총 설
생물학적 수소생산 공정
신종환·박태현† 서울대학교화학생물공학부
151-742 서울시관악구신림동산56-1 (2005년 12월 20일접수, 2006년 2월 6일채택)
Biological Hydrogen Production Processes
Jong-Hwan Shin and Tai Hyun Park†
School of Chemical and Biological Engineering, Seoul National University, San 56-1, Shilim-dong, Gwanak-gu, Seoul 151-742, Korea (Received20 December 2005; accepted 6 February 2006)
요 약
생물학적수소생산공정은다른열화학적공정이나전기화학적공정에비하여환경친화적이며에너지를덜소모하 는공정이다. 생물학적수소생산공정은크게두가지로구별할수있는데, 광합성에의한수소생산과혐기발효에의 한수소생산이그것이다. 광합성에의한수소생산공정은주로물로부터수소를생산하고동시에공기중의이산화탄 소도저감하는특징을가지고있으며, 혐기발효에의한수소생산공정은유기탄소원을섭취하는박테리아에의한발 효를통해이루어지는공정이다. 본논문에서는생물학적수소생산공정에대한그간의연구들에대하여살펴보았다.
Abstract −Biological hydrogenproductionprocesses are more environment-friendly and less energy intensive than thermo- chemical and electrochemical processes. The biological process can be divided into two categories: photosynthetic hydrogen production and hydrogen production by dark fermentation. Photosynthetic process produces hydrogen mainly from water and reduces CO2 simultaneously. Dark fermentation is a dark and anaerobic process that produces hydrogen by fermentative bac- teria from organic carbon. The article presents a survey of biological hydrogenproductionprocesses.
Key words: Biological Hydrogen Production, Photosynthetic Hydrogen Production, Dark Fermentation
1.
서 론오늘날우리는에너지부족과환경오염이라는두가지위기에직 면해있다. 세계의에너지수요는대부분화석연료에의존하고있 으며이로인한화석연료의편재화와고갈로인해여러가지정치·
경제·사회적문제들을야기하고있다. 또한, 화석연료의사용으로 배출되는배기가스는지구온난화및환경오염문제들을야기하고 있다. 이런현실에서여러국가는이위기를해결하기위해대체에 너지를개발하기위한연구에몰두하고있다. 이러한문제점에대 한해결책의하나로서수소에너지에대한관심이고조되고있다. 수 소에너지는태양광, 태양열, 석탄, 석유등과같은 1차에너지를변 환시켜얻을수있는 2차에너지에해당하며, 단위질량당에너지함
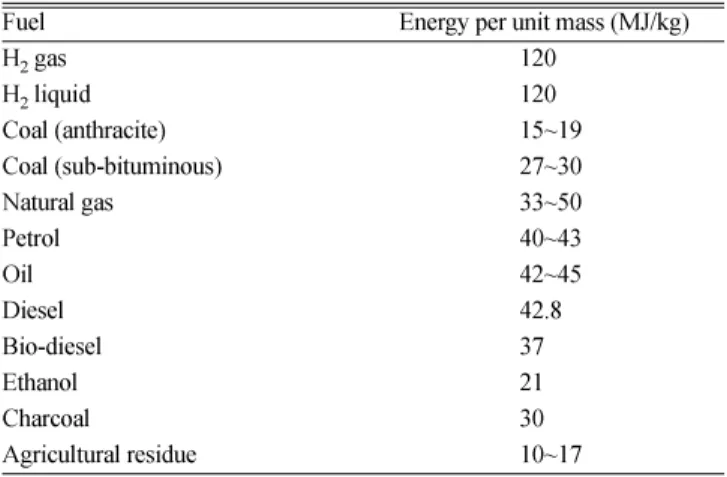
유량이매우높고(Table 1) 연소부산물로서주로물을발생시키므로
어떤다른에너지원보다더욱환경친화적인청정에너지라고할수
있다[1, 2]. 또한, 수소에너지는연료전지의개발에힘입어더욱그
필요성이증대되고있으며군사/우주개발용뿐만아니라민간/상업 용으로주목받고있다.
현재수소는 96%이상이화석연료로부터제조되고있으나, 이또 한매장량의한계와환경오염물질배출로인해궁극적인제조방 법이라고할수는없다. 따라서더욱더환경친화적인태양광, 수 력, 풍력, 미생물과같은청정기술을이용하여수소를제조하는방
†To whom correspondence should be addressed.
E-mail: [email protected]
Table 1 Comparison of energy per unit mass
Fuel Energy per unit mass (MJ/kg)
H2 gas 120
H2 liquid 120
Coal (anthracite) 15~19
Coal (sub-bituminous) 27~30
Natural gas 33~50
Petrol 40~43
Oil 42~45
Diesel 42.8
Bio-diesel 37
Ethanol 21
Charcoal 30
Agricultural residue 10~17
법이연구되고있다. 특히최근에는미생물을이용하여물이나유 기성폐자원으로부터수소를제조하는생물학적수소생산방법에 대한관심이점점증가하고있다. 생물학적인수소생산방법은화 학공학적인수소생산방법에비해상온·상압조건에서조업이이루 어지기때문에덜에너지집약적이며, 물이나바이오매스, 유기성 폐자원등과같은재생가능한연료로부터수소생산이이루어지기 때문에이론적으로는무한정수소생산이가능하다. 또한, 생물학적 수소생산방법은유기성폐자원처리, 이산화탄소저감등환경처 리기술과결합하여환경오염감소와대체에너지생산이라는두 가지장점을가지고있는기술이다. 따라서현재상용화되어있는 화학공학적방법에의한수소제조보다는궁극적으로위와같은 장점을갖는생물공학적인방법에의한바이오수소제조로전환 될것으로기대된다. 이논문에서는최근에집중연구되고있는생 물학적수소생산방법의종류및그와관련된미생물그리고현재 연구현황에관하여기술하였다. 또한, 이논문에서는보다실용화 에한발짝더다가선혐기발효공정에관해심도있게다루고자 한다.
