Corresponding author: Sam-Jun Lee, Department of Physical Education, College of Health, Social Welfare, and Education, Tong Myong University, 428, Sinseollo, Nam-gu, Busan 608-711, Korea
Tel: +82-51-629-2106, E-mail: [email protected] Received April 5, 2011, Revised May 15, 2011
Accepted May 20, 2011
This work was supported by the National Research Foundation of Korea Grant funded by the Korea Government (NRF-2008-332-G00082).
Treadmill Exercise Reduces Lipopolysaccharide-induced Apoptotic Neuronal Cell Death in the Hippocampus of the Young and Old Rats
*Department of Exercise Physiology & Prescription, Graduate School of Health Promotion, Hanseo University, Seosan,
†
Department of Physiology, College of Medicine, Kyung Hee University, Seoul,
‡Department of Physical Education, College of Health, Social Welfare, and Education, Tong Myong University, Busan, Korea
Il-Gyu Ko*, Sung-Eun Kim
†, Mal-Soon Shin
†, Chang-Ju Kim
†, Sam-Jun Lee
‡Systemic inflammation exerts detrimental effects on the various organs, especially on the central nervous system (CNS), and leading to multiple organ failure. Lipopolysaccharide (LPS) is a lipid-containing polysaccharide which is endotoxin and acts as important group-specific antigen. LPS induces immune activation, causes inflammation, and results in deterioration of cellular function. Hippocampal neurons are particularly susceptible to this LPS. Treadmill exercise is known to ameliorate neurologic impairment induced by various brain insults. In the present study, we investigated the effects of treadmill exercise on short-term and spatial memories in relation with apoptotic neuronal cell death in the hippocampus following LPS-induced systemic inflammation. For the evaluation of age-dependent effect of treadmill exercise on these parameters, we used both young-aged and old-aged rats. In the present results, aging process impaired short-term and spatial memories through increase of apoptosis in the hippocampus. Treadmill exercise alleviated aging-induced impairment of short-term and spatial memories and suppressed aging-induced apoptosis in the old-aged rats. LPS-induced systemic inflammation disturbed short-term and spatial memories with increased apoptosis in both young-aged and old-aged rats. Treadmill exercise alleviated LPS-induced impairment of short-term and spatial memories and suppressed LPS-induced apoptosis in both young-aged and old-aged rats. Here in this study, we showed that treadmill exercise may inhibit LPS-induced neuronal apoptosis, thus facilitates recovery of memory function following systemic inflammation. Concerning that aging process disturbs memory function by enhancing of apoptotic neuronal cell death, these effects of treadmill exercise may be more important in the elderly. (Korean J Str Res 2011;19:97∼105)
Key Words: Lipopolysaccharide, Treadmill exercise, Hippocampus, Apoptosis, Memory
INTRODUCTION
Inflammation is the part of the complex biological responses of
various tissues to harmful stimuli, such as pathogens, damaged
cells, and irritants. Especially, a systemic inflammatory reaction
may have detrimental effects on the organs, including the central
nervous system (CNS), and leading to multiple organ failure and
sometimes death (Siami et al., 2008; Czapski et al., 2010). These
changes in the brain are implicated in the pathogenesis of chronic neurodegenerative disorders, such as Alzheimer’s disease and Parkinson’s disease (Ekdahl et al., 2003).
Systemic inflammation of brain is accompanied by neurode- generation, including necrotic and apoptotic neuronal cell death as well as destruction of neuritis connecting different neuronal populations (Trendelenburg, 2008). The previous studies indicated that mild systemic inflammation or septic shock induces long-lasting cognitive impairment through activation of apoptotic process and, resulting in neuronal loss (Semmler et al., 2007;
Jacewicz et al., 2009; Czapski et al., 2010).
