12 책임저자:권택규, 700-712, 대구시 중구 동산동 194번지
계명대학교 의과대학 면역학교실 Tel: 053-250-7846, Fax: 053-250-7074 E-mail: [email protected]
접수일:2008년 3월 3일, 게재승인일:2008년 3월 18일
Correspondence to:Taeg Kyu Kwon
Department of Immunology, School of Medicine, Keimyung University, 194, Dongsan-dong, Jung-gu, Daegu 700-712, Korea
Tel: +82-53-250-7846, Fax: +82-53-250-7074 E-mail: [email protected]
암세포에서 NAG-1의 발현 및 기능
계명대학교 의과대학 면역학교실 임준희ㆍ권택규
Expression and Functional Role of NAG-1 in Cancer
Jun Hee Lim and Taeg Kyu Kwon
Department of Immunology, School of Medicine, Keimyung University, Daegu 700-712, Korea
Nonsteroidal anti-inflammatory drug (NSAID)-activated gene-1 (NAG-1) was identified from indomethacin (a cyclooxygenase (COX) inhibitor)-induced gene library. NAG-1 is a member of TGF-β superfamily and is also known as macrophage inhibitory cytokine-1 (MIC-1), placental transforming growth factor β (PTGF-β), or prostate derived factor (PDF). Purified recombinant NAG-1 is able to inhibit lipopolysaccharide-induced TNF-α production in macrophages, suggesting that NAG-1 acts as an autocrine regulatory molecule. In vitro and in vivo NAG-1 has anti-tumorigenic and pro-apoptotic activities independent of COX inhibition. Not only NSAIDs but also several anti-tumorigenic compounds with chemopreventive activities, including resveratrol, genistein, catechins, and peroxisome proliferators- activated receptor-γ ligands, regulate NAG-1 expression in a prostaglandin-independent manner. Thus, pro-apoptotic activity of NAG-1 may provide a molecular basis to explain chemopreventive agents- mediated anti-tumorigenesis. However, the biological activity of NAG-1 is very poorly characterized, and reports in the literature show high expression in tumors. Thus, understanding the regulatory mechanism of NAG-1 by chemotherapeutic and chemopreventive agents and their implications in cancer treatments may provide a rational approach for using such as tumor therapeutic target. In this study, we summarize biological role of NAG-1 and discuss the potential molecular basis for their expressional regulation.
(Cancer Prev Res 13, 12-17, 2008)
Key Words: NAG-1, Macrophage inhibitory cytokine-1 (MIC-1), Apoptosis, Chemopreventive agents
서 론
NAG-1 유전자는 혈액암세포인 U937세포에서 대식세 포의 활성화에 관여하는 유전자들을 subtraction hybridiza- tion 방법을 이용하여 탐색하는 과정에서 처음으로 동정 되어 macrophage inhibitory cytokine-1이라 명명되었고 아 미노산 서열분석결과 TGF-β superfamily에 속하는 단백 질의 특성을 가지는 것으로 알려졌다.
1)또 다른 그룹은 대장암 세포에서 indomethacin에 의해 유도되는 유전자
를 PCR-based subtraction hybridization 방법으로 탐색하는
과정에서 동정하여 nonsteroidal anti-inflammatory drugs
(NSAIDs) activated gene (NAG-1)이라고 명명하였다.
2)그
외에도 이 유전자는 여러 그룹에서 다양한 방법에 의해
확인되어 placental transformation growth factor-b (PTGFB),
prostate derived factor (PDF), growth differentiation factor 15
(GDF- 15), placental bone morphogenetic protein (PLAB) 등
으로 명명되었다. 이러한 다양한 명칭은 이 단백질이 생
리학적으로 여러 기능을 가지고 있음을 암시해 준다. 지
금까지 NAG-1 단백질은 여러 암세포에서 다양한
Fig. 1. The expression levels of NAG-1 by treatment with various phytochemical compounds in HT29 cells. HT29 cells were treated with the indicated concentrations of baicalein (BE), quercetin (Quer), luteolin (Luteo), resveratrol (Res), cur- cumin (Cur) and triptolide (Trip).
chemopreventive agents, peroxisomal proliferator-activated receptor (PPAR) agonists, cyclooxygenase (COX) inhibitors 등 에 의해 발현이 증가하고 이 증가가 세포사멸을 촉진한 다고 알려져 왔다. NAG-1 단백질은 태반에서 많이 발현 되며 그 외에도 신장, 췌장, 전립선, 대장 등에서 약하게 발현되는 것으로 보고되었다.