2.
생물학적 수소 생산 공정미생물에의한수소생산연구는 1800년대부터시작되었으나기
초연구에국한되었고수소에너지와관련하여연구가시작된것은
1970년대부터이다. 즉, 에너지위기의시대에그대안으로수소에
너지를연구하기시작한것이다. 생물학적수소생산은주로미생 물을이용한방법으로서, 미생물의특성에따라크게두가지로분 류할수있다[3]. 하나는빛을이용하는광합성(photosynthesis) 미생
물에의한수소생산방법이고[4-6], 다른하나는빛을이용하지않
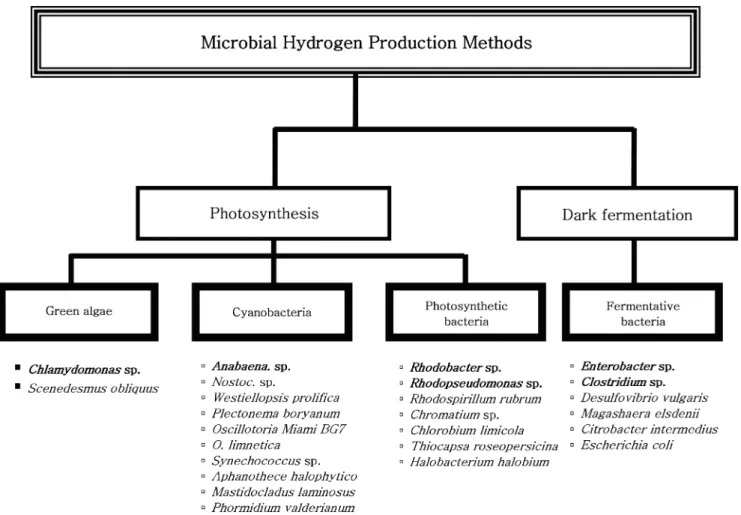
는미생물에의한혐기발효방법이다[7-9]. Fig. 1은다양한미생물
에의한수소생산방법을미생물의특성에따라분류한그림이다.
광합성을이용한수소생산방법에관여하는미생물은크게 green
algae, cyanobacteria(blue green algae), photosynthetic bacteria로구 분할수있으며, 혐기발효에관여하는미생물은 fermentative bacteria
가있다[3, 10]. 이들미생물은자신이가지고있는효소를이용하여
수소를생산하며[3], 수소생산에관여하는효소로서 hydrogenase
(H2ase), nitrogenase(N2ase), formate hydrogen-lyase(FHL) 등이알 려졌다[11-16].
따라서, 수소생산미생물에의한수소생산조건최적화 연구 와 함께 그효소에 관한연구들이 진행되고있다. 그러나이들 효소에관하여충분한 연구가되어있지는않은실정이다. 그러 므로앞으로는미생물을통한수소생산뿐아니라, 수소생산에직 접관여하는효소들을미생물로부터분리하여효소반응공정최 적화를 통해수소생산성을 높이려는보다 심도있는연구가필 요할것으로생각된다. 일부효소들은산소에매우민감하기때 문에현재그문제를해결하기위한노력이진행중이다. 다음으 로, 각 특성별미생물에의한수소생산방법에관하여기술하고 자 한다.
Fig. 1. Biological hydrogen production methods by various microorganisms.
3.
광합성 미생물에 의한 수소 생산지구상에풍부하게존재하고있는물과태양빛을사용하여수소 를생산한다면가장이상적인방법일것이다. 광합성미생물중에 는태양빛을이용하여물로부터수소를생산하는것이있다. 이는 지구상에무한히존재하는물과태양빛을이용하여에너지를생산 한다는의미에서중요한가치를지니고있다. 이들미생물은환경 오염의원인이되는이산화탄소를수소생산과정중에탄소원으로 사용하므로, 이산화탄소제거와수소에너지생산이라는일석이조의 효과를가진다. 이와관련된미생물로서 Green algae와 Cyanobacteria
가있으며각기다른메커니즘에따라수소를생산한다. 이들미생 물은매우이상적인시스템을갖추고있으나수소생산성이낮아실 용적인측면에서는아직갈길이멀다는문제점을가지고있다. 이 러한공정에서해결해야할가장큰문제점은광합성에의해발생 하는산소가수소생산에관여하는효소의활성을떨어뜨리는것이 다. 따라서이산소를제거하는방법에관한연구가여러방면에서 진행되고있다. 또한, 이와관련하여산소에덜민감한효소를개발 하기위해분자생물학적연구가시도되고있다. 이런문제들이해결 된다면수소생산성이높아져실용화에한걸음더다가설것으로 생각된다. 이와비교하여수소생산성이비교적높은광합성미생물 로서, photosynthetic bacteria가있다. 이박테리아는비록물로부터 수소를생산하지는않고유기탄소원을공급해주어야하지만, 태양 광을이용한다는관점에서는이범주에속하게된다.