Apoptosis, which is also known as programmed cell death, is a form of cell death that serves to eliminate dying cells in proliferating or differentiating cell populations (Kerr et al., 1972), thus apoptosis plays a crucial role in normal development and tissue homeostasis (Woodle et al., 1998). Nevertheless, inappro- priate or excessive apoptosis has been implicated in several neuro- goical disorders (Johnson et al., 1995; Lee MH et al., 2003; Ko IG et al., 2009). Terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) assay detects the characteristic of apoptotic cell death, DNA fragmentation (Gavrieli et al., 1992). Another important characteristic of apoptosis is caspases activation. Caspase-3 is one of the most widely studied caspases, and it is a key executor of apoptosis (Cohen, 1997). Lipopoly- saccharide (LPS) is a lipid-containing polysaccharide which is endotoxin and acts as important group-specific antigen. It is also derived from the cell wall of gram-negative bacteria and induces immunoglobulin secretion. LPS has widely been used for the induction of inflammation. Systemic inflammation by LPS treat- ment showed excessive apoptotic neuronal cell death in the various brain areas (Czapski et al., 2007). LPS treatment to the rats also induced electrophysiological, metabolic, and morpholo- gical changes with a decrease in the numbers of neuronal cells in the cortex and hippocampus (Czapski et al., 2010).
Physical exercise has been recommended for the prevention and treatment of many chronic brain diseases. Several studies reported that exercise improves cognitive function and delays the onset of Alzheimer’s disease (Rovio et al., 2005). In rodents, running exercise ameliorated various brain injury-induced neurological impairments and facilitates functional recovery by reducing neuronal loss (Carro et al., 2001). In addition, regular physical
exercise exerted anti-inflammatory effect on the various organs including brain (Aoi et al., 2010; Curry et al., 2010; Leem et al., 2011).
Many effects of exercise on brain function have been reported, however the effects of physical exercise on memory function in relation with apoptotic neuronal cell death following systemic inflammation remain uncertain. Moreover, age-dependence of the physical exercise on these effects has not been studied yet. In the present study, we investigated the effects of treadmill exercise on short-term and spatial memories in relation with apoptotic neuronal cell death in the hippocampus following LPS-induced systemic inflammation. For the evaluation of age-dependent effect of treadmill exercise on these parameters, we used both the old-aged rats and the young-aged rats. In this study, step-down inhibitory task for short-term memory, radial 8-arm maze task for spatial memory, and TUNEL assay with caspase-3 immuno- histochemistry for apoptosis were conducted.
MATERIALS AND METHODS
1. Experimental animals
Four-month old Sprague-Dawley rats (n=30; weighing 320±
10 g) were used as the young-aged group and 22-month old rats (n=30; weighing 360±15 g) were used as the old-aged group.
The experimental procedures were performed in accordance with the animal care guidelines of the National Institutes of Health (NIH) and the Korean Academy of Medical Sciences. Each animal was housed under controlled temperature (22±2
oC) and lighting (08 : 00 h∼20 : 00 h) conditions with food and water made available ad libitum.
The animals were randomly divided into six groups (n=10 in each group): young-aged control group, young-aged and LPS injection group, young-aged and LPS injection with treadmill exercise group, old-aged control group, old-aged and LPS injection group, old-aged and LPS injection with treadmill exercise group.
2. Induction of systemic inflammation
Systemic inflammation was induced by intraperitoneal injection
of 500 μg/kg LPS (055 : B5; Sigma-Aldrich Co., St. Louis, MO,
USA) on the 7, 14, 21, and 28 days after the starting of
experiment, while the rats in the control groups received 0.9%
saline intraperitoneally, for the same duration of time.
3. Treadmill exercise protocols
The rats in the exercise groups were forced to run on a motorized treadmill for 30 min once a day for 8 weeks after starting of experiment. The exercise load consisted of running at a speed of 2 m/min for the first 5 min, 5 m/min for the next 5 min, and 8 m/min for the last 20 min, with a 0
oinclination.
The rats in the control groups were left on the treadmill without running for the same period as the exercise groups.
4. Step-down avoidance task
The latency of the step-down avoidance task was determined for the evaluation of short-term memory. The rats were trained in a step-down avoidance task 5 days before sacrificed. One hour after training, the latency (sec) of the animals in each group was determined.
