Corresponding author: Chang-Ju Kim, Department of Physiology, College of Medicine, Kyung Hee University, 1, Hoigi-dong, Dong- daemoon-gu, Seoul, 130-701, Korea
Tel: +82-2-961-0407, E-mail: [email protected] Received November 24, 2010, Revised December 5, 2010 Accepted December 10, 2010
Protective Effect of Codonopsis pilosula (Franch) Nannf on Intracerebral Hemorrhage-Induced Memory Deficit and
Apoptotic Neuronal Cell Death in Rats
*Department of Physiology, College of Medicine, Kyung Hee University, Seoul,
†Department of Ophthalmology, Chungju Hospital, Konkuk University College of Medicine, Chungju, Korea
Dae-Young Kim*, Il-Gyu Ko*, Bo-Kyun Kim*, Sung-Eun Kim*, Shin-Ho Lee*, Young-Ho Jin*, Dong-Hee Kim†, Chang-Ju Kim*
Intracerebral hemorrhage (ICH) is a common cause of stroke, and it occurs mainly in the striatum, thalamus, cerebellum, and pons. Codonopsis pilosula (Franch) Nannf (Dangshen) is the roots of a perennial herb, the same family as ginseng. It comprises a rich resource of neuroactive compounds, and they are potential signaling molecules. In the present study, we investigated the effects of the Codonopsis pilosula intrastriatal on hemorrhage-induced memory deficit and apoptotic neuronal cell death in rats. ICH in the striatum was induced by injection of collagenase using strereotaxic instrument. The rats in the Codonopsis pilosula treated groups received Codonopsis pilosula orally once a day for 14 consecutive days, starting one day after surgery. For this study, step-down avoidance task, radial 8-arm maze task, terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) staining, immunohistochemistry for caspase-3, Western blot for Bax and Bcl-2 were performed. The present results showed that cognitive incapacity by ICH was associated with the increase of apoptosis in the striatum. Codonopsis pilosula treatment alleviated the ICH-induced impairment of spatial learning ability and short-term memory by suppressing neuronal apoptosis in the striatum. Here in this study, we suggest that Codonopsis pilosula is effective for the recovery after ICH insult. (Korean J Str Res 2010;18:305∼
314)
Key Words: Intracerebral hemorrhage, Codonopsis pilosula, Apoptosis, Spatial learning ability, Short-term memory
INTRODUCTION
Hemorrhagic stroke occurs when a blood vessel bursts inside the brain (Wang et al., 2007). There are two main subtypes of
hemorrhagic stroke: intracerebral hemorrhage (ICH) and subara- chnoid hemorrhage. Of these, ICH is a common cause of stroke, occurring mainly in the striatum, thalamus, cerebellum, and pons (Thrift et al., 1995). ICH is associated with severe neurological deficits and it has a considerable mortality rate. Brain damage by ICH occurs through multiple mechanisms such as direct tissue destruction, space-occupying effect of hematoma, ischemia damage to adjacent tissues, clot-derived toxic factors, edema formation (Lee KR et al., 1997), and ultimately it causes neuronal cell death (Mendelow, 1993). ICH induces brain cell death through apoptosis.
Apoptosis, also known as programmed cell death, is a form of cell death that serves to eliminate dying cells in proliferating or differentiating cell populations (Kerr et al., 1972; Gong et al., 2001), thus apoptosis plays a crucial role in normal development and tissue homeostasis (Woodle et al., 1998). On the other hand, inappropriate or excessive apoptosis has been implicated in stroke and several types of neurodegenerative disorders (Johnson et al., 1995). Apoptosis is an important from of ICH-induced cell death.
In the previous study, apoptosis is implicated in ICH-induced neuronal cell death in rats (Del Bigio et al., 2001). In addition, it was also reported that apoptotic cell death significantly contributes to the neuronal cell loss in the striatum following ICH in rats (Gong et al., 2001; Lee HH et al., 2005; Jung KH et al., 2007; Yang YJ et al., 2007).
Codonopsis pilosula (Franch) Nannf (Dangshen) is the root of a perennial herb, the same family as ginseng. Codonopsis pilosula possesses saponin, 5-hydroxymethyl-2-furaldehyde, alkaloid, Vita- min B1, B2, and others. Codonopsis pilosula exerts anti-aging effect and has curing effects on various diseases (Jin et al., 1996).
Previous studies reported that Codonopsis pilosula comprises a rich resource of neuroactive compounds, and they are the potential signaling molecules (Greene et al., 1986; Xia et al., 1995).
Codonopsis pilosula has been demonstrated that it is helpful to cure the animal model of remembering dysfunction (Pan, 1992).
However, although the functional roles of Codonopsis pilosula have been suggested, its effects on ICH in relation with memory are still unknown.
In the present study, we investigated the effects of the Codonopsis pilosula on intrastriatal hemorrhage-induced memory deficit and apoptotic neuronal cell death in rats. For this study, step-down avoidance task, radial 8-arm maze task, terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) staining, immunohistochemistry for caspase-3, Western blot for Bax and Bcl-2 were performed.