3-1. Green algae
Green algae는광합성메커니즘(photosystem I, II)에의해물로부
터양성자와전자를공급받아수소를생산하는직접물분해(direct
biophotolysis) 방법을이용하고있다[10, 17]. 즉, algae 내에있는 엽록체에서가시광선을받고이빛에너지에의해서물이분해되어 산소, 양성자(H+), 전자(e−)를발생한다. 빛에너지는물에서발생한
전자를고준위의에너지로만들고 ferredoxin(Fd)과같은몇단계의
전자전달경로를거쳐최종적으로수소생산효소로전달된다. 이 효소는촉매작용을하여양성자와전자를통해수소를발생시킨다.
Fig. 2는이와같은직접물분해에의한수소생산공정을알기쉽
게나타낸그림이다[18]. Fig. 2에서나타낸바와같이이러한직접
물분해공정에관련되는중요한효소는 hydrogenase이며이효소
는산소에매우민감한단점을가지고있다. 이는앞에서언급한바
와같이광합성과정에서발생하는산소가 hydrogenase의활성에
영향을주어수소생산을저해시킨다. 따라서이러한단점을보완하 기위해최근에는황이없는배지에서 Chlamydomonas reinhardtii
를배양하는공정이개발되었으며, 이는광합성과정에서발생하는 산소를점차감소시켜혐기상태로변화시키는기술이다[19]. 이밖 에도낮과밤의주기를최적화하여수소를증가시키는연구가보고 되었으며[20], 유전자조작을통해산소민감성이감소한 hydrogenase
를갖은균주를개발함으로써수소생산성이향상된결과가보고되 었다[21].
3-2. Cyanobacteria
Cyanobacteria(blue green algae)는광합성에의해물을분해하여 산소를발생하고, 동시에공기중의이산화탄소를고정하여고분자 저장물질(cell material)로균체내에합성한후혐기발효또는광합 성발효에의해수소를생산하는간접물분해(indirect biophotolysis)
방법을사용한다(Fig. 3) [10, 18, 22]. 이러한수소생산공정의핵심
효소는질소고정화효소인 nitrogenase로써이는산소와질소가없
는조건에서수소를생산한다. 그러나 nitrogenase는수소를생산하는 데 ATP를이용하기때문에 hydrogenase에비해덜효율적이다. 또한,
이러한공정에서는수소를소비하는효소인 uptake hydrogenase가존 재하기때문에수소생산량이낮아진다. 따라서, 이러한문제점을해 결하기위해 Cyanobacteria의일종인Anabaena PCC 7120으로부터
uptake hydrogenase의기능을없앤 AMC 414(a Hup minus mutant)
의배양이연구되어대기상에서도수소를생산할수있었고, 기체 상을대기에서 Ar으로치환함으로써수소생산성을향상시키는연 구도보고되었다. 이와같은방법을이용하여옥외배양에서최대수 소생산 14.9ml H2/h/l를얻었다[23]. 또한, Anabaena PCC 7120에서
∆hupL(uptake hydrogenase 결여), ∆hoxH(bidirectional hydrogenase
결여)와∆hupL/∆hoxH(2개의유전자결여) 등의다양한 hydrogenase
mutant들을제작함으로써수소생산능력을향상시키는연구고수
행되었다. ∆hupL과 ∆hupL/∆hoxH의경우가야생형보다최적조건
에서 4~7배수소생산속도가높았다[23]. 이밖에질산염제약조건
과빛, 산소가없는상태에서Gloeocapsa alpicola에관한연구가보
고되었다[25]. 이러한유전자조작및배양조건에의한수소생산향
Fig. 2. Direct biophotolysis of water by Green algae. Fig. 3. Indirect biophotolysis of water by Cyanobacteria.
상외에도미생물성장단계와수소생산단계를나누는 2단계조 업에의한수소생산을향상시키려는연구가수행되었다[26].
3-3. Photosynthetic bacteria
Purple non-sulfur bacteria는유기산으로부터광합성발효(photosynthetic fermentation)에의해수소를생산한다(Fig. 4) [10, 18]. 즉, 광합성 박테리아는조류및식물이광합성계(photosystem, PS) I, II를모두이 용하는것과는달리 PS I(bacterial photosystem)만을이용하여광합성 작용을하고수소를발생한다. 다시말해, bacterial photosystem에 존재하는색소복합체인반응쎈터에있는클로로필과카로티노이드 색소에서빛에너지를흡수하여반응쎈터복합체의양면의전위차
로전환하며, 이러한전위차는 cyclic 전자전달계를생기게하고이
것은다시 ATP 등의고에너지화합물을만들게된다. 이때 organic acid와같은기질이공급하는전자가 ferredoxin을환원하며, 이환 원력과 ATP를이용하여 nitrogenase가질소원이없는조건에서분 자상의수소를발생한다. Fig. 4는이러한과정을나타낸그림이다[18].
이러한광합성발효공정의최적화를위해Rhodobacter sphaeroides O.U. 001을사용하여, 접종균주의나이에관한연구[27], 조사하는
빛에관한연구[28], 배양배지를최적화하는연구[29] 등이수행되
었다. 이밖에수소생산에저해를주는산소를제거하기위한연구
[30]와세포고정화를통한수소생산성향상[31] 및광배양기의최 적화에관한연구[32] 등이보고되었다.
