대한소화기학회지 2004;44:53-56 □ EDITORIAL □
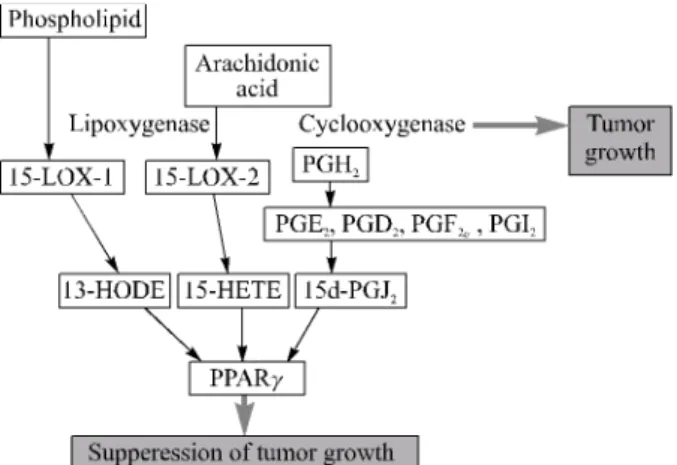
1)COX-2 (cyclooxygenase-2)를 과발현시킨 형질전환 생쥐 에서 암 발생이 증가하고 위암을 비롯한 각종 인체 암에서 COX-2의 발현이 증가한다. 또한 COX-2의 활성을 억제하는 비스테로이드성 소염제(nonsteroid anti-inflammatory drug, NSAID) 및 COX-2를 목표로 하는 약물의 투여는 대장암, 대장용종, 위암 및 다양한 종양의 발생을 현저하게 억제하 여 화학적 암 예방 효과를 보인다. 이러한 사실들은 COX-2 가 위암의 발생에 직접적으로 관여하고 있음을 시사한다.1-6 COX는 지질 대사에 중요한 단백질로서 phospholipase A2 (PLA2)에 의해 생성된 arachidonic acid (AA)를 prosta- glandin H2 (PGH2)로 전환하는 효소이다. 이러한 PGH2는 PGE2, PGF2α, PGI2, PGD2, 그리고 TXA2로 대사되어 염증 반응 및 다양한 세포 생리 작용에서 중요한 매개인자 역할 을 한다(Fig. 1).7-10 COX 단백질은 두 종류의 이종형(iso- form)을 가지는데, 그 중 COX-1은 대부분의 세포에서 항상 발현되는 house-keeping protein이며 COX-2는 세포의 비자 극 상태에서는 극히 소량이 발현되지만 호르몬, 친염증 사 이토카인, 세포성장 인자, 암 촉진자(tumor promoter) 등의 세포 자극에 의해 발현이 급격히 증가되는 단백질이다. 따 라서 COX-2는 PG 합성에 있어 중요한 속도 조절 효소(rate limiting enzyme)이다.11-13
COX-2는 대장암, 식도암, 및 위암을 비롯한 여러 종류의 소화기 암에서 과발현되며,14-16 세포사멸의 감소, 세포의 과 증식, 신생혈관의 발달, 림프절 전이 등과 밀접한 상관 관
연락처: 함기백, 443-721, 경기도 수원시 팔달구 원천동 산5 아주대학교병원 간 및 소화기유전체센터
Tel: (031) 219-4381, Fax: (031) 219-4399 E-mail: [email protected]
계가 있다.17-20 특히 이러한 COX-2 발현은 위암의 진행 단 계와 관계를 가지고 있어 정상 위점막이나 만성 위염 또는 만성 위축성 위염에서는 발현되지 않지만 장상피화생에서 발현이 증가하여 이형성의 44%에서, 장상피형 위선암에서 58%로 발현이 증가된다.21-23 그리고 이러한 COX-2의 과발 현은 위암의 예후와 밀접한 관련이 있다.24,25 2)
COX-2에 의해서 생성된 PG는 세포의 분열, 혈류의 조 절, 면역반응의 매개, 골격계의 발달, 상처의 치유 등 많은 세포의 정상 생리에서 중요한 조절자로서 작용한다. 뿐만 아니라 이러한 PG는 염증반응이나 암화와 같은 질병의 발 생 기전과도 관계가 있다. 특히 COX-2에 의해 생성된 PG 중 PGD2는 PGJ2, Δ12-PGJ2를 거쳐 15d-PGJ2를 형성한다 (Fig. 1). 이러한 15d-PGJ2는 peroxisome proliferator-activated receptor (PPAR)를 활성화시키는 배위자(ligand)로 잘 알려 져 있다.