The rats were placed on a 7×25 cm platform 2.5 cm high. The platform faced 42×25 cm grid of parallel 0.1 cm-caliber stainless steel bars spaced 1 cm apart. In training session, the animals received 0.5 mA, scramble foot shock for 2 sec immediately upon stepping down. The interval of gerbils stepping down and placing all four paws on the grid was defined as the latency time. The latency over 180 sec was counted as 180 sec.
5. Radial 8-arm maze task
The spatial memory was determined using a radial 8-arm maze apparatus. The radial-arm maze apparatus consisted of a central octagonal plate (30 cm in diameter) and radiating eight arms (50 cm in length and 10 cm in width). The apparatus was placed 1 m above the floor. A small receptacle filled with water (3 cm in diameter and 1 cm in depth) was located at the end of the arms.
The rats were trained three times before the spatial memory test.
The rats deprived of water for 24 hours were allowed to explore for water and to drink water for 5 min. Test was conducted on the one day before sacrificed. The time spent for the seeking of water at the end of the arms was counted. The test was terminated when a rat found water in all eight arms or over 8 min elapsed. Re-entering to the previously visited arm was counted as error. In addition, the number of correct choice before the first error was counted.
6. Tissue preparation
The rats were sacrificed on the 57 day of the starting of experiment (One day after determining the radial 8-arm maze task). The animals were weighted and then overdosed with Zoletil 50
Ⓡ(10 mg/kg, i.p.; Vibac Laboratories, Carros, France). After a complete lack of response was observed, the rats were transcardially perfused with 50 mM phosphate-buffered saline (PBS) and fixed with a freshly prepared solution consisting of 4%
paraformaldehyde in 100 mM phosphate buffer (pH 7.4). The brains were dissected and pos-fixed in the same fixative overnight, and then the brains were transferred into a 30% sucrose solution for cryoprotection. Serial coronal sections of 40 μm thickness were made using a freezing microtome (Leica, Nussloch, Ger- many). Ten slice sections on average in the hippocampus were collected from each rat. The sections of 2.5 mm to 2.7 mm posterior from the bregma were used for immunohistochemistry.
7. TUNEL staining
To visualize DNA fragmentation, a marker of apoptosis, TUNEL staining was performed using an In Situ Cell Death Detection Kit
Ⓡ(Roche, Mannheim, Germany) according to the manufacturer’s protocol (Ko IG et al., 2009). The sections were post-fixed in ethanol-acetic acid (2 : 1) and rinsed. The sections were then incubated with proteinase K (100 μg/ml), rinsed, and incubated in 3% H
2O
2, permeabilized with 0.5% Triton X-100, rinsed again, and incubated in the TUNEL reaction mixture. The sections were rinsed and visualized using Converter-POD with 0.03% 3,3’-diaminobenzidine (DAB). Mayer’s hematoxylin (DAKO, Glostrup, Denmark) was used as a counter staining, and the sections were mounted onto gelatin-coated slides. Slides were air-dried overnight at room temperature, and coverslips were mounted using Permount
Ⓡ.
8. Caspase-3 immunohistochemistry
The sections were selected from each brain and incubated over-
night with mouse anti-caspase-3 antibody (1 : 500; Santa Cruz
Biotechnology, Santa Cruz, CA, USA), and then with biotinylated
mouse secondary antibody (1 : 200; Vector Laboratories, Burlin-
game, CA, USA) for another hour. The secondary antibody was
amplified with the Vector Elite ABC kit
Ⓡ(1 : 100; Vector Labo-
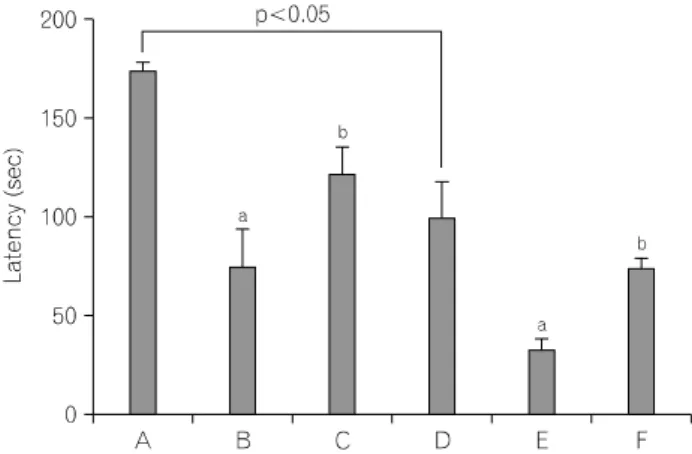
Fig. 1. Effect of treadmill exercise on latency in the step-down avoidance
task. (A) Young-aged control group, (B) young-aged and LPS injection group, (C) young-aged and LPS njection with treadmill exercise group, (D) old-aged control group, (E) old-aged and LPS injection group, (F) old-aged and LPS injection with treadmill exercise group. The results are presented as the mean±standard error of the mean (S.E.M.).