4. Dark fermentation
혐기발효에의한수소생산은빛이없는혐기적조건에서유기물 을분해하여수소를생산하는공정이다. 기존의유기물분해공정
(anaerobic treatment process)은 산성화(acidification)와 메탄생성
(methane production) 공정으로나눌수있다. 산성화공정의경우 는부산물로서수소를생성하며이수소는여러가지산업적으로
응용되었으며[33], 최근에는연료전지를위한청정에너지원으로
사용되고있다[34]. 메탄생성공정에서는생성된메탄이전기생산
을위한연료로서사용되고있다[3]. 그러나이러한메탄보다는수
소가더연료전지에서에너지로의전환효율이좋기때문에앞의 두공정보다는수소생산량이더많은수소생산공정을통해유기 물을분해한후여기서생산된수소를청정에너지로사용하려는연 구가진행되고있다. 이러한혐기발효에의한수소생산공정은광
합성을이용한공정에비해 (1) 높은수소생산속도및미생물성장
속도, (2) 빛없이수소생산가능, (3) 폐수자원이용가능등의장점 을가지고있다. 그러나단점으로는 (1) 낮은수소생산수율(yield), (2) 발생가스중에 CO2함유등이있다.
수소생산을위한혐기발효에관여하는미생물은 fermentative bacteria로서산소노출에매우민감한절대혐기성균주(strict anaerobe)
와산소에덜민감한통성혐기성균주(facultative anaerobe)로나눌수 있다. 이들혐기발효균주를이용한수소생산은 sewage sludge 또 는 토양으로부터미생물을 분리하여 사용하는 순수배양(pure culture) 또는복합배양(mixed culture)이수행되고있다. 최근에는
sewage sludge 또는토양으로부터여러우수균주들이분리되어수
소생산을위한최적조건연구및균주개량, 공정개발등에이용되 고있다. 더욱많은연구를통해더욱다양하고우수한균주들이분 리될것으로기대된다. 이와더불어고온성미생물을이용한수소 생산연구가진행되고있다. 고온성미생물은다른미생물에비해 수소이외의부산물이적어서이론적수율에가장가까운균주이다.
그러나고온을유지하여야하고주로절대혐기적균주이기때문에 그공정이매우까다롭다. 앞으로이런공정상의단점을극복한다 면높은수소생산성을위한좋은연구분야가될것으로기대된다.
미생물을이용하여혐기발효를통해수소를생산하는방법에는
pyruvate 분해 metabolism에따라다음과같은두가지로나눌수 있다[3].
1. Pyruvate: formate lyase (PFL) system:
(Pyruvate + CoA)→acetyl-CoA + formate 2. Pyruvate: ferredoxin oxido reductase (PFOR) system:
Pyruvate + CoA + 2Fd(ox)→acetyl-CoA + CO2+ 2Fd(red) Enterobacteria와같은통성혐기성미생물은주로 PFL system에 의해 pyruvate로부터 formate가생성된후 FHL에의해수소와이 산화탄소로분해된다.
HCOOH (formate)FHL→H2+ CO2
이에반해Clostridia와같은절대혐기성미생물은위에서설명
된 pyruvate 분해 metabolism 중 PFOR system에의해환원된상태 의 ferredoxin을얻고이로부터전자를전달받아 glycolysis 과정중 생성된 NADH를 hydrogenase가 reoxidation시켜수소를생산한다.
H2ase
NADH + H++ Fd(red)→H2+ NAD++ Fd(ox)
두가지 pathway 중전자는 formate pathway이며, 후자는 NADH pathway이다[16, 35].
이러한혐기발효연구의주된목표는수소생산성증대및수율
(yield)의향상이다. 수율향상을위해서는이론적인수율인 glucose Fig. 4. Photodecomposition of organic compounds by photosynthetic
bacteria.
1 mol 당 12 mol의수소를생산하는것을목표로연구가진행되고 있다. 그러나열역학적인관점에서 glucose 발효를통해 acetate가 주된 metabolite라가정할때이론적인수소생산량은 4 mol이다[3].
이러한이론적인 4 mol의수소도여러가지 metabolite(lactic acid, succinic acid, butyric acid, ethanol, butanediol, etc)가생산되는방 향으로진행되는 metabolic flux에의해더낮은수율을나타낸다.
또한, 이들대사산물들은다시 metabolic pattern에영향을준다. 따
라서수소의생산수율을높이기위해서는각미생물의 metabolic
pattern에대한연구도필요하다. 대사산물은배지의조성, pH, 온도 등에영향을받기때문에이들조건에따른 metabolic pattern 연구 가선행되어야할것이며생성된대사산물의농도에의한영향도 함께연구되어야할것이다. 이밖에배양기내의수소분압은미생
물의성장 pattern에영향을주어미생물성장과수소생산을저해
하기때문에수율이더낮아진다[36]. 이는생산물저해를나타내
며이는절대혐기성균주일수록더큰영향을나타낸다. 따라서생 산물저해를줄이기위해배양기내의수소를제거하는방법에관 한연구가필요하다. 이는공정개발을통한고농도배양시더큰 문제를야기할것으로예상되기때문에반드시해결해야할문제이 다. 이밖에수율을향상시키기위해서배양조건최적화, 대사공학,
분자생물학적방법등을이용하는연구가수행되고있다. 다음은각 각의 fermentative bacteria에대해어떤연구가진행되고있는지를 살펴보고자한다.