3∼5)그러나 acute injury, 염 증, 암등의 병적 상태가 되면 NAG-1 단백질의 발현이 급격히 증가된다고 보고되었다.
5∼7)여러 agents에 의해 이 단백질의 발현이 증가하면 암세포에서 세포사멸을 촉진시킨다는 여러 보고들은 이 단백질이 암 억제 기능 을 가질 가능성을 보여주지만 한편으로 다양한 종양조 직에서 그 발현이 증가되어 있다는 사실은 NAG-1이 암 의 진행에 따라 다른 기능을 가질 수도 있음을 암시해준 다. NAG-1이 TGF-β superfamily에 속하는 단백질이라는 것을 고려해 볼 때 이는 예상할 수 있는 특징이긴 하나 아직까지 이 단백질의 생물학적 활성이 무엇인지에 명 확하지 않고 그 수용체에 대한 보고도 아직 밝혀지지 않 고 있다. 그러므로 이 단백질에 대한 깊은 연구와 이해는 암의 진행 및 암세포의 세포사멸 기전을 이해하는데 많 은 도움을 줄 것이라고 기대되어진다.
본 론
1. NAG-1 단백질은 TGF-β superfamily의 diver- gent member이다.
NAG-1 유전자는 19번 염색체의 p12.1∼13.1에 위치하 며 두 개의 exon으로 구성된다. Exon 1은 71 bp의 5’ un- translated region (UTR)과 238 bp의coding region을 포함하 고, exon 2는 647 bp의 coding region과 244 bp의 3’- untranslated region (UTR)을 포함하며, 이 두 exon은 1,820 bp의 intron에 의해 구분되어 진다.
3)NAG-1 단백질은 비 활성의 precursor로 합성되고 소포체에서 disulfide-linked dimerization된 후 골지체로 이동한다. Furin-like convertase 가 이 dimeric precursor의 RXXR site를 절개함으로써 생성 된 24.5 kDa의 dimeric 단백질이 세포 밖으로 분비된다.
1)NAG-1 단백질은 매우 보존된 약 80개의 아미노산으로 구성된 seven-cysteine domain을 가지는데 이는 TGF-β su- perfamily 단백질들이 공통적으로 가지는 특징이다. TGF- β superfamily에 속하는 다른 단백질들과의 multiple- sequence align 결과는 NAG-1의 cysteine residue들이 다른 단백질들과 약 15∼29 %의 일치함을 보여준다. 많은 TGF-β superfamily 단백질들에서 보존되어 있는 두 개의 또 다른 sequence인 DLGWW와 PCCVP도 NAG-1 단백질 에서 보존되어 있는 것을 확인할 수 있다.
1)이러한 단백
질 서열상의 여러 특징들은 NAG-1이 TGF-β superfamily 의 divergent member임을 보여준다. NAG-1은 다른 TGF- β superfamily의 단백질들이 가지는 일반적인 기능을 공 유하는데 예를 들면 TGF-β1 knockout mice는 광범위한 염증의 발생으로 죽는데 이는 TGF-β1의 주된 기능이 항염증 작용임을 나타낸다. NAG-1 단백질을 전처리하 였을 경우에 대식 세포에서 LPS에 의해 증가된 TNF-α 의 분비를 감소시켜 항염증 작용을 나타낸다. 이는 TGF- β1의 전처리와 같은 효과를 보여 준다.
1)또한 NAG-1의 발현은 primitive hemopoietic progenitors와 여러 상피 암 세 포주의 증식을 저해한다고 보고되었는데 이는 NAG-1이 다양한 활성을 나타내는 cytokine임을 보여준다.
2,8∼10)2. NAG-1 발현 조절 기전
앞에서 언급한 바와 같이 NAG-1 단백질은 NSAIDs, COX inhibitors, PPAR agonist등 다양한 자극에 의해 발현 이 증가하는 것으로 보고되었다. 특히 대장암 세포에서 항 종양물질인 indomethacin, resveratrol, genistein, diallyl disulfide, quercetin, PPAR agonist인 troglitazone, PGJ2 (15- deoxy-Δ
12,14-prostaglandin J
2), COX-2 저해제인 NS-398, DFU 등에 의해 NAG-1 발현이 증가되는 것이 보고되었
다.