MATERIALS AND METHODS
1. Animals and treatments
Adult male Sprague-Dawley rats, weighing 300±10 g (10 weeks old), were obtained from a commercial breeder (Orient Co., Seoul, Korea) for the experiment. The experimental procedures
were performed in accordance with the animal care guidelines of the National Institutes of Health (NIH) and the Korean Academy of Medical Sciences. Each animal was housed under controlled temperature (20±2oC) and lighting (08:00∼20:00 h) conditions with food and water made available ad libitum before and after surgery.
The animals were randomly divided into five groups (n=8 in each group): the sham-operation group, the ICH-induced group, the ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, the ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, and the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group.
The rats in the Codonopsis pilosula-treated group received Codonopsis pilosula orally once a day for 14 consecutive days, starting one day after surgery. The rats in the sham-operation group and in the ICH-induced group received an equal amount of distilled water for the same duration.
2. Induction of ICH
For the induction of hemorrhage in the striatum, the rats were anesthetized with Zoletil 50Ⓡ (10 mg/kg, i.p.; Vibac Laboratories, Carros, France) and placed in a stereotaxic frame. Through a hole drilled in the skull, a 26-gauge needle was implanted into the striatum at the following coordinates: 2.6 mm lateral to the midline, 0.7 mm anterior to the coronal suture, and depth 4.5 mm deep from the surface of the brain. One μl of saline containing 0.2 U collagenase (Type 4; Sigma Chemical Co., St Louis, MO, USA) was infused over 1 min. The needle remained in place for an additional 2 min after the infusion and subsequently was withdrawn slowly.
3. Preparation of aqueous extract of Codonopsis pilosula
Codonopsis pilosula used in this experiment was obtained from Kyung-Dong Market (Seoul, Korea). After washing, Codonopsis pilosula was immersed in cold water for 1 hour. To obtain the aqueous extract of Codonopsis pilosula, 200 g of Codonopsis pilosula was added to distilled water, heat-extracted at 100oC, concen- trated with a rotary evaporator and lyophilization (EYELA, Tokyo, Japan). The resulting powder, weighing 25 g, was then diluted with saline.
4. Step-down avoidance task
The latency of the step-down avoidance task was determined to evaluate the short-term memory capability, according to the presiously described method (Lee HH et al., 2006). The rats were trained in a step-down avoidance task 11 days after induction of ICH. Two hours after training, the latency (sec) of the animals in each group was determined.
The rats were placed on a 7×25 cm platform with 2.5 cm height. The platform faced a 42×25 cm grid of parallel 0.1 cm-caliber stainless steel bars spaced 1 cm apart. In training sessions, the animals received 0.5 mA, scramble foot shock for 2 sec immediately upon stepping down. The interval of rats stepping down and placing all four paws on the grid was defined as the latency time of step-down avoidance task. In the step- down avoidance task, the latency was determined and latency over 180 sec was counted as 180 sec.
5. Radial 8-arm maze test
The spatial learning ability was tested using a radial 8-arm maze apparatus as the previously described method (Kim H et al., 2006). The radial-arm maze apparatus consisted of a central octagonal plate (30 cm in diameter) and radiating eight arms (50 cm in length and 10 cm in width). The apparatus was placed 1 m above the floor. A small receptacle filled with water (3 cm in diameter and 1 cm in depth) was located at the end of the arms.
The rats were trained three times before the spatial learning test.
The rats deprived of water for 24 h were allowed to explore for water and to drink water for 5 min. Test was conducted on the 13th day following ICH induction or sham operation. The time spent for the seeking of water at the end of the arms was counted. The test was terminated when a rat found water in all eight arms or over 5 min elapsed. Re-entering to the previously visited arm was counted as error. In addition, the number of correct choice before the first error was counted.
6. Tissue preparation
The animals were sacrificed on the 14th day following ICH induction or sham operation. The animals were anesthetized using Zoletil 50Ⓡ (10 mg/kg, i.p.; Vibac Laboratories), transcardially perfused with 50 mM phosphate-buffered saline (PBS), and fixed
with a freshly prepared solution consisting of 4% parafor- maldehyde in 100 mM phosphate buffer (PB, pH 7.4). The brains were dissected and postfixed in the same fixative overnight and transferred into a 30% sucrose solution for cryoprotection.
Coronal sections of 40 μm thickness were made with a freezing microtome (Leica, Nussloch, Germany). Six to eight sections on average were selected from each brain in the region spanning from Bregma 0.70 to 0.48 mm for striatum.
7. TUNEL staining
For visualization of DNA fragmentation, a marker of apoptotic cell death, TUNEL staining was performed using In Situ Cell Death Detection KitⓇ (Roche, Mannheim, Germany) according to the manufacturer’s protocol (Sim YJ et al., 2004). To begin the procedure, the sections were post-fixed in ethanol-acetic acid (2 : 1) and rinsed. Then, the sections were incubated with proteinase K (100 μg/ml), rinsed, incubated in 3% H2O2, permeabilized with 0.5% Triton X-100, rinsed again, and incubated in TUNEL reaction mixture. The sections were rinsed and visualized using Converter-POD with 0.03% 3,3’-diaminobenzidine (DAB).