aRepresents p<0.05 compared to control group.
bRepresents p<0.05 compared to LPS-injection group.
ratories). Antibody-biotin-avidin-peroxidase complexes were visua- lized using 0.03% DAB, and the sections were mounted onto gelatin-coated slides. The slides were air-dried overnight at room temperature, and coverslips were mounted using Permount
Ⓡ.
9. Data analysis
The numbers of TUNEL-positive and caspase-3-positive cells in the hippocampal dentate gyrus were counted hemilaterally under a light microscope (Olympus, Tokyo, Japan), and they were expressed as the numbers of cells per square millimeter in the dentate gyrus. The area of the dentate gyrus was measured by Image-Pro
ⓇPlus image analysis system (Media Cyberbetics Inc., Silver Spring, MD, USA.).
Statistical analysis was performed using one-way ANOVA followed by Duncan's post-hoc test, and the results are expressed as the mean±standard error of the mean (S.E.M.). Significance was set as p<0.05.
RESULTS
1. Effect of treadmill exercise on the short-term memory in the step-down avoidance task The latencies of the step-down avoidance task are presented in
Fig. 1. The latency was 174.83±3.48 sec in the young-aged control group, 75.33±18.77 sec in the young-aged and LPS injection group, 122.66±13.21 sec in the young-aged and LPS injection with treadmill exercise group, 100.00±18.29 sec in the old-aged control group, 33.66±5.35 sec in the old-aged and LPS injection group, 74.83±4.55 sec in the old-aged and LPS injection with treadmill exercise group.
The present results showed that latency in the old-aged rats was significantly decreased compared to that in the young-aged rats (p<0.05). LPS treatment decreased latency both in the young-aged rats and in the old-aged rats (p<0.05), whereas, treadmill exercise increased latency both in the young-aged rats and in the old-aged rats (p<0.05).
2. Effect of treadmill exercise on the spatial memory in the radial 8-arm maze task The time of successful performance, the number of the correct, and the number of error choice in the radial 8-maze task are presented in Fig. 2. The time of the successful performances was 139.33±16.22 sec in the young-aged control group, 209.50±
21.96 sec in the young-aged and LPS injection group, 151.16±
11.01 sec in the young-aged and LPS injection with treadmill exercise group, 296.66±40.03 sec in the old-aged control group, 356.50±30.35 sec in the old-aged and LPS injection group, 274.83±21.56 sec in the old-aged and LPS injection with treadmill exercise group. The number of correct choice before the first error was 6.33±0.14 in the young-aged control group, 5.33±0.73 in the young-aged and LPS injection group, 6.16±0.32 in the young-aged and LPS injection with treadmill exercise group, 4.66±0.41 in the old-aged control group, 3.33±0.37 in the old-aged and LPS injection group, 4.50±0.57 in the old-aged and LPS injection with treadmill exercise group. The number of error made before eight successful performances was 3.88±0.36 in the young-aged control group, 9.16±0.58 in the young-aged and LPS injection group, 6.33±0.54 in the young-aged and LPS injection with treadmill exercise group, 13.50±0.66 in the old-aged control group, 16.66±0.73 in the old-aged and LPS injection group, 13.16±0.32 in the old-aged and LPS injection with treadmill exercise group.