PPAR은 핵막에 있는 호르몬 receptor family 중 하나로 PPARα, PPARβ/δ, PPARγ의 이종형를 가지며 지질의 대사 와 지방세포의 분화에 관여한다.26-28 15d-PGJ2는 PPAR 중 PPARγ의 배위자로 가장 잘 알려져 있으며 PPARγ는 유방 암, 전립선암, 폐암, 위암, 대장암, 췌장암을 비롯한 다양한 인체 암에서 발현된다.29,30 15d-PGJ2를 비롯한 PPARγ의 배 위자는 세포증식의 억제 및 세포사멸을 촉진함으로써 종양 세포의 성장을 억제할 수 있는 물질로 주목받고 있다. 하지 만 최근 들어 저농도의 PG에 의해서는 암세포의 성장이
Correspondence to: Ki-Baik Hahm, M.D.
San 5 Wonchon-dong, Paldal-gu, Suwon 443-721, Korea Department of Gastroenterology, Ajou Univestity School of Medicine
Tel: +82-31-219-4381, Fax: +82-31-219-4399 E-mail: [email protected]
위암의 발생에서 COX-2 및 PPAR의 역할
아주대학병원 소화기내과·간 및 소화기유전체센터
여말희·함기백
Role of COX-2 and PPAR in Gastric Carcinogenesis
Marie Yeo, Ph.D. and Ki-Baik Hahm, M.D.Genome Research Center for Gastroenterology, Ajou University School of Medicine, Suwon, Korea
대한소화기학회지: 제44권 제1호, 2004
Fig. 1. Activation of PPARγ by ligands produced by lipid meta- bolism. Three metabolites synthesized by COX and LOX enzyme potentially activate PPARγ. 15d-PGJ2 among these PPARγligands is formed by the cyclooxygenase pathway. COX-2 is the enzyme that catalyzes the first steps in the biosynthesis of the prostaglandins (PGs) from the substrate arachidonic acid (AA). The PGH2 is transformed into PGE2, PGF2α, PGD2, and PGI2. Other two ligand molecules, 13-HODE and 15-HETE, are metabolites produced by the action of lipoxygenase.
Fig. 2. A proposed mechanism of PPARγsignaling. PPARγforms complex of a heterodimer with RXR which is tightly associated with corepressor (R) in resting status. Following binding with ligand such as 15d-PGJ2, thiazolidinedione and NSAID, the corepressor is replaced with coactivator (A) and ligand-receptor complex binds at PPRE motif of relevant target gene promoter (eg.
PTEN). Genes induced by PPARγ lead to cell growth arrest or apoptosis via inhibition of PI3K signaling. This schematic figure was modified from Na HK, et al.35
오히려 유도된다는 상반적인 보고도 있다. 따라서 이러한 PPARγ는 세포내 배위자의 농도에 의존적으로 세포의 사멸 을 억제 또는 촉진하는 상반된 역할을 한다.31-34 PPARγ는 세포 내에서 retinoid X receptor (RXR)와 같이 peroxisome proliferator response element (PPRE)에 결합하여 phos- phatase and tensin homolog deleted on chromosome 10 (PTEN)의 발현을 증가시키며, 이러한 PTEN은 AKT 활성 억제를 통해 세포사멸을 유도하게 된다(Fig. 2).35
최근 장 등36은 ‘위암에서 cyclooxygenase-2 (COX-2)와 PPAR 유전자의 역할’이라는 논문에서 COX-2와 이의 대사 산물인 PGE2가 위암 환자의 위암 조직에서 주위 비위암 조 직에 비해 현저하게 과발현됨을 보고하였고, 이에 반해 PPARδ, PPARγ와 그의 배위자(ligand)인 15d-PGJ2의 생성 은 통계적으로 유의한 상관관계가 없음을 보고하였다. 이 러한 보고는 위암의 생성에 있어서 COX-2와 그의 대사 산 물인 PGE2의 생성이 결정적으로 기여함을 시사한다. 이들 은 15명의 조기 위암 환자와 30명의 진행성 위암 환자, 그 리고 대조군으로 정상인 30명의 내시경 생검으로부터 채 취한 조직을 사용하여 real time PCR 및 enzyme-linked immunosorbent assay (ELISA) 방법을 사용하여 COX-2, PPARδ, PPARγ와 PGE2, 15d-PGJ2를 각각 측정하였다.