Mayer’s hematoxylin (DAKO, Glostrup, Denmark) was used for counter-staining, and the sections were finally mounted onto gelatin-coated slides. The slides were air dried overnight at room temperature, and coverslips were mounted using PermountⓇ.
8. Caspase-3 immunohistochemstry
For visualization of caspase-3 expression, caspase-3 immunohis- tochemistry was performed as previously described (Yang YJ et al., 2007). The sections were drawn from each brain and incubated overnight with mouse anti-caspase-3 antibody (1:500; Santa Cruz Biotechnology, Santa Cruz, CA, USA) and then for another 1 h with biotinylated mouse secondary antibody. Bound secondary antibody was then amplified with Vector Elite ABC kitⓇ (Vector Laboratories, Burlingame, CA, USA). The antibody-biotin-avidin- peroxidase complexes were visualized using 0.03% DAB and the sections were finally mounted onto gelatin-coated slides. The slides were air dried overnight at room temperature, and coverslips were mounted using PermountⓇ.
9. Western blot analysis for Bax and Bcl-2 The striatum tissues were collected, and then were immediately
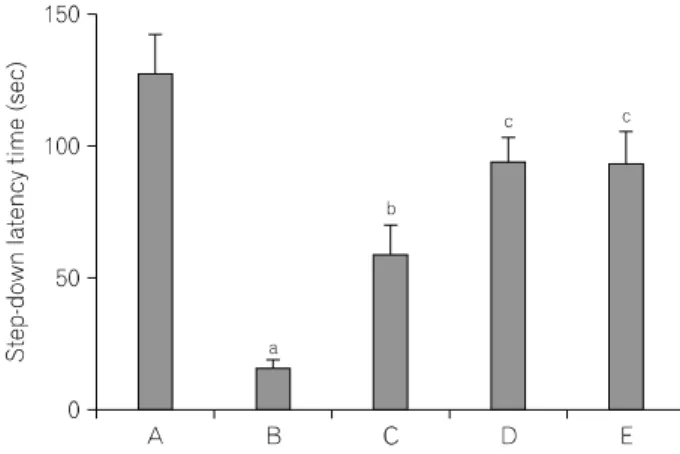
Fig. 1. Effect of Codonopsis pilosula on latency of a step-down avoidance task. (A) Sham-operation group, (B) ICH-induced group, (C) ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, (D) ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, (E) ICH-induced and 200 mg/kg Codonopsis pilosula-treated group. aRepresents p<0.05 compared to the sham-operation group. bRepresents p<0.05 compared to the ICH-induced group. cRepresents p<0.05 compared to the ICH-induced and 50 mg/kg Codonopsis pilosula-treated group.
frozen at −70oC. The tissues were homogenized with lysis buffer containing 50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 10%
glycerol, 1% Triton X-100, 1.5 mM MgCl2·6H2O, 1 mM EGTA, 1 mM PMSF, 1 mM Na2VO4, and 100 mM NaF, and then centrifuged at 14,000 rpm for 30 min. Protein content was measured using a Bio-Rad colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Protein of 30 μg was separated on SDS-polyacrylamide gels and transferred onto a nitrocellulose membrane. Mouse actin antibody (1:1,000; Santa Cruz Biotech, CA, USA), mouse Bax antibody (1:1,000; Santa Cruz Biotech) and mouse Bcl-2 antibody (1:1,000; Santa Cruz Biotech) were used as primary antibodies. Horseradish peroxidase-conjugated anti-mouse antibody for Bax and Bcl-2 (1:2,000; Amersham Pharmacia Biothech GmbH, Freiburg, Germany) were used as secondary antibodies. Experiment was performed in normal lab conditions and at room temperature except membrane transfer.
Membrane transfer was performed at 4oC with the cold pack and prechilled buffer. Band detection was performed using the enhanced chemiluminescence (ECL) detection kit (Santa Cruz Biotech).
10. Date analysis
To compare relative expression of proteins, detected bands were calculated densitometrically using Molecular AnalystTM version 1.4.1 (Bio-Rad). The numbers of TUNEL-positive and caspase-3 positive cells were assessed quantitatively in the four filededs (250×250 μm) within the striatum adjacent to the hematoma as the previously described method (Yang YJ et al., 2007). Statistical analysis was performed using one-way ANOVA followed by Duncan's post-hoc test, and the results were expressed as the mean±standard error of the mean (S.E.M). Significance was accepted at p<0.05.
RESULTS
1. Effect of Codonopsis pilosula on the step- down avoidance task
The latencies of the step-down avoidance task are presented in Fig. 1. The latency time was 129.90±14.61 sec in the sham- operation group, 18.41±2.99 sec in the ICH-induced group, 61.13±11.33 sec in the ICH-induced and 50 mg/kg Codonopsis
pilosula-treated group, 96.50±9.58 sec in the ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, and 96.50±12.45 sec in the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group, respectively.