4-1. Strict anaerobe
절대혐기성균주에의한수소생산연구는주로Clostridium sp.
를이용하고있다. 이러한공정에서는산소에매우민감하기때문 에외부산소와의접촉을최소화하는것이중요하다. 따라서이를 위해배지내에환원제를넣어준다. 절대혐기성균주를이용한수
소생산공정에관여하는핵심효소는 hydrogenase이다. 이효소는
산소에매우민감하기때문에산소와의접촉이없는절대혐기적 조건에서만수소를생산한다. 이공정에의한수소생산수율은통 성혐기성균주에의한것보다일반적으로더높아서이론적수율 에더가깝다. 그러므로 sewage sludge 또는토양으로부터우수균 주를분리하거나기존에알려진균주를통해수소생산향상을위한 연구가진행되고있다[8, 37, 38]. 가장최근에분리된 Clostridium butyricum CGS5는 pH 5.5와 20g COD/l의조건에서 2.91 mol H2/mol sucrose의최대수율을얻었으며 pH 5.5와 20 g COD/l의조건에서 는 209 ml/l/h의최대수소생산속도(volumetric H2 production rate)를 나타낸결과를보고하였다[38]. 이밖에 C. acetobutylicum[39]은
1.97 mol H2/mol glucose의수율을보고하였고C. thermolacticum은
1.5 mol H2/mol glucose의수율과 2.58 mmol/l/h의 H2 productivity
를보고하였다[40]. 또한, 더높고지속적인수소생산을위해연속 배양이나세포고정화를통해수소생산량향상을보고하고있다. 이 와함께세포내효소의특성에대한이해가중요하기때문에, 효소를 세포밖으로분리하여효소특성을파악하려는연구가진행중이다.
4-2. Facultative anaerobe
통성혐기성균주에의한수소생산연구는주로Enterobacter sp.
를이용하여수행되고있다. 통성혐기성균주의장점은산소에덜 민감하여배지내에고가의환원제를넣어줄필요가없는등공정
이비교적단순하여 scale-up시손쉽다는점을들수있으며단점으
로는절대혐기성균주에비해수소생산수율이낮다는점을들수 있다. 그러나수율은낮지만높은수소생산속도와공정의단순함 으로인해수소생산을위해널리사용되고있다[9, 16, 35, 41, 42].
이러한통성혐기성균주는앞서말한바와같이 PFL system에의
해 formate를만들고 FHL에의해수소와이산화탄소가생성된다.
이와함께 glycolysis과정에생성된 NADH를 reoxidation하여수소 를생산하는 NADH pathway 또한존재한다고알려졌다[10, 35].
Enterobacter cloacae IIT-BT은, 2.2 mol H2/mol glucose의 수율과
35.62 mmol H2/l/h의 수소생산속도가보고되었다[43]. 이 밖에
Enterobacter aerogens E82005는 Ar gas purging을통해 1.58 mol/H2/mol molasses의향상된수율이보고되었으며[35] Enterobacter aerogens HU-101은 0.56 mol H2/mol glucose의수율그리고그것의 mutant
인 AY-2를통해 1.17 mol H2/mol glucose의수율향상이보고되었
다[44]. 또한, 연속배양과세포고정화연속배양에의해지속적인수
소생산을얻는연구들이수행되고있다[45, 46].
4-3. Thermophilic bacteria
고온성미생물을이용한수소생산에대한연구는, 수소가원하 지않는발효부산물로여겨졌기때문에[47, 48] 지금까지많이진행 되지않았다. Pyrococcus furiosus[49], Pyrococcus abyssi ST549[47], Thermotoga maritime[48], Acetothermus paucivorans[50], Acetomicrobium flavidum[51] 등이수소를생성할수있는균주로알려졌으나수소 생산효율에대한연구는최근들어서활발히진행되고있다. 수소 생산자로서의고온성미생물을고려해볼때, 흥미롭게도고온성미 생물은중온성통성혐기성균주나절대혐기성균주에비해수소 생산수율이월등히높다[52]. 이론적으로발효를통해 1 mol hexose
당 4 mol hydrogen을얻을수있는데, Thermotoga maritima의경 우는최대이론값인 4 mol hydrogen/mol glucose를얻을수있다고 보고된 바가 있다[48]. Caldicellulosiruptor saccharolyticus와
Thermotoga elfii의경우에, 이균주들은양론적으로 1 mol hexose당
3.3 mol hydrogen의수율을나타내어이론적인수소생산수율의
83%의값을달성했다[53]. 또한, maximal hydrogen production rate
이각각 8.4 mmol/l/h와 4.5 mmol/l/h의우수한값을나타내었다. 또 한, 일본의 Tamotsu는 Thermococcus kodakaraensis KOD1로부터 연속배양을통해 59.6 mmol/g/h의수소생산성을달성하였다[54].
5.
결 론수소는더이상미래의에너지가아니다. 수소는화석연료를대 체하는에너지로서매우높은효율과환경친화적이기때문에실용 화가눈앞에와있다. 그중생물학적수소생산은기타화학공학적 방법에의한수소생산보다더욱환경친화적이며이산화탄소저감,
유기성폐자원처리등의장점을가지고있기때문에더욱주목받 고있다. 현재생물학적수소생산방법은아직해결해야할문제들 이있지만, 급속한진전이이루어지고있다.