Chang 등의 연구 결과로부터 COX-2는 정상인(3.46±1.72) 에 비해 조기 위암 환자와 진행성 위암 환자에서 8.32±
4.84와 8.16±2.67로 현저하게 발현이 증가되어 있으며 이 의 대사 산물인 PGE2 역시 조기 위암 환자(5.31±0.49)와 진행성 위암환자(5.46±0.54)에서 정상인(4.22±0.80)에 비 해 통계적으로 유의하게 증가하였다. 이에 반해 PPARδ와 PPARγ는 정상인(125.1±92.5와 0.78±0.61)에 비해 조기 위 암 환자와 진행성 위암 환자에서 98.7±108.9와 0.57±0.47, 그리고 102.0±76.3와 0.60±0.43로 다소 감소하였다. 그리 고 이의 배위자인 15d-PGJ2도 역시 조기 위암 환자(197.7±
102.6)와 진행성 위암 환자(211.0±76.9)에서 정상인(224.9
±89.2)에 비해 감소하였다. 하지만 이들 유전자 발현은 암 의 침윤 정도, 림프절 전이 및 원격 전이 등의 TNM 병기와 유의한 관계가 없었으며, 조직학적으로 장형이나 미만형 분류와도 상관관계가 없었다. 또한 이들은 H. pylori 감염 여부와의 상관관계를 조사하였고 COX-2의 발현에 있어서 H. pylori 감염 음성군(5.1±7.02)에 비해 H. pylori 감염 양 성군에서 7.0±3.75로 증가하였다. 이는 위암 발생에 있어 COX-2와 이 효소에 의해 생성된 대사 산물의 중요성에 대 해 다시 상기시켜 주며, 따라서 이 효소의 활성을 억제함으 로써 위암 발생 예방에 기여할 수 있는 유용한 토대를 제시 한다.
하지만 COX-2의 발현과 위암의 병기 및 조직 소견과의 상관관계을 확인하지 못하였고 이에 대한 폭넓은 이해를 필요로 한다. 왜냐하면 이미 여러 역학적 연구 결과에 의해 COX-2의 발현은 위암의 진행 정도와 밀접한 상관관계를 가지며 특히 전암 병변인 장상피화생 및 위선암에서 높게 발현된다. 또한 암의 침윤 정도와 전이 유무와도 관련성이 높으며 미만형 위암보다 장형 위암에서 발현 정도가 높
다.17-23 이러한 결과는 전암 병소에서부터 COX-2의 발현이
증가되고 증가된 COX-2와 이 효소에 의한 대사 산물이 위 암 발생 및 진전에 결정적인 기여를 한다고 추측할 수 있으 54
여말희 외 1인. 위암의 발생에서 COX-2 및 PPAR의 역할
며, 따라서 이 효소의 활성을 억제함으로써 위암 발생을 감 소시킬 수 있는 근거가 된다.
또한 위암의 유무와 상관없이 H. pylori 비감염군에 비해 H. pylori 감염군에서 COX-2의 발현이 증가되었지만 이의 대사 산물인 PGE2의 유의한 증가는 확인되지 않았다. 역시 위암 조직에서 증가된 COX-2나 PGE2는 이의 대사 산물 중 하나인 15d-PGJ2의 증가를 유도하지 않았으며 PPAR의 발 현 차이를 확인하지 못하였다. 따라서 COX-2 및 PPAR의 발현 정도의 비교는 물론 조금 더 나가 COX-2 증가가 암 발생에 미치는 분자생물학적 기전 및 PPAR 발현 간의 구 체적인 상관관계에 대한 분석이 요구된다. COX-2의 발현 이 PPAR에 의하여 되먹이 방식으로 조절될 수 있기 때문 에37 이를 고려하여 분석하게 되면 좀더 구체적인 발암 기 전의 역할을 규명할 수 있으리라 기대된다.
이러한 장 등36의 보고는 지질의 대사에 의한 세포 신호 전달체계의 조절이 정상 세포 생리뿐 아니라 암 발생의 병 인 기전에서 얼마나 괄목할 역할을 하는지 제시하고 있으 며, 이러한 신호전달체계는 COX뿐만 아니라 여러 인자에 의해 보다 복잡하고 정교하게 조절되고 있음을 암시한다.
Linoleic acid는 phospholipase에 의해 AA를 생산하고 이러 한 AA는 COX에 의해 여러 종류의 PG들을 형성한다. 한편 으로는 15-LOX-1과 15-LOX-2에 의해 15-HETE와 13- HODE이 형성된다. COX에 의해 생성된 15d-PGJ2와 LOX 에 의한 15-HETE와 13-HODE는 강력한 PPARγ의 배위자 이다(Fig. 1).38,39 이러한 COX와 LOX에 의해 생성된 배위
자는 PPARγ와 RXR의 heterodimer 복합체에 결합하여 전 사 억제자(repressor)을 해리하고 coactivator와 결합함으로 써 PPRE의 element에 결합하여 세포의 자극에 적절한 유전 자의 전사를 활성화시킨다(Fig. 2). 이러한 PPAR의 활성화는 종양세포에서 세포성장을 억제하거나 세포사멸, 세포의 분 화을 유도함으로써 암 형성을 억제한다. 활성화된 PPAR 신 호전달체계는 역으로 COX-2의 활성화를 상호적(reciprocally) 으로 MAPK와 NF-κB 활성을 억제를 통해 조절한다(Fig.