These results showed that short-term memory was disturbed by induction of ICH, and Codonopsis pilosula treatment alleviated ICH-induced short-term memory impairment.
2. Effect of Codonopsis pilosula on spatial learning ability
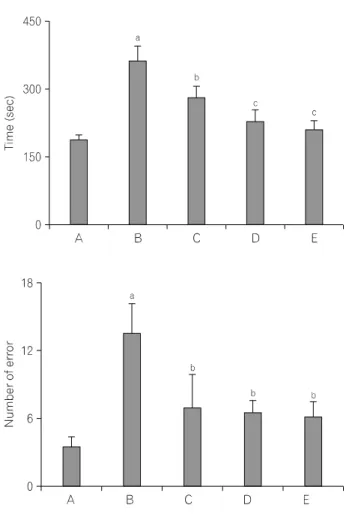
The time of successful performance, the number of the correct, and the number of error choice in the radial 8-maze task are presented in Fig. 2. The time in the completed eight successful performances was 187.51±10.91 sec in the sham-operation group, 367.12±31.21 sec in the ICH-induced group, 282.16±24.97 sec in the ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, 232.56±21.44 sec in the ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, and 212.51±17.66 sec in the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group, respectively.
The number of correct choice before the first error was 6.33±0.41 in the sham-operation group, 4.67±0.61 in the ICH- induced group, 5.00±0.54 in the ICH-induced and 50 mg/kg
Fig. 2. Effect of Codonopsis pilosula on performance time, correct number, and error score of the radial 8-arm maze task. (A) Sham-operation group, (B) ICH-induced group, (C) ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, (D) ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, (E) ICH-induced and 200 mg/kg Codonopsis pilosula-treated group. aRepresents p<0.05 compared to the sham-operation group. bRepresents p<0.05 compared to the ICH-induced group.
cRepresents p<0.05 compared to the ICH-induced and 50 mg/kg Codonopsis pilosula-treated group.
Codonopsis pilosula-treated group, 6.00±0.48 in the ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, and 6.00±0.59 in the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group, respectively.
The number of error made before eight successful performances was 3.67±0.99 in the sham-operation group, 13.80±2.56 in the ICH-induced group, 7.17±2.96 in the ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, 6.60±1.11 in the ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, and 6.25±1.38 in the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group, respectively.
These results demonstrated that ICH-induction in rats showed increased time to seek water and lower number of correct choice and higher number of error. Treatment with Codonopsis pilosula showed shorted time to seek water, increased number of correct choice, and decreased error number.
3. Effect of Codonopsis pilosula on TUNEL- positive cells in the striatum
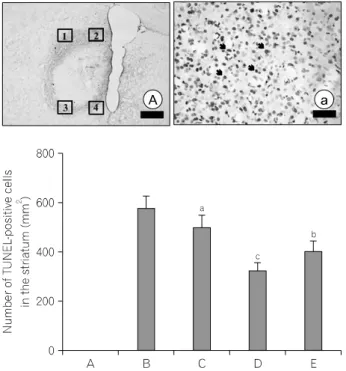
Photomicrographs of TUNEL-positive cells are shown in Fig. 3.
The number of TUNEL- positive cells was nearly zero in the sham-operation group, 589.69±48.33/mm2 in the ICH-induced group, 499.31±59.25/mm2 in the ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, 332.12±31.97/mm2 in the ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, and 411.23±41.33/mm2 in the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group, respectively.
The present results showed that hemorrhagic insult enhanced DNA fragmentation in the striatum and that Codonopsis pilosula treatment significantly suppressed the ICH-induced DNA fragmentation. The number of TUNEL-positive cells in the 100 mg/kg Codonopsis pilosula-treated group was most potently suppressed compared to the other groups.
Fig. 3. Effect of Codonopsis pilosula on DNA fragmentation in the striatum induced by intracerebral hemorrhage. Upper: Photomicrographs of terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL)-positive cells in the striatum. The number of TUNEL-positive cells in the field of 1, 2, 3, and 4 were counted. The sections were stained for TUNEL (arrow). The scale bar represents 800 mm (A) and 100 mm (a). Lower: Number of TUNEL-positive cells in each group. (A) Sham-operation group, (B) ICH-induced group, (C) ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, (D) ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, (E) ICH-induced and 200 mg/kg Codonopsis pilosula-treated group. aRepresents p<0.05 compared to the sham-operation group. bRepresents p<0.05 compared to the ICH- induced group. cRepresents p<0.05 compared to the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group.