광합성미생물에의한수소생산은물과태양광으로부터수소를 생산하는이상적인시스템으로광합성에의해생성된산소가수소 를생산하는효소를저해하는문제만해결된다면더많은수소생 산량을얻어실용화될것으로기대된다. 이는분자생물학적인방법 으로해결이가능할것으로생각된다.
한국, 일본과같은유기성폐자원이풍부한국가에서주목하고있
는혐기발효기술의실용화를위해서는수소생산수율을향상시켜 야한다. 혐기발효기술은수소생산과함께전세계적으로문제가 되고있는유기성폐자원을처리할수있다는큰장점을가지고있 다. 이러한엄청난양의유기성폐자원을모두에너지로전환한다 면에너지소비량의 30%가량을충당할수있다. 혐기발효공정이 실용화되기위한가장큰걸림돌은낮은수소생산수율이다. 따라 서수소의생산성과수율을높이는것이혐기발효연구의핵심이라 할것이다. 수율을향상시키기위한방안으로수소생산을위한배양 조건최적화, 변이주(mutant) 개발, 유전자조작(genetic operations),
대사공학(metabolic engineering) 등이적용되고있다. 이들방법에 의해점차수율의향상이이루어지고있으며지속적인수소생산을 위한공정개발도함께수행되고있다.
수소에너지의제조와함께수소에너지사회로전환하기위해서는 수소의저장수송을위한기술개발이필요할것이며이를실제이 용하기위한연료전지의개발과함께수소를안전한에너지원의하 나로인식하는사회적인식의변화가필요하다.
감 사
본연구는과학기술부의지원으로수행하는 21세기프론티어연 구개발사업의일환으로수행되었습니다.
참고문헌
1. Suzuki, Y., “On Hydrogen as Fuel Gas,”Int. J. Hydrogen Energy, 7(3), 227-230(1982).
2. Bockris, J. O. M., “The Economics of Hydrogen as a Fuel,”Int.
J. Hydrogen Energy, 6(3), 223-241(1981).
3. Vijayaraghavan, K. and Soom, M. A. M., “Trends in Biological Hydrogen Production—a Review,”Int. J. Hydrogen Energy, in press, available online at www.sciencedirect.com(2004).
4. Lichtl, R. R., Bazin, M. J. and Hall, D. O., “The Biotechnology of Hydrogen Production by Nostoc Flagelliforme Grown Under Chemostat Conditions,”Appl. Microbiol. Biotechnol.,47(6), 701-707 (1997).
5. Hansel, A. and Lindblad, P., “Towards Optimization of Cyano- bacteria as Biotechnological Relevant Producers of Molecular Hydrogen a Clean and Renewable Energy Source,”Appl. Micro- biol. Biotechnol., 50(2), 153-160(1998).
6. Matsunaga, T., Hatano, T., Yamada, A. and Matsumoto, M., “Microaer- obic Hydrogen Production by Photosynthetic Bacteria in a Dou- ble-Phase Photobioreactor,”Biotechnol. Bioeng., 68(6), 647-651 (2000).
7. Oh, Y.-K., Seol, E.-H., Lee, E. Y. and Park, S. H., “Fermentative Hydrogen Production by a New Chemoheterotrophic Bacterium Rhodopseudomonas palustris P4,”Int. J. Hydrogen Energy, 27(11-12), 1373-1379(2002).
8. Fumiaki, T., Chang, J. D., Mizukami, N., Tatsuo, S. T. and Kat- sushige, H., “Continuous Hydrogen Production by Clostridium sp. Strain No. 2 from Cellulose Hydrolysate in an Aqueous Two- phase System,”J. Ferment. Bioeng., 82(1), 80-83(1996).
9. Yokoi, H., Tokushige, T., Hirose, J., Hayashi, S. and Takasaki, Y., “Hydrogen Production by Immobilized Cells of Aciduric Enterobacter aerogenes strain HO-39,”J. Ferment. Bioeng., 83(5),
481-484(1997).
10. Das, D. and Veziroglu, T.N., “Hydrogen Production by Biologi- cal Processes: a Survey of Literature,”Int. J. Hydrogen Energy, 26(1), 13-28(2001).
11. Vignais, M. V., Billoud, B. and Meyer, J., “Classification and Phylogeny of Hydrogenases,”FEMS Microbiol. Rev., 25(4), 455- 501(2001).
12. Adams, M. W., Mortenson, L. E. and Chen, J. S., “Hydroge- nase,”Biochim. Biophys. Acta., 594(2-3), 105-176(1980).
13. Appel, J. and Schulz, R., “Hydrogen Metabolism in Organisms with Oxygenic Photosynthesis: Hydrogenases as Important Reg- ulatory Devices for a Proper Redox Poising,”J. Photochem. Pho- tobiol. B: Biol., 47(1), 1-11(1998).
14. Schulz, R., “Hydrogenases and Hydrogen Production in Eukary- otic Organisms and Cyanobacteria,”J. Mar. Biotechnol., 4, 16-22 (1996).
15. Boichenko, V. A. and Homann, P., “Photosynthetic Hydrogen Production in Prokaryotes and Eukaroytes: Occurrence Mecha- nism and Functions,”Photosynthetica.,30, 527-552(1994).
16. Gorman, J., “Hydrogen: the Next Generation,”Science News(2002).
17. Melis, A., Zhang, L., Forestier, M., Ghirardi, M. L. and Seibert, M., “Sustained Photobiological Hydrogen gas Production Upon Reversible Inactivation of Oxygen Evolution in the Green Alga Chlamydomonas reinhardtii,”Plant Physiol., 122(1), 127-135 (2000).
