N-메철-4-페날퍼러디니움 및 6-허드록시도파민으로 유도된 도파민계 신경세포 사멸 기작의 규명
오 영 준*, 최 원 석
연세대학교이과대학생물학파
(Received December 18. 1996)
Characterization of Dopaminergic Neuronal Cell Death Induced by either N-Methyl-4-phenylpyridinium or
6-hydroxydopamine
Yong J. Oh* and WorrSeok Choi
Department of Biology, Yonsei University College of Science, Seoul 120-749
Abstract--Even though both N-methy 1-4-phenyl-l,2,5,6-tetxahydropyridine (MPTP) and 6-hy
droxydopamine have been widely used to establish the experimental model for dopaminergic neuronal cell death, mechanisms underlying this phenomenon have not been firmly explored. To investigate how these dopaminergic neurotoxins induce neuronal cell death, murine dopaminer^c neuronal cell line. MN9D cells were treated with various concentration of either 6-hydroxydopaniine or active form of MPTP. N-methyl-4-'phenylpyridinium (MPP^). Treatment of cells with 5-100 uM G-hydroxydopamine resulted in apoptotic cell death whereas cell death induced by 5-50uM MPP* was not demonstrated typical apoptotic characteristics such as cell shrinkage, apoptotic body and nuclear condensation. Cell death induced by 6-hydroxydopamine was partially blocked in the presence of antioxidants including soluble form of vitamin E or desferrioxamine suggesting that generation of oxidative stress may be as
sociated with 6-hydroxydopamine-induced cell death in MN9D cells. In contrast, MPFMnduced cell death was not blocked by treatment with any of antioxidants tested. As previously demonstrated that MPP^ caused metabolic alterations such as glucose metabolism, removal of glucose from the medium partially inhibited MPP^'-induced cell death suggesting excessive cycles of glycolysis may be associated with MPP'^-induced cell death. Taken together, these studies demonstrate that two types of do
paminergic neurotoxins recruit distinct neuronal cell death pathways.
중 추 신 경 계 퇴 행 성 질 환 의 대 표 적 인 에 인 파 킨 슨 증 후 군 은 nigrostriatal pathw ay에 존 재 하 는 도 파 민 계 신 경 세 포 가 사 멸 함 으 로 써 일 어 나 는 병 리 현 상 으 로 서 이 환 자 에 게 서 는 운 동 기 능 외 이 상 으 로 인 한 증 상 인
rhythmical tremor, rigidity, akinesia, bradykine- sia등 이 나 타 나 게 된 다. 이 에 대 한 연 구 는 단 기 적 인 처 료 를 목 적 으 로 L-dopa등 을 투 여 하 여 고 갈 된 도 파 민 을 대 체 하 는 등 의 약 물 학 적 인 방 범 으 로 부 터. 태 아 중 뇌 세 포 외 이 식 등 에 이 르 기 까 지 다 각 적 인 방 법 이 실 험 적 으
"본 논문에 관한 문의는 이 저자에게로 ( 전화) 02-361-2662 (팩스) 02-312-5657
로 사 용 되 여 왔 다. 또 한. 도 파 민 합 성 외 rate-limiting enzyme인 tyrosine hydroxylase나 신 경 세 포 의 생 존 과 관 련 된 것 으 로 알 려 진 여 러 중 류 외 neurotrophic factor등 의 유 전 자 를 세 포 내 로 주 입 시 킨 후, 실 험 적 으 로 유 도 된 동 물 모 델 외 뇌 로 투 입 하 여 이 둘 유 전 자 들 의 기 능 을 분 석 하 는 방 법 등 도 시 용 되 고 있 다. 이 와 같 은 연 구 가 가 능 하 였 던 것 온 바 로 파 킨 슨 증 후 군 외 실 험 동 물 모 델 을 제 조 할 수 있 는 도 파 민 계 룩 이 성 신 경 득 소 인 6- hydroxydopamine 이나 N-m ethyl-4-phenyl-l .2.5, 6-tetra-hydropyridine (M PTP) 이 존 재 하 였 기 때 문 이 다 (Heikkila an Cohen, 1971; Gerlach et al., 1991). 이 돌 은 dopamine transporter들 통 하 여 도 파