Fig. 4. Effect of Codonopsis pilosula on caspase-3 in the striatum induced by intracerebral hemorrhage. Upper: Photomicrographs of caspase-3- positive cells in the striatum. The number of caspase-3-positive cells in the field of 1, 2, 3, and 4 were counted. The sections were stained for caspase-3 like immunoreactivity. The scale bar represents 800 mm (A) and 100 mm (a). Lower: Number of caspase-3-positive cells in each group. (A) Sham-operation group, (B) ICH-induced group, (C) ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, (D) ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, (E) ICH-induced and 200 mg/kg Codonopsis pilosula-treated group. bRepresents p<0.05 compared to the ICH-induced group. cRepresents p<0.05 compared to the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group.
4. Effect of Codonopsis pilosula on caspase-3 expression in the striatum
Photomicrographs of caspase-3 positive cells are shown in Fig. 4.
The number of caspase-3 positive cells was nearly zero in the sham-operation group, 444.31±67.95/mm2 in the ICH-induced group, 421.39±51.45/mm2 in the ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, 265.46±33.21/mm2 in the ICH- induced and 100 mg/kg Codonopsis pilosula-treated group, and 339.56±29.38/mm2 in the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group, respectively.
The present results showed that hemorrhagic insult enhanced caspase-3 expression in the striatum and that Codonopsis pilosula
treatment significantly suppressed the ICH-induced caspase-3 expression. The number of caspase-3 positive cells in the 100 mg/kg Codonopsis pilosula-treated group was mostpotently suppressed compared to the other groups.
5. Western blot analysis of Bax and Bcl-2 proteins
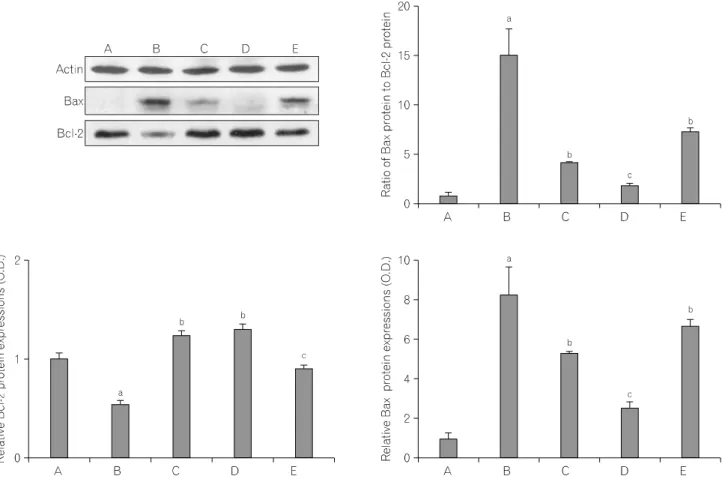
We ascertained the relative expressions of Bax and Bcl-2 proteins in the striatum. There were significant differences among groups (Fig. 5). When the level of Bax (23 kDa) in the sham-operation group was set as 1.00, the levels of Bax were 8.24±1.43 in the ICH-induced group, 5.27±0.10 in the ICH-induced and 50 mg/kg Codonopsis pilosula-treated group,
Fig. 5. Effect of Codonopsis pilosula on the expression of Bax and Bcl-2 proteins. (A) Sham group, (B) ICH-induced group, (C) ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, (D) ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, (E) ICH-induced and 200 mg/kg Codonopsis pilosula-treated group. aRepresents p<0.05 compared to the sham-operation group. bRepresents p<0.05 compared to the ICH-induced group. cRepresents p<0.05 compared to the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group (left and right panels).
2.56±0.28 in the ICH-induced and 100 mg/kg Codonopsis pilosula- treated group, and 6.74±0.28 in the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group, respectively.
ICH-induction decreased Bax expression. In contrast, Codonopsis pilosula administration reduced the expression of Bax. Treatment with 100 mg/kg Codonopsis pilosula most potently suppressed the expression of Bax protein.
When the level of Bcl-2 (26∼29 kDa) in the sham group was set as 1.00, the levels of Bcl-2 were 0.55±0.04 in the ICH- induced group, 1.24±0.06 the ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, 1.29±0.06 the ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, and 0.90±0.05 in the ICH-induced and 200 mg/kg Codonopsis pilosula-treated group, respectively.
ICH-induction decreased Bcl-2 expression. In contrast, Codonopsis
pilosula administration increased the expression of Bcl-2. Treatment with 100 mg/kg Codonopsis pilosulamost potently increased the expression of Bcl-2 protein.
The Bax/Bcl-2 ratio, one of the crucial factors related to apoptosis, was calculated. When the ratio in the sham-operation group was set as 1.00, the Bax/Bcl-2 ratio were 15.00±2.60 in the ICH-induced group, 4.26±0.08 in the ICH-induced and 50 mg/kg Codonopsis pilosula-treated group, 1.98±0.21 in the ICH-induced and 100 mg/kg Codonopsis pilosula-treated group, and 7.45±0.31 in the ICH-induced and 200 mg/kg Codonopsis pilosula- treated group, respectively.