18. Hallenbeck, P. C. and Benemann, J. R., “Biological Hydrogen Production; Fundamentals and Limiting Processes,”Int. J. Hydrogen Energy, 27(11-12), 1185-1193(2002).
19. Laurinavichene, T. V., Tolstygina, I. V., Galiulina, R. R., Ghirardi, M.
L., Seibert, M. and Tsygankov, A. A., “Dilution Methods to Deprive Chlamydomonas reinhardtii Cultures of Sulfur for Sub- sequent Hydrogen Photoproduction,”Int. J. Hydrogen Energy, 27(11-12), 1245-1249(2002).
20. Tsygankova, A., Kosourova, S., Seibertb, M. and Ghirardi, M.
L., “Hydrogen Photoproduction Under Continuous Illumination by Sulfur-Deprived, Synchronous Chlamydomonas reinhardtii Cultures,”Int. J. Hydrogen Energy, 27(11-12), 1239-1244(2002).
21. Flynn, T., Ghirardi, M. L. and Seibert, M., “Accumulation of O2- Tolerant Phenotypes in H2-Producing Strains of Chlamydomo- nas reinhardtii by Sequential Applications of Chemical Mutage- nesis and Selection,”Int. J. Hydrogen Energy, 27(11-12), 1421- 1430(2002).
22. Janssen, M. and Hoekema, S., “Biological Hydrogen Produc- tion, 2003,” available from: www.ftns.wau.nl/prock/Research/Rene/
Photobacteria.htm, accessed 7 April(2004).
23. Lindblad, P., Christensson, K., Lindberg, P., Fedorov, A., Pinto, F. and Tsygankov, A., “Photoproduction of Hydrogen by Wild- type Anabaena PCC 7120 and a Hydrogen Uptake Deficient Mutant: from Laboratory Experiments to Outdoor Culture,”Int.
J. Hydrogen Energy, 27(11-12), 1271-1281(2002).
24. Masukawa, H., Mochimaru, M. and Sakurai, H., “Disruption of Uptake Hydrogenase Gene, but not of Bidirectional Hydroge- nase Gene, Leads to Enhanced Photobiological Hydrogen Pro- duction by the Nitrogen-Fixing Cyanobacterium Anabaena sp.
PCC 7120,”Appl. Microbiol. Biotechnol., 58(5), 618-624(2002).
25. Troshina, O., Serebryakova, L., Sheremetieva, M. and Lindblad, P., “Production of H2 by the Unicellular Cyanobacterium Gloeo-
capsa alpicola CALU 743 During Fermentation,”Int. J. Hydro- gen Energy, 27(11-12), 1283-1289(2002).
26. Yoon, J. H., Sim, S. J., Kim, M. S. and Park, T. H., “High Cell Density Culture of Anabaena variabilis Using Repeated Injec- tions of Carbon Dioxide for the Production of Hydrogen,”Int. J.
Hydrogen Energy, 27(11-12), 1265-1270(2002).
27. Koku, H., Eroglu, I., Gündüz, U., Yücel, M. and Türker, L.,
“Kinetics of Biological Hydrogen Production by the Photosyn- thetic Bacterium Rhodobacter sphaeroides O.U. 001,”Int. J.
Hydrogen Energy, 28(4), 381-388(2003).
28. Ko, I. B. and Noike, T., “Use of Blue Optical Filters for Suppres- sion of Growth of Algae in Hydrogen Producing Non-Axenic Cultures of Rhodobacter sphaeroides RV,” Int. J. Hydrogen Energy, 27(11-12), 1297-1302(2002).
29. Lee, C. M., Chen, P. C., Wang, C. C. and Tung, Y. C., “Photo- hydrogen Production Using Purple Nonsulfur Bacteria with Hydrogen Fermentation Reactor Effluent,”Int. J. Hydrogen Energy, 27(11-12), 1309-1313(2002).
30. Maness, P. C. and Weaver, P. F., “Hydrogen Production from a Carbonmonoxide Oxidation Pathway in Rubrivivax Gelatino- sus,”Int. J. Hydrogen Energy, 27(11-12), 1407-1411(2002).
31. Singh, A., Pandey, K. D. and Dubey, R. S., “Enhanced Hydro- gen Production by Coupled System of Halobacterium Halobium and Chloroplast after Entrapment Within Reverse Micelles,”Int.
J. Hydrogen Energy, 24(8), 693-698(1999).
32. Kondo, T., Arakawa, M., Wakayama, T. and Miyake, J., “Hydro- gen Production by Combining two Types of Photosynthetic Bac- teria with Different Characteristics,”Int. J. Hydrogen Energy, 27(11-12), 1303-1308(2002).
33. Kirk, R. E., Othmer, D. F., Grayson, M. and Eckroth, D., “Con- cise Encyclopedia of Chemical Technology XIII”, NewYork, Wiley-Interscience, 838-893(1985).
34. Hart, D., “Hydrogen Power: the Commercial Future of the Ulti- mate Fuel,” London, Financial Times Energy Publishing(1997).
35. Tanisho, N., Kuromoto, M. and Kadokura, N., “Effect of CO2
Removal on Hydrogen Production by Fermentation,”Int. J.
Hydrogen Energy, 23(7), 559-563(1998).