86
민 신 경 세 포 내 로 들 어 가 세 포 내 의 발 생 기 산 소 를 증 가 시 키거나 마 이 토 콘 드 리아 의 electron transport chain의 기 능 을 각 각 억 제 시 킴 으 로 서 이 들 신 경 세 포 의 사 멸 을 초 래 하 는 것 으 로 보 고 는 되 고 있 으 나 (Nicklas
et al.. 1985)
아 직 도 정 확 한 작 용 기 전 이 나 이 들 약 물 의 처 리 로 유 도 된 신 경 세 포 사 멸 외 형 태 가 확 립 되 있 지 못 하 다.위 에 서 언 급 된 바 와 같 은 연 구 외 주 방 향 은 파 킨 슨 증 후 군 으 로 인 한 증 상 자 체 룰 일 시 적 으 로 개 선 해 보 는 데 초 점 을 두 어 진 행 되 어 왔 다. 이 와 는 달 리 파 킨 슨 증 후 군 의 직 접 적 인 원 인 인 도 파 민 계 신 경 세 포 외 사 멸 과 정 과 사 멸 의 형 태 및 기 작 자 체 를 밝 히 는 등 과 같 은 연 구 의 펄 요 성 은 최 근 에 세 포 사 멸 의 한 형 태 인 apoptosis에 대 한 연 구 가 활 발 해 지 면 서 더 욱 부 각 되 기 시작 하 였 다
(Thompson, 1995). Apoptosis는 necrosis와 대 별 되 어 발 생 과 정 중 에 광 범 위 하 게 나 타 나 는 세 포 사 멸 의 한 형 태 로 서 대 개 의 경 우 에 세 포 크 기 의 측 소 (shrin- kage of cell size), 세 포 막 의 돌 출 (membrane bleb- bing), 및 세 포 막 으 로 쌓 여 져 서 떨 어 져 나 온 apop- totic body의 형 성 과 같 은 세 포 막 의 변 화 가 수 반 되 는 특 징 을 가 지 고 있 다. 또 한, chromatin condensation
이 나 DN A fragmentation등 파 같 은 핵 내 의 번 화 도 동 반 된 다 (Wyllie, 1993).
Apoptosis는 발 생 과 정 에 서 뿐 만 아 니 라 외 부 의 자 극 에 의 한 여 러 형 태 외 병 리 적 인 세 포 의 사 멸 에 도 관 련 되 어 있 옴 이 밝 혀 지 고 있 다 (Reed. 1994). 그 대 표 적 인 예 로 는 gamma- 또 는 UV-radiation, heat shock, lipid peroxidation-inducing reagents, tumor necrosis factor, virus infection 및 chemoth- erapeutic drug등 의 처 리 로 야 기 된 세 포 외 사 멸 현 상 을 들 수 가 있 다. 실 제 로 많 은 연 구 가 hemopoietic cells이 나 암 세 포 들 을 이 용 하 여 apoptosis의 발 생 여 부 가 암 이 나 자 가 면 역 질 환 등 의 발 현 과 밀 접 한 관 련 이 있 다 는 것 을 밝 혀 왔 으 며 , 이 와 같 은 연 구 의 방 향 은 병 리 적 상 황 의 치 료 틀 위한 연 구 의 일 환 으 로 현 재 까 지 진 행 되 여 지 고 있 다 (Thompson, 1995). 이 와 는 달 리 특 정 신 경 세 포 의 사 멸 로 발 생 하 는 중 추 신 경 계 퇴 행 성 질 환 과 같 은 병 리 상 황 에 는 어 떠 한 형 태 의 세 포 사 멸 현 상
(apoptosis 흑 은 necrosis) 이 발 생 하 는 지 와 이 에 관 련 된 기 작 등 에 대 한 연 구 가 아 주 미 약 한 상 태 이 며, 이 와 같 은 질 환 이 사 회 적, 경 제 적 으 로 미 처 는 영 향 의 심 각 성 에 비 추 어 연 구 의 필 요 성 이 그 어 느 때 보 다 도 대 두 되 고
있 는 실 정 이 다 (Thompson, 1995). 따 라 서, 본 연 구 는 이 와 같 은 목 표 를 추 구 하 기 위 하 여 최 근 에 제 조 되 고 성 격 이 규 명 된 도 파 민 계 신 경 세 포 주 인 M N9D라 는 세 포 를 이 용 하 여 (Choi et al., 1991; T ang et al., 1994) 실 험 적 으 로 도 파 민 계 특 이 성 신 경 득 소 를 처 리 하 여 세 포 사 멸 을 유 도 한 후, 세 포 사 멸 의 과 정, 형 태 및 기 작 을 밝 히 고 저 하 였 다. 본 연 구 의 결 과 에 의 하 면 i) 6~hydroxy- dopamine의 처 리 로 유 도 된 세 포 사 멸 은 apoptosis특 이 성 변 화 가 동 반 됨 에 비 하 여 M P T P외 활 성형 인 N - methyl-4-phenylpyridinium (M P P"*")의 처 리 로 유 도 된 세 포 사 멸 은 이 와 같 은 현 상 이 나 타 나 지 않 았 으 며; ii) 6-hydroxydopamine을 처 리 했 을 경 우 에 는 발 생 기 산 소 에 외 한 apoptosis가 초 래 되 였 으 며, antioxi- d ants외 처 리 로 사 멸 이 둔 화 되 는 것 이 관 찰 되 였 으 나 반 면 에 M PP^ 를 처 리 했 을 경 우 에 는 발 생 기 산 소 에 외 한 직 접 적 인 영 향 없 이 glucose up take외 증 가 에 따 른 과 도 한 glycolysis에 의 해 세 포 사 멸 이 발 생 함 을 밝 혔 다.