Administration of Codonopsis pilosula significantly decreased the expression of Bax protein and increased expression of Bcl-2 protein. As a result, the Bax/Bcl-2 ratio was decreased by Codonopsis pilosula treatment. Treatment with 100 mg/kg
Codonopsis pilosula most potently suppressed the Bax/Bcl-2 ratio.
DISCUSSION
ICH induces neuronal cell death through complex mechanisms, and it causes memory impairment. The striatum is the main input structure of the basal ganglia, and receives input from all major areas of neocortex as well as the hippocampal formation and pyriform cortex (McGeorge et al., 1989; Groenewegen et al., 1994). The striatum also plays a critical role in cognitive flexibility (Cools et al., 2003), learning, and performance of tasks (Atallah et al., 2007). Damage to the striatum can result in deficits of motor, cognitive, associative, and limbic functions (Benke et al., 2003; GilRobles et al., 2005; Muslimovic et al., 2005).
In the present study, the results of the behavior test revealed that ICH incapacitated short-term and spatial memory abilities, indicating that ICH-induction in the striatum impaired the rats’
capacity for short-term and spatial learning memory.
Apoptosis appears to play a key role in neuronal cell death in stroke (Matsushita et al., 2000; Qureshi et al., 2003). Especially, apoptosis may contribute to brain damage after ICH in rat striatum (Matsushita et al., 2000). The morphological characteristics of apoptosis including cell shrinkage, chromatin condensation, membrane blebbing, internucleosomal DNA fragmentation, and formation of apoptotic bodies have been reported in cells undergoing ischemic death (Li et al., 1995). TUNEL staining is a widely used method for the detection of DNA fragmentation, a characteristic of apoptotic cell death (Gavrieli et al., 1992; Lee HH et al., 2005). In the ICH animal models, TUNEL-positive cells represent apoptotic neuronal cell death (Lee HH et al., 2005). Caspase-3 is one of the most widely studied members of the caspase family, and it is one of the key executors of apoptosis (Cohen, 1997). Up-regulation and activation of caspase-3 at the early stage of apoptosis following ischemia and hemorrhage have been reported (Benchoua et al., 2001; Sim YJ et al., 2004).
In the present results, the numbers of TUNEL-positive and caspase-3-positive cells in the striatum were significantly increased following ICH attack, indicating that ICH-induced apoptotic neuronal cell death in the striatum.
The apoptotic common pathway involves molecules such as
caspases, mitochondrial factors, and the Bcl-2 family of proteins (Benn et al., 2004; Danial et al., 2004). These proteins have been proposed to act as the critical determinant in the regulation of cell death, and they are divided into anti-apoptotic protein and pro-apoptotic protein (Cheng et al., 2006). Bcl-2 can inhibit apoptosis by preventing the release of cytochrome-c from mitochondria. However, Bcl-2 and Bcl-xL form heterodimers with the main pro-apoptotic member Bax, and it can be incapacitated in their protective function (Reed et al., 1998; Kuwana et al., 2003). That is the reason that the Bax and Bcl-2 balance is used one of the crucial factors determining whether cells undergo apoptosis.
Our present study showed that the expression of Bax was increased and the expression of Bcl-2 was decreased in the striatum after ICH attack. These results revealed that induction of ICH accelerated apoptotic neuronal cell death in the striatum.
Codonopsis pilosula resembles ginseng and the therapeutic properties are similar, but its actions are not strong as ginseng and it is not so expensive. Codonopsis pilosula had been demonstrated helpful to cure the memory impairment in animal models (Pan, 1992). Codonopsis pilosula is known to contain saponin. Saponin is well known potent neuroprotective effect agent (Kim YC et al., 1998). Especially, the anti-apoptotic effect of saponin on neurons was well proved in vivo and in vitro experiments (Chen et al., 2002). Li et al.(2009) reported that total saponins markedly reduced the numbers of TUNEL-positive and caspase-3-positive cells in brain after stroke. This study indicated that saponin could attenuate neuronal apoptosis induced by stroke and promote neuronal survival via inhibiting caspase-3 activation.
Saponin is also known to improve learning ability and memory capability in many disease models and senile dementia (Chen et al., 2001; Wang et al., 2001; Cheng et al., 2005; Zhang et al., 2008).
Our present study showed that Codonopsis pilosula significantly suppressed both ICH-induced increase in DNA fragmentation and caspase-3 expression in the striatum. In addition, Codonopsis pilosula suppressed the Bax/Bcl-2 ratio, showing that ICH-induced apoptosis was inhibited by Codonopsis pilosula treatment. These results suggest that certain ingredients of Codonopsis pilosula inhibited neuronal apoptosis through up-regulation of Bcl-2 and down-regulation of Bax. This inhibiting effect on apoptosis of
Codonopsis pilosula was most potently appeared at 100 mg/kg.
Our study also revealed that ICH-related increased of apoptosis in the striatum is closed associated with ICH-induced disturbance of cognitive function. Codonopsis pilosula treatment ameliorated ICH-induced decline of cognitive functions very effectively by suppressing apoptosis. Here in this study, we suggest that Codonopsis pilosula treatment can overcome ICH-induced neuronal apoptosis, thus facilitates recovery following ICH.
