biol. Chem. 279:44320-44326.
18. Otvos, Jr. L., Haspinger, E., Russa, F. L., Maspero, F., Graziano, P., Kovalszky, I., Lovas, K., Hoffmann, R., Knappe, D., Cassone, H., Wade, J. and Sumacz, E.
2011. Design and development of a peptide-based adiponectin receptor agonist for cancer treatment.
BMC Biotech. 11:90-105.
19. Richard, A. A., Stephens, T., Chariton, H. K., Jones, A., Macdonald, G. A., Prins, J. B. and Whitehead, J.
P. 2006. Adiponectin multimerization is dependent on conserved lysines in the collagenous domain:
evidence for regulation of multimerization by alterations in posttranslational modifications. Mol.
Endocrinol. 20(7):1673-1687.
20. Sakakura, T. 1981. Accelerated mammary cancer development by fetal salivary mesenchyme isografted to adult mouse mammary epithelium. J. Nation.
Cancer Insti. 66:953-959.
21. Scheer, A., Fanelli, F., Costa, T., DeBenedetti, P. G.
and Cotecchia, S. 1996. Constitutively active mutants of the alpha 1β-adrenergic receptor: role of highly conserved polar amino acids in receptor activation.
EMBO J. 15:3566-3578.
22. Scherer, P. E., Williams, S., Fogliano, M., Baldini, G.
and Lodish, H. F. 1995. A novel serum protein similar to C1q, produced exclusively in adipocytes. J.
Biol. Chem. 270:26746-26749.
23. Schraw, T., Wang, Z. V., Halberg, N., Hawkins, M.
and Scherer, P. E. 2008. Plasma adiponectin complexes have distinct biological characteristics.
Endocrinol. 149:2270-2282.
24. Scott, R. A., Bauman, D. E. and Clark, J. H. 1976.
Cellular gluconeogenesis by lactating bovine mammary tissue. J. Dairy Sci. 59:50-56.
25. Surmacz, E. 2013. Leptin and adiponectin: Emerging therapeutic targets in breast cancer. J. Mamm. Gland.
Biol. Neopl. 18:321-332.
26. Threadogold, L. C. and Kuhn, N. J. 1979. Glucose 6-phosphate hydrolysis by lactating rat mammary gland. Inter. J. Biochem. 10:683-685.
27. Tomas, E., Tsao, T. S., Saha, A. K., Murrey, H. E., Zhang, C. C., Itani, S. I., Lodish, H. F. and Ruderman, N. B. 2002. Enhanced muscle fat oxidation and glucose transport by ACRP30 globular domain: acetyl-CoA carboxylase inhibition and AMP-activated protein kinase activation. Proc. Nat.
Acad. Sci. USA. 99:16309-16313.
28. Tsuchida, A., Yamauchi, T., Ito, Y., Hada, Y., Maki, T., Takekawa, S., Kamon, J., Kobayashi, M., Suzuki, R., Hara, K., Kubo, N., Terauchi, Y., Froguel, P., Nakae, J., Kasuga, M., Accili, D., Tobe, K., Ueki, K., Hagai, R. and Kadowaki, J. 2004. Insulin/Foxo1 pathway regulates expression levels of adiponectin recepors and adiponectin sensitivity. J. Biol. Chem.
279(29):30817-30822.
29. Vona-Davis, L. and Rosa, D. P. 2007. Adipokines as endocrine, paracrine, and autocrine factors in breast cancer risk and progression. Endocrin-relat. Cancer.
14(2):189-206.
30. Wess, J. 1997. G-protein-coupled receptors: molecular mechanisms involved in receptor activation and
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM
www.earticle.net
selectivity of G-protein recognition. FASEB J.
11:346-354.
31. Wong, G. W., Wang, J., Hug, C., Tsao, T. S. and Lodish, H. F. 2003. A family of Acrip30/ adiponectin structural and functional paralogs. Proc. Nat. Acad.
Sci. USA. 101(28):10303-10307.
32. Wu, X., Motoshima, H., Mahadev, K., Stalker, T. J., Scalia, R. and Goldstein, B. J. 2003. Involvement of AMP-activated protein kinase in glucose uptake stimulated by the globular domain of adiponectin in primary rat adipocytes. Diabetes. 52:1355-1363.
33. Yamauci, T., Kamon, J., Ito, Y., Tsuchida, A., Yokomizo, T., Kita, Sugiyama, S., Miyagishi, T. M., Hara, K., Tsunoda, M., Murakami, M., Ohteki, T., Uchida, S., Takekawa, S., Waki, H., Tsuno, N. H., Shibata, Y., Terauchi, Y., Froguel, P., Tobe, K., Koyasu, S., Taira, K., Kitamura, T., Shimizu, T.,
Nagai, R. and Kadowaki T. 2003. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature. 423:762-69.
34. Yamauchi, T., Kamon, J., Waki, H., Terauchi, Y., Kubota, N., Hara, K., Mori, Y., Ide, T., Murakami, K., Tsuboyama-Kasaoka, N., Ezaki, O., Akanuma, Y., Gavrilova, O., Vinson, C., Reitman, M. L., Kagechika, H., Shudo, K., Yoda, M., Nakano, Y., Tobe, K., Nagai, R., Kimura, S., Tomita, M., Froguel, P. and Kadowakiet, T. 2001. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med.
7:941-946.
(Received 03 April 2015, Revised 11 November 2015, Accepted 12 November 2015)
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:56 PM

![Fig. 3. Effect of adiponectin or/and insulin on uptake of 2-deoxy-D-glucose,[1,2, 3 H] in the glucose and phenol red free DMEM for 30 minutes incubation in NMuMG cells](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5176823.595073/7.892.235.657.673.899/effect-adiponectin-insulin-uptake-glucose-glucose-minutes-incubation.webp)