따 라 서 , 두 도 파 민 계 특 이 성 신 경 득 소 들 이 다 른 작 용 기 전 에 의 해 각 기 특 정 적 인 형 태 의 세 포 사 멸 을 유 발 시 키 는 것 으 로 나 타 나, 향 후 이 와 같 은 득 소 를 이 용 한 동 물 실 험 모 델 의 결 과 해 석 에 새 로 운 시 각 을 제 공 할 것 으 로 사 료 된 다.
실험방법
Immunocytochemistry - M N9D cell을 round glass cover slip에 서 배 양 시킨 후, cover slip을 P B S
로 3번 세 척 한 다 옴. 4% paraform aldehyde의 존 재 하 에 서 5분 간 고 정 을 하 였 다. P B S로 다 시 3번 세 척 한 후,
\%
bovine serum album in과 0.1% Triton X-100를 포 함 하 는 P B S로 primary antibody ( 1 :50 mouse monoclonal anti-tyrosine h yd ro xylase: B oehringer M annheim) 를 희 석 하 여 상 온 에 서 1 시 간 동 안 반 응 을 시 켰 다. 그 후 P B S로 3차 례 세 척 을 한 후
FITC-conjugated goat anti-m ouse antibody ( 1:
100; Cappell) 존 재 하 에 상 온 에 서 40 분 간 배 양 하 고, P B S로 다 시 세 척 을 하 였 다. 형 광 현 미 경 으 로 관 찰 하 기 위 하 여 1:1의 glycerol :P B S 용 액 으 로 cover slip을
mounting 하 였 으 며, 결 과 는 K odak Ektachrom e 400을 사 용 하 여 촬 영 하 였 다.
Reverse transcription-polymerase chain reaction (RT-PCR) - MN9D cell을 24-well p late에 서 배 양 하
여 이 로 부 터 total RN A를 acid-phenol을 사 용 하 는
Chom czynski와 Sacchi (1987) 의 방 법 을 이 용 하 여 추 출 하 였 다. R T-PCR은 Gandelm an동 이 기 술 한 방 법 (1991) 을 약 간 변 형 하 여 실 시 하 였 다. 이 때 tyro- sine hydroxylase 유 전 자 의 primer로 사 용 된 oli- gonucleotide의 염 기 배 열 은 reverse primer외 경 우
5느 A G G CA TG G C G G A TA T A CT-3^ 이 며 forward prim er의 경 우 에 는 5 -A T A T G T C A C G C T G A A - GGG-3 서 며 이 둘 은 각 각 Exon 7과 8에 위 처 한 염 기 배열 로 부 터 선 정 하 였 다. Reverse transcription 반 웅 을 위 하 여 0.5 ug total RN A가 사 용 되 였 고 10 U의
A M V reverse transcriptase (Promega) 를 가 해 주 어
4Z'C에 서 배 양 함 으 로 서 cDNA틀 제 조 하 였 다. 이 와 같 이 생 성 된 cDNA틀 사 용 하 여 tyrosine hydroxylase m R N A외 존 재 여 부 를 polymerase chain reaction으 로 측 정 하 였 다. 이 때 사 용 된 denaturation, an
nealing 및 extension tem perature■는 각 각 94°C, 5(TC. 72°C이 며 총 25 cycle을 실 시 하 였 다.
Cell culture - MN9D cell은 P-100 dish (Coming)
에 서 대 량 배 앙 을 하 였 으 며 본 실 험 을 위 하 여 서 는
0.25% trypsin/0.05% DNase로 처 리 를 하 여 single cell suspension을 만 든 후, trypan blue exclusion 방 범 을 사 용 하 여 hematocytometer로 세 포 수 를 세 였 다.