2,11∼15)또한 전립선 암세포, 폐암세포, 위암세포와 유
방암세포에서도 다양한 자극에 의한 NAG-1 발현 증가 가 보고되었다.
16∼18)다양한 cancer chemopreventive 기능을 함유한 phytochemical 화합물을 대장암 세포주에 처리한 결과 quercetin, luteolin, resveratrol 및 curcumin에 의하여 NAG-1 발현이 증가됨을 확인하였다(Fig. 1). 여러 암세포 에서 다양한 자극에 의해 NAG-1 발현이 증가한다는 많 은 보고는 다양한 기전에 의하여 NAG-1 단백질의 발현 조절에 관여하고 있음을 시사해준다.
NAG-1 유전자가 19번 chromosome P arm (19p12∼13.1)
에 위치함이 in site hybridization을 통해 밝혀졌고,
3)그 후
인간 염색체 19번 특이적인 library를 이용한 screening을
통하여 이 유전자의 promoter를 클로닝하였다.
19)클로닝
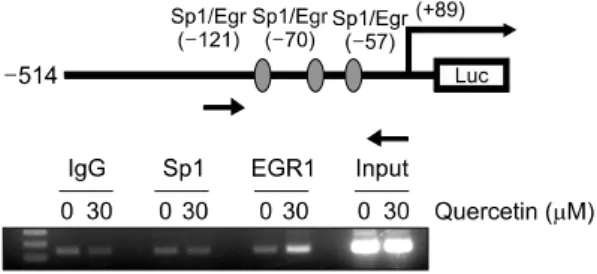
Fig. 2. Schematic structure of NAG-1 promoter construct and ChIP assay. EGR-1 is associated with quercetin-mediated up-regulation of NAG-1. Chromatin immunoprecipitation analyses were performed with anti-Sp1 antibody and anti-EGR-1 antibody. The DNAs were extracted from total sonicated nuclei (Input), protein A without antibody ( ), protein A with antibody (anti-Sp1 or anti-EGR-1), or with pre-immune serum (anti-IgG).
Specific promoter regions of the NAG-1 genes were amplified by PCR, separated on 1.5% agarose gels, and stained with ethidium bromide. Also indicated is the relative position of the PCR product generated in ChIP assay.
된 NAG-1 promoter를 분석해 본 결과 Sp1, AP-1, c-Myc, NF-κB, PU-1 등의 cis-acting elements 등을 포함하고 있다 는 것을 알 수 있었다. 특히 NAG-1 promoter의 120에서
50 사이에 세 개의 Sp1 site가 있고 이 들 Sp1 site가 promoter의 basal activity에 중요함이 밝혀졌다.
19)뿐만 아 니라 세포주기 조절과 그 이상에 의한 세포사멸 조절에 중요한 전사인자인 p53의 cis-acting element도 NAG-1 promoter region에 존재하고 이 유전자가 p53의 target gene 임이 보고된 바 있다.
9)또 다른 보고들에서도 대장암세 포에서 genistein, diallyl disulfide, resveratrol에 의해 유도되 는 세포사멸에서 p53에 의존적으로 NAG-1 발현이 증가 됨을 밝혔다.
11∼13)그 외에도 여러 전사 인자가 NAG-1의 발현 조절에 관여 하는 것으로 보고되었는데 특히 본 연 구실에서 quercetin에 의한 세포 사멸에서 NAG-1이 증가 되는데 p53과 더불어 전사인자 EGR-1이 중요한 역할을 한다고 보고하였다. EGR-1은 NAG-1 promoter의 basal activity에 중요한 전사인자인 Sp1과 cis-acting element를 공유하는 것으로 알려져 있고 PPAR agonist, troglitazone 에 의한 NAG-1 증가에 중요하다고 보고되었다.
14,15)NAG-1 promoter에 위치하는 3개의 Sp1 site의 각각을 두 bases를 다른 것으로 치환하면 quercetin에 의한 promoter 활성 증가 정도가 감소하는 것을 확인하였고 이 때 관련 된 전사인자를 확인하기 위해 chromatin immunoprecipi- tation (ChIP) assay를 해본 결과 quercetin에 의해 Sp1이 아 닌 EGR1의 NAG-1 promoter에 대한 binding이 증가됨을 확인하였다(Fig. 2).