The present results showed that the time for the successful
performances was longer, the number of the correct was lower,
and the number of error choice was higher in the old-aged rats than those in the young-aged rats (p<0.05). LPS treatment increased the time for the successful performances and the number of error choice both in the young-aged rats and in the old-aged rats (p<0.05), whereas, treadmill exercise decreased LPS-induced increase in the time for the successful performances and the number of error choice both in the young-aged rats and in the old-aged rats (p<0.05). LPS treatment decreased the number of correct choice in the old-aged rats, whereas, treadmill exercise increased the number of the correct in the LPS-treated old-aged rats.
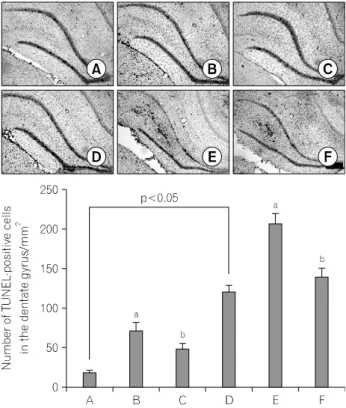
3. Effect of treadmill exercise on the number of TUNEL-positive cells in the hippocampal dentate gyrus
Photomicrographs of TUNEL-positive cells in the hippocampal dentate gyrus are presented in Fig. 3. The number of TUNEL-positive cells was 18.49±2.06/mm
2in the young-aged control group, 71.93±9.60/mm
2in the young-aged and LPS
injection group, 48.18±7.13/mm
2in the young-aged and LPS injection with treadmill exercise group, 121.78±8.15/mm
2in the old-aged control group, 207.78±13.08/mm
2in the old-aged and LPS injection group, 140.81±10.39/mm
2in the old-aged and LPS injection with treadmill exercise group.
The present results showed that the number of TUNEL-posi- tive cells in the old-aged rats was significantly enhanced compa- red to that in the young-aged rats (p<0.05). LPS treatment enhanced the number of TUNEL-positive cells both in the young-aged rats and in the old-aged rats (p<0.05), whereas, treadmill exercise suppressed LPS-induced increase in the number of TUNEL-positive cells both in the young-aged rats and in the old-aged rats (p<0.05).
4. Effect of treadmill exercise on the number of caspase-3-positvie cells in the hippocam- pal dentate gyrus
Photomicrographs of caspase-3-positive cells in the hippocampal
dentate gyrus are presented in Fig. 4. The number of
Fig. 2. Effect of treadmill exercise on performance time, correct number,and error score in the radial 8-arm maze task. (A) Young-aged control
group, (B) young-aged and LPS injection group, (C) young-aged and LPS
njection with treadmill exercise group, (D) old-aged control group, (E)
old-aged and LPS injection group, (F) old-aged and LPS injection with
treadmill exercise group. The results are presented as the mean±standard
error of the mean (S.E.M.).
aRepresents p<0.05 compared to control
group.
bRepresents p<0.05 compared to LPS-injection group.
Fig. 3. Effect of treadmill exercise on the DNA fragmentation in the
hippocampal dentate gyrus. Upper: Photomicrographs of terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL)-positive cells in the hippocampal dentate gyrus. (A) Young-aged control group, (B) young-aged and LPS injection group, (C) young-aged and LPS njection with treadmill exercise group, (D) old-aged control group, (E) old-aged and LPS injection group, (F) old-aged and LPS injection with treadmill exercise group. The scale bar represents 200 μm. Lower: Number of TUNEL-positive cells in each group. The results are presented as the mean±standard error of the mean (S.E.M.).
a
Represents p<0.05 compared to control group.
bRepresents p<0.05 compared to LPS-injection group.
Fig. 4. Effect of treadmill exercise on the caspase-3 expression in the
hippocampal dentate gyrus region. Upper: Photomicrographs of caspase-3-positive cells in the hippocampal dentate gyrus. (A) Young-aged control group, (B) young-aged and LPS injection group, (C) young-aged and LPS jection with treadmill exercise group, (D) old-aged control group, (E) old-aged and LPS injection group, (F) old-aged and LPS injection with treadmill exercise group. The scale bar represents 200 μm. Lower: Number of caspase-3-positive cells in each group. The results are presented as the mean±standard error of the mean (S.E.M.).
a