이 로 부 터 4xl(y* MN9D cell을 0.1 mg/m/외 poly-D- lysine (Sigma) 으 로 도 포 된 24-well plate (Coming)
에 plating을 한 다 옴. 3일 간 10% CO2와 습 기 가 일 정 하 게 유 지 되 는 37T:세 포 배 양 기 에 서 계 대 배 양 을 하 였 다. 사 용 된 medium 외 조 성 은 high-glucose가 보 급 된
DMEM (Gibco) 과 10% heat-inactivated fetal bo
vine serum (F B S; Gibco) 을 포 함 하 였 다. 본 실 험 직 전 에 DMEM으 로 3번 세 척 한 다 옴, 5 -10 0 uM 6-hy- droxydopamine 이 나 M P P ^ 단 득, 또 는 an- tioxidant돌 을 figure에 명 기 된 바 와 같 이 병 용 처 러 를 위 하 여 N2 serum-free medium (Bottenstein and Sato, 1979) 으 로 교 체 한 후 일 간 계 속 배 양 하 였 다.
이 배 앙 기 간 이 끝 나 면 아 래 항 에 명 기 된 방 범 으 로 실 험 결 과 를 동 정 하 였 다.
MTT reduction assay - 세 포 외 생 존 율 을 측 정 하 기 위 하 여 Fig. 2에 명 기 된 농 도 의 약 물 을 단 득 또 는 병 행 투 여 한 후 일 정 시 간 후 에 Hansen등 이 기 슬 한 방 법 을
(1989) 이 용 하 여 3-[4,5-dimethylthiazol-2-yl]-2.5- diphenyltetrazolium bromide (M T T: Sigma) as-
say를 행 하 였 다. 간 단 허 기 술 하 면 상 기 에 기 술 된 바 와 같 이 도 파 민 계 신 경 득 소 로 단 득. 또 는 병 용 처 러 된 세 포 외 배 양 액 에 최 종 농 도 가 1 mg/m/ 가 되게 M TT를 가 한 다 옴 2시 간 가 량 더 배 양 을 시 켜 생 존 세 포 돌 외 dehy- drogenase activity에 의 해 M TT가 환 원 되 도 록 했 다.
그 후 20% sodium dodecylsulfate가 포 함 된 50%
dimethylformamide수 용 액 을 가 하 여 세 포 내 에 형 성 된 formazan grain을 용 즐 해 낸 다 옴 에 ELISA plate reader를 사 용 하 여 540 nm상 에 서 optical den- sity를 측 정 하 였 다. 각 실 험 군 으 로 부 터 얻 어 진 실 험 결 과 는 약 물 이 처 러 되 지 않 은 대 조 군 (100% ) 에 대 한 상 대 값 으 로 표 현 하 였 다.
TUNEL staining- 이 방 법 외 원 리 는 D N A가
apoptosis시 잘 려 지 게 되 면 3' end에 -0 H기 가 생 기 게 되 며 이 곳 에 terminal transferase를 이 용 하 여 면 역 반 응 검 출 에 용 이 한 nucleotide틀 붙 혀 주 는 데 있 다.
간 략 히 기 술 을 하 면 약 물 로 처 리 된 세 포 를 phos- phate-buffered saline (PB S) 로 3차 례 세 척 해 준 뒤
4% paraformaldehyde로 상 온 에 서 5분 간 고 정 을 시 킨 후 P B S로 다 시 3 차 례 세 척 을 하 였 다. Proteinase K (20 g/m/ ; Boehringer Mannheim) 존 재 하 에 상 온 에 서 5분 간 배 양 하 여 DN A에 붙 어 있 는 단 백 질 을 제 거 시 킨 다 옴 P B S로 3차 례 에 걸 쳐 세 척 하 였 다. 이 후
Terminal transferase 배 양 액 존 재 하 에 37°C에 서 1시 간 동 안 반 응 을 시 켰 다. 이 때 사 용 한 400 U/ 배 양 액 내 외 조 성 은 2 .41 1/ biotinylated dUTP (Boehringer M a n n h e im ), 4.8 u l te rm in a l tra n s fe ra s e
(Boehringer Mannheim) 을 포 함 하 였 다. 반 응 후 에
300 mM sodium chloride/30 mM sodium citrate존 재 하 에 상 온 에 서 15 분 간 배 양 하 여 te rm in a l transferase외 반 응 을 증 료 시 킨 다 옴 alkaline phos
phatase-linked avidin을 가 한 후 상 온 에 서 30분 간 배 양 시 켰 다. Substrate를 가 하 여 반 응 의 결 과 를 색 깔 로 발 현 시킨 것 을 light microscope로 관 찰 한 후 K o- dak Ektachrome 160T로 촬 영 을 하 였 다.