REFERENCES
Atallah HE, Lopez-Paniagua D, Rudy JW et al. (2007) Separate neural substrates for skill learning and performance in the ventral and dorsal striatum. Nat. Neurosci. 10:126-131.
Benchoua A, Guegan C, Couriaud C et al. (2001) Specific caspase pathways are activated in the two stages of cerebral infarction.
J. Neurosci. 21:7127-7134.
Benke T, Delaze M, Bartha L et al. (2003) Basal ganglia lesions and the theory of fronto-subcortical loops: Neuropsychological findings in two patients with left caudate lesions. Neurocase 9:70-85.
Benn SC, Woolf CJ (2004) Adult neuron survival strategies-slamming on the brakes. Nat. Rev. Neurosci. 9:686-700.
Chen J, Gong YS, Zhang JT (2001) Effects of 17 estradiol and total ginsenoside on the spatial learning and memory impairment of ovariectomy rats. Chin. Pharm. J. 36:522-526.
Chen XC, Chen Y, Zhu YG et al. (2002) Protecive effect of ginsenoside Rg1 against MPTP-induced apoptosis in mouse substantia nigra neurons. Acta Pharmacol. Sin. 23:829-834.
Cheng WC, Berman SB, Ivanovska I et al. (2006) Mitochondrial factors with dual roles in death and survival. Oncogene 25:4697- 4705.
Cheng Y, Shen LH, Zhang JT (2005) Anti-amnestic and anti-aging effects of ginsenoside Rg1 and Rb1 and its mechanism of action.
Acta Pharmacol. Sin. 26:143-149.
Cohen GM (1997) Caspases: the executioners of apoptosis. Biochem.
J. 326:1-16.
Cools R, Barker RA, Sahakian BJ et al. (2003) L-Dopa medication remediates cognitive inflexibility, but increases impulsivity in patients with Parkinson’s disease. Neuropsychologia 41:1431-1441.
Danial NN, Korsmeyer SJ (2004) Cell death: critical control points.
Cell 116:205-219.
Del Bigio MR, Yan HJ, Xue M (2001) Intracerebral infusion of a second-generation ciliary neurotrophic factor reduces neuronal loss in rat striatum following experimental intracerebral hemorrhage. J. Neurol. Sci. 192:53-59.
Gavrieli Y, Sherman Y, Ben-Sasson SA (1992) Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 119:493-501.
GilRobles S, Gatignol P, Capelle L et al. (2005) The role of dominant striatum in language: Astudy using intraoperative electrical stimulations. J. Neurol. Neurosurg. Psychiatry 76:940-946.
Gong C, Boulis N, Qian J et al. (2001) Intracerebral hemorrhage- induced neuronal death. Neurosurgery 48:875-882.
Greene LA, Aletta JM, Rukenstein A et al. (1986) PC12 pheochromocytoma cells: culture, nerve growth factor treatment, and experimental exploitation. Methods Enzymol. 147:207-216.
Groenewegen HJ, Berendse HW (1994) The specificity of the 'nonspecific' midline and intralaminar thalamic nuclei. Trends Neurosci. 17:52-57.
Jin ZL, Wen J, Li SF (1996) Antiaging effect of shenqi mixture tonic. Chin. Trad. Herb. Drugs 27:90-92.
Johnson EM Jr, Greenlund LJ, Akins PT et al. (1995) Neuronal apoptosis: current understanding of molecular mechanisms and potential role in ischemic brain injury. J. Neurotrauma 12:843- 852.
Jung KH, Chu K, Lee ST et al. (2007) Blockade of AT1 receptor reduces apoptosis, inflammation, and oxidative stress in normotensive rats with intracerebral hemorrhage. J. Pharmacol.
Exp. Ther. 322:1051-1058.
Kerr JF, Wyllie AH, Currie AR (1972) Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics.
Br. J. Cancer. 26:239-257.
Kim H, Lee MH, Chang HK et al. (2006) Influence of prenatal noise and music on the spatial memory and neurogenesis in the hippocampus of developing rats. Brain Dev. 28:109-114.
Kim YC, Kim SR, Markelonis GJ et al. (1998) Ginsenosides Rb1 and Rg3 protect cultured rat cortical cells from glutamate-induced neurodegeneration. J. Neurosci. Res. 53:426-432.
Kuwana T, Newmeyer DD (2003) Bcl-2-family proteins and the role of mitochondria in apoptosis. Curr. Opin. Cell Biol. 15:691-699.
Lee HH, Kim H, Lee JW et al. (2006) Maternal swimming during pregnancy enhances short-term memory and neurogenesis in the hippocampus of rat pups. Brain Dev. 28:147-154.
Lee HH, Shin MS, Kim YS et al. (2005) Early treadmill exercise decreases intrastriatal hemorrhage-induced neuronal cell death and increases cell proliferation in the dentate gyrus of strepto- zotocin-induced hyperglycemic rats. J. Diabetes Complications 19:339-346.