3).40
따라서 위암에서 COX-2 및 PPAR의 유전자 발현을 조 사한 장 등의 논문은, 이들 유전자에 의한 지질 대사의 신 호전달이 위암 형성에 미치는 영향과 이들 유전자를 이용 한 위암 치료에 대한 이해와 정확한 기전 연구에 대한 필요 성을 제시해 준 연구 결과라 하겠다.
참고문헌
1. Warner TD, Mitchell JA. Cyclooxygenase: new forms, new inhibitors, and lessons from the clinic. FASEB J 2004;18:
790-804.
2. Wang WH, Huang JQ, Zheng GF, Lam SK, Karlberg J, Wong BC. Non-steroidal anti-inflammatory drug use and the risk of gastric cancer: a systematic review and meta-analysis.
J Natl Cancer Inst 2003;3:1784-1791.
3. Eberhart CE, Dubois RN. Eicosanoids and the gastrointestinal tract. Gastroenterology 1995;109:285-301.
4. Sjodahl R. Nonsteroidal anti-inflammatory drugs and the gastrointestinal tract. Extent, mode, and dose dependence of anticancer effects. Am J Med 2001;110:66S-69S.
5. Jiang XH, Wong BC. Cyclooxygenase-2 inhibition and gastric cancer. Curr Pharm Des 2003;9:2281-2288.
6. Saukkonen K, Rintahaka J, Sivula A, et al. Cyclooxygenase-2 and gastric carcinogenesis. APMIS 2003;111:915-925.
7. Dannenberg AJ, Subbaramaiah K. Targeting cyclooxygenase-2 in human neoplasia: rationale and promise. Cancer Cell 2003;
4:431-436.
8. Helliwell RJ, Adams LF, Mitchell MD. Prostaglandin synthases: recent developments and a novel hypothesis.
Prostaglandins Leukot Essent Fatty Acids 2004;70:101-113.
9. Murakami M, Kudo I. Recent advances in molecular biology and physiology of the prostaglandin E2-biosynthetic pathway.
Prog Lipid Res 2004;43:3-35.
10. Murakami M, Nakatani Y, Kuwata H, Kudo I. Cellular components that functionally interact with signaling phospholipase A(2)s. Biochim Biophys Acta 2000;1488:
159-166.
11. Turini ME, DuBois RN. Cyclooxygenase-2: a therapeutic Fig. 3. Feed-back control of COX-2 expression by PPARγ. COX-2
produces several prostaglandins (PGs) such as PGD2 or PGE2. PGD2 formed by COX-2 converts 15d-PGJ2 via PGJ2, and the biosynthesized 15d-PGJ2binds PPARγreceptor complex. Activated PPARγ signaling with conjugation of ligand controls reciprocally COX-2 transcription through interfering with NF-κB pathway and MAPK pathway.
55
대한소화기학회지: 제44권 제1호, 2004
target. Annu Rev Med 2002;53:35-57.
12. van der Donk WA, Tsai AL, Kulmacz RJ. The cyclo- oxygenase reaction mechanism. Biochemistry 2002;41:15451- 15458.
13. Claria J. Cyclooxygenase-2 biology. Curr Pharm Des 2003;
9:2177-2190.
14. Stoehlmacher J, Lenz HJ. Cyclooxygenase-2 inhibitors in colorectal cancer. Semin Oncol 2003;30:10-16.
15. Altorki N. COX-2: a target for prevention and treatment of esophageal cancer. J Surg Res 2004;117:114-120.
16. Sinicrope FA, Gill S. Role of cyclooxygenase-2 in colorectal cancer. Cancer Metastasis Rev 2004;23:63-75.
17. van der Woude CJ, Kleibeuker JH, Jansen PL, Moshage H.
Chronic inflammation, apoptosis and (pre-)malignant lesions in the gastro-intestinal tract. Apoptosis 2004;9:123-130.
18. O'Byrne KJ, Dalgleish AG, Browning MJ, Steward WP, Harris AL. The relationship between angiogenesis and the immune response in carcinogenesis and the progression of malignant disease. Eur J Cancer 2000;36:151-169.