실험결과 및고찰
Characterization of MN9D cells - 본 설 험 에 사 용 된
MN9D cell은 embryonic day 14의 mouse mesen
cephalic neuron과 N18TG neuroglioma의 융 합 반 응 을 통 해 확 립 된 cell line으 로 서 도 파 민 계 신 경 세 포 특 유
J. Pharm. Soc. Korea
의 여 러 가 지 성 질 을 가 지 고 있 는 것 으 로 밝 혀 져 있 다
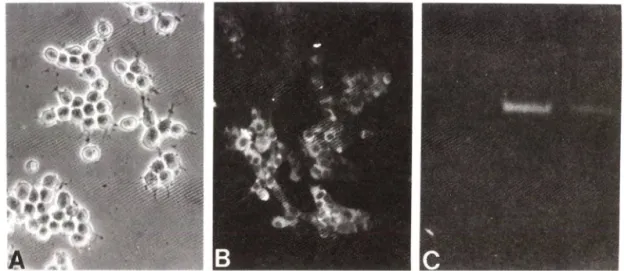
(Choi et a l, 1991: Tang et a l, 1994). Fig. lA에 서 보 듯 이 정 상 적 인 상 태 외 MN9D cell의 세 포 모 양 은 동 그 라 며 . neurite이 존 재 하 되 아 주 짧 은 것 이 특 정 이 다. Im- munocytochemistry 와 reverse transcription-po - lymerase chain reaction의 결 과 에 서 보 듯 이 MN9D cell내 에 는 도 파 던 합 성 의 rate-limiting enzyme인
tyrosine hydroxylase외 mRNA와 단 백 질 이 모 두 존 재 하 고 있 옴 을 밝 혔 다(Fig. IB, C). 이 외 에 도 도 파 민 계 신 경 세 포 가 갖 는 다 른 특 성 중 의 하 나 인 도 파 민 의 재 흡 수
(dopamine uptake) 가 MN9D cell에 서 도 이 루 어 짐 으 로 서 SV40 large T antigen등 으 로 변 형 되 여 생 성 된
cell line 과 는 달 리 cellular phenotype을 유 지 하 는 장 점 을 가 짐 과 동 시 에. 생 체 내 에 존 재 하 는 도 파 민 계 신 경 세 포 로 서 외 거 외 모 든 기 능 을 유 지 함 으 로 서 정량 적 인 설 험 을 목 적 으 로 하 는 연 구 에 사 용 될 수 있 는 좋 은
model system임 이 확 인 되 였 다.
Cell death induced by dopaminergic neurotoxins - 우 선 적 으 로 도 파 민 계 특 이 성 득 소 로 널 러 사 용 되 어 온 6-hydroxy dopamine 과 M PP 객 작 용 력 을 검 증 하 기 위 하 여 각 각 을 5 M 0 0 u M로 2 4 -4 8 시 간 처 리 한 후. MTT assay를 통 하 여 세 포 생 존 율 을 측 정 하 였 다. Fig. 2에 서 보 듯 이 MN9D cell은 가 해 준 득 소 외 농 도
에 비 례 하 여 사 멸 함 을 관 찰 하 였 으 며. 이 농 도 외 범 위 는
primary dopaminergic neuronal culture® 이 용 한 여 러 실 험 에 서 사 되 여 온 점 으 로 미 루 어 보 아 두 신 경 득 소 에 MN9D cell도 endogeneous counterpart와
B)
6>hydroxydopamme (나M) MPP* (^M)
Fig. 2 — Kinetics of neurotoxinnnduced cell death. MN9D cells were plated at 3x10
*흐cells/24-well plates and maintained in DMEM supplemented with 10% fetal bovine serum for 3 day. Ceils were switched to serum-free N
2medium containing in dicated concentration of (A) 6~hydroxydopamine for 24 horn's or (B) MPF" for 48 hours. Viability of cells was measured by MTT reduction assay.
Results represent the mean of three in dependent experiments in triplicate±SEM.
Value from each treatment were expressed as a percent control over the non-treated matching control(100% survival).
Fig. 1 — Characterization of MN9D dopaminergic neuronal cell line. Cells were plated onto 25 ug/m/ poly-D-Iysine coat
ed 24-well plate(Coming). Immunocytochemical localization of tyrosine hydroxylase was carried out as des
cribed in Materials and Methods. Reverse transcription-polymerase chain reaction (RT-PCR) was performed as described in Materials and Methods using total RNA as a template. (A) Phase-contrast micrograph of MN9D ceils and (B) immunofluorescent localization of tyrosine hydroxylase. (C) RT-PCR agarose gel stained with ethidium bromide (Lane 1) negative control, without template
:(Lane 2) with 50 ng total RNA from M N 9D cells
;(Lane 3) positive control, with plasmid containing tyrosine hydroxylase cDNA.