Lee KR, Kawai N, Kim S et al. (1997) Mechanisms of edema formation after intracerebral hemorrhage: effects of thrombin on cerebral blood flow, blood-brain barrier permeability, and cell survival in a rat model. J. Neurosurg. 86:272-278.
Li H, Deng CQ, Chen BY et al. (2009) Total saponins of Panax
= 국문초록 =
뇌내출혈은 선조체, 시상, 소뇌 그리고 교에서 주로 나타난다. 당삼(Codonopsis pilosula)은 풍부한 신경자극성 화합물을 포함하고 있는 생약제로서, 분자적 신호전달체계를 활성화 시키는 약효가 있는 것으로 알려져 있다. 본 연구에서는 뇌출혈에 의해 기억력 감소와 신경세포 사멸이 야기된 쥐에서 당삼의 효과를 규명하고자 하였다. 뇌출혈을 유발하 기 위해 stereotaxic instrument를 사용하여 선조체 부위에 collagenase를 주사하였다. 당삼 처치 그룹은 수술 다음 날부터 그룹별로 매일 14일 동안 당삼을 구강 투여하였다. 본 연구에서는 기억력 검사는 step-down avoidance task와 radial 8-arm maze task로 측정하였으며, 세포사멸은 terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) 염색과 caspase-3 면역조직화학법, 그리고 Bax와 Bcl-2의 발현을 Western blot으로 확인하였다. 본 실험의 결과 뇌출혈에 의해 공간학습 능력과 단기 기억이 감소되었고, 선조체에서 세포사멸이 증가되었다. 당삼의 처치는 뇌내출 혈에 의한 세포사멸을 억제시켰으며, 공간학습 능력과 단기 기억의 저하를 감소시켰다. 본 연구를 통하여 당삼은 뇌출혈 시 세포사멸을 억제하여 인지력의 감퇴를 억제하는 효과가 있음을 알 수 있었다.
중심단어: 뇌출혈, 당삼, 세포사멸사, 공간학습능력, 단기기억력 Notoginseng modulate the expression of caspases and attenuate
apoptosis in rats following focal cerebral ischemia-reperfusion. J.
Ethnopharmacol. 121:412-418.
Li Y, Chopp M, Jiang N et al. (1995) Induction of DNA fragmentation after 10 to 120 minutes of focal cerebral ischemia in rats. Stroke 26:1252-1257.
Matsushita K, Meng W, Wang X et al. (2000) Evidence for apoptosis after intercerebral hemorrhage in rat striatum. J. Cereb.
Blood Flow Metab. 20:396-404.
McGeorge AJ, Faull RL (1989) The organization of the projection from the cerebral cortex to the striatum in the rat. Neuroscience 29:503-537.
Mendelow AD (1993) Mechanisms of ischemic brain damage with intracerebral hemorrhage. Stroke 24:115-117.
Muslimovic D, Post B, Speelman JD et al. (2005) Cognitive profile of patients with newly diagnosed Parkinson disease. Neurology 65:1239-1245.
Pan SY (1992) Circadian effects of scopolamine on memory, exploratory behavior, and muscarinic receptors in mouse brain.
Acta Pharmacol. Sin. 13:323-326.
Qureshi AI, Ling GS, Khan J et al. (2003) Quantitative analysis of injured, necrotic, and apoptotic cells in a new experimental model of intracerebral hemorrhage. Crit. Care Med. 29:152-157.
Reed JC, Jurgensmeier JM, Matsuyama S (1998) Bcl-2 family proteins and mitochondria. Biochim. Biophys Acta 10:127-137.
Sim YJ, Kim SS, Kim JY et al. (2004) Treadmill exercise improves short-term memory by suppressing ischemia-induced apoptosis of neuronal cells in gerbils. Neurosci. Lett. 372:256-261.
Thrift AG, Donnan GA, McNeil JJ (1995) Epidemiology of intracerebral hemorrhage. Epidemiol. Rev. 17:361-381.
Wang J, Doré S (2007) Heme oxygenase-1 exacerbates early brain injury after intracerebral haemorrhage. Brain 130:1643-1652.
Wang XY, Zhang JT (2001) Effects of ginenoside Rg1 on learning and memory impairment induced by β-amyloid peptide (25-35) and its mechanism of action. Acta. Pharm. Sin. 36:1-4.
Woodle ES, Kulkarni S (1998) Programmed cell death.
Transplantation 66:681-691.
Xia Z, Dickens M, Raingeaud J et al. (1995) Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 270:
1326-1331.
Yang YJ, Kim YS, Shin MS et al. (2007) Effects of acupuncture on the intrastriatal hemorrhage-induced caspase-3 expression and newly cell birth in rats. Neurol. Res. 29:65-71.
Zhang G, Liu A, Zhou Y et al. (2008) Panax ginseng ginsenoside- Rg2 protects memory impairment via anti-apoptosis in a rat model with vascular dementia. J. Ethnopharmacol. 115:441-448.