ATF-3, NF-κB 등도 NAG-1 발현 조절에 관여함이 밝
혀졌고,
15,17)최근의 또 다른 보고는 유방암 세포주에서
미생물의 대사산물인 prodigiosin에 의한 NAG-1 발현에 glycogen synthase kinase-3β (GSK-3β)의 활성이 필요함을 보여준다.
20)3. 여러 암에서 NAG-1의 발현
암세포의 세포사멸을 유발하는 많은 물질들에 의해 NAG-1의 발현이 증가하고 이로 인해 세포 사멸이 증가 한다는 많은 보고들과 상반되게 암 조직에서 이 단백질 의 발현이 증가되어 있다는 여러 증거들이 있다. 특히 대장암과 전립선암에서 이 단백질의 발현이 매우 증가 되어 있으며 이러한 혈청 NAG-1의 발현 정도는 암의 진 행과정과 매우 밀접한 연관성이 있음이 보고되어 있
다.
21,22)또 다른 보고에 의하면 그 외에도 흑색종, 췌장
암, 갑상선암에서도 이 단백질의 발현이 증가되어 있음 이 보고되어 있다.
23,24)그러나 이와는 반대로 전립선암의 초기단계의 80%에서 NAG-1의 발현이 감소되어 있음이 보고되었다.
25)이러한 상반되는 결과는 지금까지 명확하 게 설명되지 못하고 있으나 이는 NAG-1 단백질이 암 진 행 과정에 따라 다른 작용을 할 가능성을 보여준다.
암의 진행과정과 연관된 NAG-1 단백질의 발현 정도 는 이 단백질이 전립선암, 갑상선암, 췌장암 대장암 등의 진단과 모니터링에 적용될 수 있음을 보여준다. 특히 췌 장암의 경우 진단이 매우 어렵고 생존율이 낮을 뿐만 아 니라 일반적인 종양 marker가 도움을 주지 못하는데 혈 청 내 NAG-1과 CA19-9를 함께 측정하여 진단에 이용할 경우 그 민감도가 70%에 이르고 정확도가 85%에 이른 다는 보고가 있다.
24)혈청 내 NAG-1의 발현 정도와 더불 어 이 유전자의 polymorphism도 암 진단에 이용될 수 있 음을 보여주는 보고들이 있다.
21,26)NAG-1 유전자에서 아 미노산의 치환을 일으킨다고 보고된 single nucleotide polymorphism (SNP)은 V9L, S48T, H202D 등 세 가지로 이 중 마지막 SNP (H202D)는 mature NAG-1 단백질의 6번째 아미노산이므로 H6D로 언급되기도 한다.
27)이 H6D polymorphism은 base G가 C로 치환되면서 히스티딘 (H)이 아스파르트산 (D)으로 바뀌는 것으로 이 유전자를 coding 하는 allele중 D allele (아스파르트산)를 하나 이상 가지고 있는 환자의 경우 대장암의 전이 위험이 좀 더 높고 재 발되는 기간은 짧아지지만 전반적인 생존 기간은 길어 진다.
21)4. 암의 진행과 NAG-1의 역할
앞에서 언급한 것처럼 NAG-1 발현이 암 조직에서 많
이 증가되어 있다는 여러 보고가 있지만 아직까지 이 단
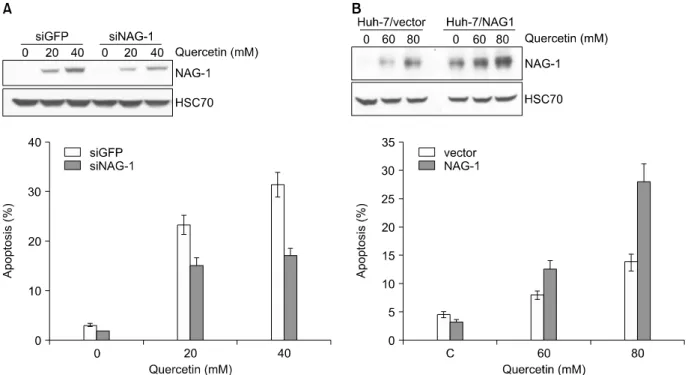
Fig. 3. Suppression of NAG-1 expression attenuates quercetin-induced apoptosis in HCT116 cells and enhanced expression of NAG-1 in Huh-7 cells promotes quercetin- induced apoptosis. (A) Down-regulation of NAG-1 by transfection with siRNA against NAG-1. HCT116 cells were transfected with NAG-1 siRNA or GFP siRNA. Down-regulation of NAG-1 reduces quercetin-induced apoptosis in HCT116 cells. Apoptosis was analyzed as a sub-G1 fraction by FACS. (B) Huh-7/NAG-1 and Hun-7/vector cells were treated with 60 or 80μM quercetin for 24 h, and apoptosis was analyzed as a sub-G1 fraction by FACS. The proportion of apoptotic cells is indicated.