라
아
비 숫 한 sensitivity가 있 음 을 보 여 주 었 다. 다 옴 으 로 는 이 와 같 이 유 도 된 세 포 사 멸 의 형 태 를 비 교 및 분 석 하 기 위 하 여 각 각 의 신 경 득 소 로 처 리 된 세 포 를 상 기 와 같 이 배 양 한 후 에 phase-contrast microscope상 에 서 사 멸 외 과 정 을 관 찰 하 였 다. 즉. 6-hydroxydopamine으 로 처러 한 경 우 에 는 apoptosis시 에 나 타 나 는 전 형적 인 변 화 인 nuclear condensation, membrane shrink
age 및 apoptotic body가 생 성 되 였 다 (data not shown). 이 와 는 대 조 적 으 로 M PP우로 처 리 로 유 도 된 세 포 사 멸 외 경 우 에 는 nuclear fragm entation이 나
nuclear condensation등 의 변 화 가 아 주 미세 하 였 으 며
membrane shrinkage나 apoptotic body외 형성등 과 같 은 cell membrane상 외 번 화 또 한 아 주 경 미 하 였 다
(data not shown). 이 와 같 은 결 과 를 바 탕 으 로 두 신 경 득 소 의 처 러 에 의 한 세 포 사 멸 이 apoptosis 인 지 를 더 윽 명확 히 밝 히기 위 하 여 chromatin condensation과
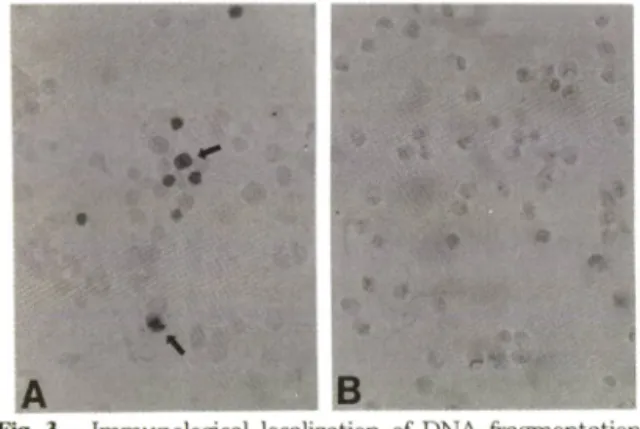
fragmentation 등 과 같 은 핵 내 외 변 화 를 즉 정 하 는 데 사 용 되 여 왔 던 TU N EL staining을 실 시 하 여 두 신 경 득 소 로 유 도 된 핵 내 의 번 화 를 구 체 적 으 로 검 증 하 였 다. Fig. 3A에 서 보 듯 이 6-hydroxydopamine으 로 유 도 된
MN9D cell외 사 멸 과 정 중 에 는 위 에 서 언 급 한 핵 내 의 번 화 가 수 반 되 는 신 경 세 포 외 핵 들 이 특 징적 으 로 염색되 는 것 을 관 찰 하 였 다. 반 면 에 M PP" 로 처 리 한 경 우 에 는 상 기 와 같 은 핵 내 외 번 화 가 없 움 을 확 인 하 였 다
(Fig. 3B). 그 러 나 이 경 우 에 도 apoptosis와 대 별 되 는
Fig. 3 — Immunological localization of DNA fragmentation in MN9D cells after neurotoxin treatment. Cells were plated, maintained and treated with (A) 100 uM 6-hydroxydopamine for 18 hours or (B) 50 uM MPP"^ for 36-40 hours. Cells were then pro
cessed for localization of fragmented DNA within the nuclei as described in Materials and Methods. Arrows indicated the positive nuclei in
dicative of typical apoptotic cell death.
형 태 외 세 포 사 멸 로 알 려 진 necrosis외 전 형 적 인 특 성 중 의 하 나 인 cell swelling이 나 cell lysis의 현 상 이 전 혀 관 찰 되 지 않 았 다. 따 라 서. M PP" 로 유 도 된 세 포 의 사 멸 은 D N A의 fragmentation을 아 주 제 한 적 으 로 동 반 하 는 특 이 한 형 태 외 apoptosis로 추 측 이 된 다. 이 를 간 접 적 으 로 뒷 받 침 하 는 증 거 로 서 M P P+ 에 의 해 유 도 된 세 포 사 멸 이 단 백 질 합 성 억 제 제 인 cycloheximide의 병 용 처 리 로 완 전 히 억 제 됨 을 밝 힌 점 을 들 수 가 있 다
(not shown). 이 는 유 전 적 으 로 이 미 정 해 진 세 포 사 멸 이 라 하 여 programmed cell death라 고 청 하 는 과 정 중 에 발 현 되 는 세 포 사 멸 유 전 자 가 거 의 모 든 경 우 에
apoptosis를 유 발 시 키 며. 따 라 서 세 포 사 멸 유 전 자 의 발 현 을 억 제 시 켜 주 는 macromolecule 합 성 억 제 제 인
cycloheximide나 actinomycin 등 을 처 러 해 주 면
apoptosis가 방 지 됨 이 밝 혀 져 왔 다(Martin et al..