36. Van Niel, E. W. J., Claassen, P. A. M. and Stams, A. J. M., “Sub- strate and Product Inhibition of Hydrogen Production by the Extreme Thermophile, Caldicellulosiruptor saccharolyticus,”
Biotechnol. Bioeng., 81(3), 255-262(2003).
37. Heyndrickx, M., Vos, P. D. and Ley, J. D., “Fermentation Char- acteristics of Clostridium pasteurianum LMG 3285 Grown on Glucose and Mannitol,” J. Appl. Bacteriol., 70, 52-58(1991).
38. Chen, W.-M., Tseng, Z.-J., Lee, K.-S. and Chang, J.-S., “Fer- mentative hydrogen production with clostridium butylicum cgs5 isolated From Anaerobic Sewage Sludge,”Int. J. Hydrogen Energy, in press, available online at www.sciencediret.com(2004).
39. Saint-Amans, S., Girbal, L., Andrade, J., Ahrens, K. and Soucaille, P.,
“Regulation of Carbon and Electron Cow in Clostridium butyri- cum VPI 3266 Grown on Glucose Glycerol Mixtures,”J. Bacte- riol., 183(5), 1748-1754(2001).
40. Christophe, C., Nevenka, A., Jean-Paul, S., Paul, P., “Hydrogen Production by Clostridium thermolacticum During Continuous Fermentation of Lactose,”Int. J. Hydrogen Energy, 29(14), 1479-
1485(2004).
41. Rachman, M. A., Furutani, Y., Nakashimada, Y. and Kakizono, T. and Nishio, N., “Enhanced Hydrogen Production in Altered Mixed Acid Fermentation of Glucose by Enterobacter aero- genes,”J. Fermen. Bioeng., 83(4), 358-363(1997).
42. Yokoi, H., Ohkawara, T., Hirose, J., Hayashi, S. and Takasaki, Y., “Characteristics of Hydrogen Production by Aciduric Enter- obacter aerogenes Strain HO-39,”J. Fermen. Bioeng., 80(6), 571-574(1995).
43. Kumar, N. and Das, D., “Enhancement of Hydrogen Production by Enterobacter cloacae IIT-BT 08,”Process Biochem., 35(6), 589-593(2000).
44. Rachman, M. A., Furutani, Y., Nakashimada, Y., Kakizono, T.
and Nishio, N., “Enhanced Hydrogen Production in Altered Mixed Acid Fermentation of Glucose by Enterobacter aerogenes,”J. Fer- men. Bioeng., 83(4), 358-363(1997).
45. Kumar, N. and Das, D., “Continuous Hydrogen Production by Immobilized Enterobacter cloacae IIT-BT 08 Using Lignocellu- losic Materials as Solid Matrices,”Enzyme Microb. Technol., 29(4-5), 280-287(2001).
46. Palazzi, E., Fabiano, B. and Perego, P., “Process Development of Continuous Hydrogen Production by Enterobacter aerogenes in a Packed Column Reactor,”Bioprocess Eng., 22, 205-213(2000).
47. Godfroy, A., Raven, N. D. H. and Sharp, R. J., “Physiology and Continuous Culture of the Hyperthermophilic Deep-sea Vent Archaeon Pyrococcus abyssi ST549,”FEMS Microbiol. Lett., 186(1), 127-132(2000).
48. Schröder, C., Selig, M. and Schönheit, P., “Glucose Fermenta- tion to Acetate, CO2, and H2 in the Anaerobic Hyperthermo- philic Eubacterium Thermotoga maritima: Involvement of the Embden-Meyerhof Pathway,”Arch Microbiol., 161(6), 460-470 (1994).
49. Fiala, G. and Stetter, K. O., “Pyrococcus furiosussp. nov. Rep- resents a Novel Genus of Marine Heterotrophic Archaebacteria Growing Optimally at 100oC,”Arch Microbiol., 145(1), 56-61(1986).
50. Dietrich, G., Weiss, N. and Winter, J., “Acetothermus paucivorans, gen. nov., sp. nov, a Strictly Anaerobic, Thermophilic Bacterium From Sewage Sludge, Fermenting Hexoses to Acetate, CO2, and H2,”Syst. Appl. Microbiol., 10, 174-179(1988).
51. Soutschek, B., Winter, J., Schindler, F. and Kandler, O., “Ace- tomicrobium flavidum, gen. nov., sp., nov., a Thermophilic, Anaer- obic Bacterium from Sewage Sludge, Forming Acetate, CO2, and H2 From Glucose,”Syst. Appl. Microbiol., 5, 377-390(1984).
52. Reith, J. H., Wijffels, R. H. and Barten, H. (Ed.), Bio-Methane &
bio-Hydrogen: Status and Perspectives of Biological Methane and Hydrogen Production, Dutch Biological Hydrogen Founda- tion, The Netherlands, 103-123(2003).
53. Van Niel, E. W. J., Budde, M. A. W., De Haas, G. G., Van der Wal, F. J., Claassen, P. A. M. and Stams, A. J. M., “Distinctive Properties of High Hydrogen Producing Extreme Thermophiles, Caldicellulosiruptor saccharolyticus and Thermotoga elfii,”Int.
J. Hydrogen Energy, 27(11-12), 1391-1398(2002).
54. Kanai, T., Fukui, T., Atomi, H. and Imanaka, T., “Continuous Hydrogen Production by the Hyperthermophilic Archaeon, Ther- mococcus kodakaraensis KOD1,” 15th WHEC, June, Japan(2004).