백질이 암 진행에 미치는 영향에 대해 명확한 해답은 없 다. 반면에 여러 chemopreventive agent, PPAR agonist, COX inhibitor 등에 의한 세포사멸에서 증가된 NAG-1은 이 세 포사멸을 촉진시켜 항암작용을 도와준다는 많은 보고들 이 있다.
14,16,17)Chemopreventive agent인 quercetin에 의한 대장암세포의 사멸에서 NAG-1의 발현이 증가되는 것을 확인하였고 siRNA를 이용하여 이 단백질의 발현의 억제 하였을 경우 quercetin에 의한 세포사멸이 저해되고, NAG-1 단백질이 과발현된 세포에서 quercetin에 의한 세 포사멸이 증가되는 것을 확인하였다(Fig. 3).
최근에 NAG-1 transgenic mice를 이용한 실험에서 NAG-1 과발현이 intestinal neoplasia를 억제한다고 보고되 었다.
28)본 연구실에서 보고하지 않은 결과에 따르면 ER stress를 일으키는 여러 물질들에 대한 세포사멸에서 NAG-1의 발현이 증가하며 NAG-1이 과발현된 세포에서 이들 물질들에 의한 세포사멸이 증대되는 것을 확인하 였다(Fig. 4).
이는 지금까지 보고된 여러 내용들과 일치하는 것으 로 발현이 증가된 NAG-1이 항암 작용을 나타낸다는 것 을 보여준다. 최근에 여러 보고는 NAG-1이 암의 성장을
조절하는 기전에 대한 여러 증거를 보여주고 있다.
NAG-1은 유방암에서 cyclin D1의 발현을 억제하고 난소 암세포에서 부분적으로 p21의 조절을 통해 세포 성장을 억제한다.
29,30)전립선암 세포 DU145에 recombinant NAG-1 을 처리하면 세포의 부착을 방해하고 anti-apoptotic 단백 질의 발현을 감소시켜 세포사멸을 유도한다.
31)결 론
암세포에서 NAG-1의 발현이 증가하면 세포사멸이 촉
진되어 이 단백질의 항암작용을 보여주는 여러 보고가
있지만 아직까지 그 작용기전에 대한 설명은 많이 부족
하다. 뿐만 아니라 NAG-1의 암 세포의 사멸에 있어서
그 작용에 대한 많은 연구는 암 조직에서 이 단백질의
발현이 증가되어 있는 이유를 명확히 설명해주지 못한
다. 이러한 상반되는 결과는 이 단백질이 세포 내 생리적
상태에 따라 다른 작용을 할 가능성을 보여준다. 이 단백
질의 발현이 다양한 인자들에 의해 복잡하게 조절 된다
는 것을 보여주는 여러 기전에 대한 많은 연구 결과는
이러한 가정을 뒷받침해준다. 특히 잘 알려진 tumor
Fig. 4. Overexpression of NAG-1 enhances ER stress- mediated apoptosis. (A) Huh-7 cells, stably transfected with pcDNA3.1 and NAG-1, were exposed to brefeldin A (BFA), thapsigargin (TG), deoxycholic acid (DOC), and cisplatin for 24 h. Apoptosis was analyzed as a sub-G1 fraction by FACS.
(B) pcDNA3.1 and NAG-1-transfected Huh-7 cells were incubated with BFA, TG, DOC, and cisplatin for 24 h. Cell lysates were prepared and subjected to Western blot analysis using NAG-1 and HSC70 antibodies.