1988; Oppenheim et al., 1990). 이 에 반 하 여
necrosis를 동 반 하 는 경 우 에 는 이 와 같 은 처 리 로 세 포 사 멸 이 전 혀 방 지 되 지 못 한 다 는 점 을 미 루 어 보 아. M PP'"에 외한 세 포 사 멸 은 apoptosis의 톡 이한 sub- type으 로 규 정 하 는 것 이 가 능 하 다 고 본 다. 이 와 같 은 현 상 은 도 파 민 계 특 이 성 신 경 득 소 에 서 뿐 만 아 니 라 일 반 적 으 로 널 러 사 용 되 는 prooxidants (hydrogen peroxide 또 는 menadione) 나 mitochondrial elec
tron transfer chain complex inhibitor인 ro- tenone이 나 3-nitropropionic acid틀 MN9D cell에 처 리 하 였 을 때 에 도 동 일 하 게 발 생 한 다(Oh et al., 1995). 즉, MPP"\ rotenone. 및 3-nitropropionic acid등 과 같 은 mitochondrial toxin의 처 리 로 m i- tochondria의 기 능 장 애 를 초 래 하 여 유 도 된 신 경 세 포 의 사 멸 은 기 존 의 apoptosis와 다 른 기 전 을 통 해 특 이 한 형 태 외 apoptosis를 유 발 하 는 것 으 로 사 료 된 다
(Oh et al., 1995).
Role for antioxidants in neurotoxin-induced cell death - 기 존 의 연 구 에 외 하 면 발 생 기 산 소 의 생 성 과 이 에 의 한 작 용 이 apoptosis와 관 련 된 것 으 로 밝 혀 져 있 다(Heikkila and Cohen, 1971: Hockenbery et
al., 1993
; Kane et al., 1993). 따 라 서. 본 실 험 에 서 는MN9D cell 의 경 우 에 6-hydroxydopamine 이 나
MPP + 에 외 한 세 포 사 멸 과 정 이 발 생 기 산 소 외 생 성 에 의 한 것 인 지 를 확 인 하 기 위 하 여 각 각 의 neurotoxin을
50~100uM의 농 도 로 처 리 함 과 동 시 에 antioxidant
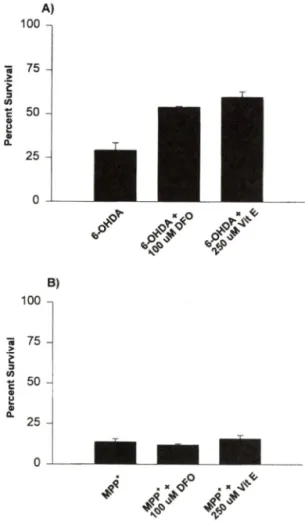
를 가 하 여 주 었 다. Fig. 4A에 서 보 듯 이 6-hy-
Pharm. Soc. Korea
droxydopamine에 의 한 세 포 사 멸 이 antioxidant로 널 리 사 용 되 는 desferrioxamine (100-^200 u M:
Kane et al.. 1993), soluble form인 Vitam ine E (250uM) 를 처 러 하 였 을 때 상 당 허 방 지 됨 을 밝 혔 다
(30%). 반 면 에 M P P ■뇌] 외 한 세 포 사 멸 은 이 둘 an tioxidants 나 Vitam in E등 을 병 행 투 여 하 여 도 전 혀 번 화 가 없 옴 을 관 찰 하 였 다(Fig. 4B). Oxygen species-
Fig. 4 — Effect of antioxidants in neurotoxin-induced cell death. Cells prepared as described above were ccrtreated with or without the indicated concentration of each antioxidant. At the end of treatm ent w ith (A) 100 uM 6-hy
droxydopamine plus 100 uM desferioxamine (DFO) or 250 uM soluble form of Vitamine E (Vit E) for 24 hours or (B) 50 uM MPP^ plus each antioxidant for 48 hours, cell viability was measured using MTT reduction assay.
Results were expressed as a percent survival over the non-treated control (100% survival).