19. Williams CS, Tsujii M, Reese J, Dey SK, DuBois RN. Host cyclooxygenase-2 modulates carcinoma growth. J Clin Invest 2000;105:1589-1594.
20. Leahy KM, Koki AT, Masferrer JL. Role of cyclooxygenases in angiogenesis. Curr Med Chem 2000;7:1163-1170.
21. Saukkonen K, Nieminen O, van Rees B, et al. Expression of cyclooxygenase-2 in dysplasia of the stomach and in intestinal-type gastric adenocarcinoma. Clin Cancer Res 2001;7:1923-1931.
22. Joo YE, Oh WT, Rew JS, Park CS, Choi SK, Kim SJ.
Cyclooxygenase-2 expression is associated with well- differentiated and intestinal-type pathways in gastric carcino- genesis. Digestion 2002;66:222-229.
23. van Rees BP, Saukkonen K, Ristimaki A, et al. Cyclo- oxygenase-2 expression during carcinogenesis in the human stomach. J Pathol 2002;196:171-179.
24. Tatsuguchi A, Matsui K, Shinji Y, et al. Cyclooxygenase-2 expression correlates with angiogenesis and apoptosis in gastric cancer tissue. Hum Pathol 2004;35:488-495.
25. Buskens CJ, Van Rees BP, Sivula A, et al. Prognostic significance of elevated cyclooxygenase 2 expression in patients with adenocarcinoma of the esophagus. Gastro- enterology 2002;122:1800-1807.
26. Rosen ED, Spiegelman BM. PPARgamma: a nuclear regulator of metabolism, differentiation, and cell growth. J Biol Chem 2001;276:37731-37734.
27. Yu K, Bayona W, Kallen CB, et al. Differential activation of
peroxisome proliferator-activated receptors by eicosanoids. J Biol Chem 1995;270:23975-23983.
28. Tontonoz P, Hu E, Spiegelman BM. Stimulation of adipogenesis in fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell 1994;79:1147-1156.
29. Koeffler HP. Peroxisome proliferator-activated receptor gamma and cancers. Clin Cancer Res 2003;9:1-9.
30. Fajas L, Debril MB, Auwerx J. Peroxisome proliferator- activated receptor-gamma: from adipogenesis to carcino- genesis. J Mol Endocrinol 2001;27:1-9.
31. Theocharisa S, Margeli A, Kouraklis G. Peroxisome pro- liferator activated receptor-gamma ligands as potent anti- neoplastic agents. Curr Med Chem Anti-Canc Agents 2003;3:
239-251.
32. Murphy GJ, Holder JC. PPAR-gamma agonists: therapeutic role in diabetes, inflammation and cancer. Trends Pharmacol Sci 2000;21:469-474.
33. Krzystyniak KL. Current strategies for anticancer chemo- prevention and chemoprotection. Acta Pol Pharm 2002;
59:473-478.
34. Kliewer S, Umesono K, Noonan PJ, Heyman RA, Evans RM.
Convergence of 9-cis retinoic acid and peroxisome pro- liferator signaling pathways through heterodimer formation of their receptors. Nature 1992;358:771-774.
35. Na HK, Surh YJ. Peroxisome proliferator activated receptor r (PPAR ) ligands as bifunctimal regulators of cell proliferation. Biochem Pharmacol 2003;66:1381-1391.
36. Chang YW, Cho HL, Jang JY, et al. Role of cyclo- oxygenase-2 (COX-2) and peroxisome proliferator-activated receptor (PPAR) in gastric cancer. Korean J Gastroenterol 2004;43:291-298.
37. Inoue H, Tanabe T, Umerson K. Feedback control of cyclooxgenase-2 expression through PPARr. J Biol Chem 2000;275:28028-28032.
38. Tontonoz P, Nagy L, Alvarez JG, Thomazy VA, Evans RM.
PPARgamma promotes monocyte/macrophage differentiation and uptake of oxidized LDL. Cell 1998;93:241-252.
39. Pham H, Banerjee T, Nalbandian GM, Ziboh VA. Activation of peroxisome proliferator-activated receptor (PPAR)-gamma by 15S-hydroxyeicosatrienoic acid parallels growth suppres- sion of androgen-dependent prostatic adenocarcinoma cells.
Cancer Lett 2003;189:17-25.
40. Hu E, Kim JB, Sarraf P, Spiegelman BM. Inhibition of adipogenesis through MAP kinase-mediated phosphorylation of PPARgamma. Science 1996;274:2100-2103.
56