Each bar represents the mean of three in
dependent experiments in triplicate+SEM.
sensitive dye인 2 .7 -dichlorofluorescin diacetate
을 이 용 하 여 생 성 된 발 생 기 산 소 의 양 을 측 정 한 예 비 실 험 의 결 과 에 의 하 면 MN9D cell에 M P F^를 처 리 한 경 우 에 발 생 기 산 소 가 생 성 은 되 는 것 으 로 나 타 났 으 며 상 기 의 antioxidant룰 처 리 하 였 을 때 에 발 생 기 산 소 외 생 성 이 억 제 는 되 나 신 경 세 포 의 사 멸 을 전 혀 방 지 하 질 못 하 였 다(not show n). 따 라 서 , 위 의 결 과 틀 증 합 해 보 면, M P P" 외 처 리 에 의 해 생 성 된 발 생 기 산 소 는 세 포 사 멸 의 직 접 적 인 원 인 온 아 니 며, 아 마 도 세 포 사 멸 과 정 중 에 이 차 적(secondary effect) 으 로 생 성 된 것 임 을 알 수 가 었 었 다(Gerlach et al.. 1991).
Role for the level of glucose in MPP'*-mediated cell death - 기 존 외 연 구 결 과 에 의 하 면 M P P누 m i
tochondrial electron transfer chain complex I외 기 능 을 방 지 함 으 로 서 이 에 따 른 에 너 지 대 사 의 기 능 이 상 을 초 래 하 여 도 파 민 계 신 경 세 포 의 사 멸 을 촉 진 시 키 는 것 으 로 밝 혀 져 었 다 (Nicklas et al.. 1985). 이 과 정 중 에 펄 연 적 으 로 예 측 되 는 세 포 내 의 번 화 로 는 m i
tochondria 에 의 한 정 상 적 인 A T P의 생 성 이 억 제 됨 으 로 서 glycolysis가 증 가 하 게 되 고, 그 결 과 로 서 세 포 내 에 lactic a d d와 proton이 측 적 이 되 여 세 포 가 사 멸 하 는 것 으 로 밝 혀 져 있 다. 따 라 서, glycolysis에 의 한 과 도 한 glucose의 u p tak e이 M P P"*"에 의한 MN9D cell
의 사 멸 이 직 접 적 으 로 관 련 이 었 는 지 를 확 인 하 기 위 하 여 100 uM M PP* 존 재 하 에 culture m edium중 의
glucose를 지 속 적 으 로 즐 여 주 면 서 세 포 사 멸 의 정 도 를 측 정 하 였 다. 이 실 험 의 결 과 에 의 하 면 Fig. 5에 서 보 듯 이 biphasic response현 상 이 나 타 났 다. 즉, 정 상 g lu cose level의 60% 까 지 즐 여 즐 경 우 에 dose-de
pendent 하 게 세 포 사 멸 이 방 지 됨 을 밝 힘 으 로 서 MPP^
의 처 리 에 의 한 신 경 세 포 외 사 멸 현 상 은 파 다 하 게 세 포 내 로 유 입된 glucose의 대 사 산 물 인 lactic acid나 pro
ton 의 측 적 으 로 인 한 세 포 내 의 pH외 산 성 화 가 주 원 인 일 것 으 로 주 즉 이 되 어 진 다(Tipton and Singer, 1993). Nigericin과 같 이 세 포 내 외 의 pH틀 산 성 으 로 유 도 시 켜 는 약 물 을 처 리 하 여 발 생 된 세 포 사 멸 이
M P P^ 로 유 도 된 세 포 사 멸 현 상 과 상 당 히 유 사 하 다 는 예 비 실 험 의 결 과 도 이 률 간 접 적 으 로 증 명 해 주 고 있 다
(not show n). 그 러 나 과 도 한 양 의 glucose가 배 지 로 부 터 제 거 되 는 경 우 (>60% ) 에 는 그 자 체 가 세 포 의 사 멸 을 촉 진 하 기 때 문 에 다 소 세 포 사 멸 방 지 능 력 이 떨 어 지 는 것 으 로 추 측 이 되 어 진 다 (Reed, 1994).
B
1
I - ---T---r 05050Q5050^52075211 I 해 AlAJnM ;uaaiedIBAIAJnw JUOOJad
0
Fig. 5
0 25 50 75 100
Glucose Amount (%)
Effect of the level of glucose in MPPMnduced cell death. Cells were treated with 50 uM MPF*"
for 48 hours in N
2medium containing a vari
ous level of glucose as indicated. Normally, N
2medium contains 4.5 g// of glucose (100%).
Data represent the mean of three independent experiments in triplicate+SEM .
이 상 외 결 과 에 의 하 면 파 킨 슨 증 후 군 외 연 구 모 델 을 설 정 하 기 위 하 여 널 리 사 용 되 여 왔 던
6-hydroxy- d o p a m in e
이 나M P P "
는 도 파 민 계 신 경 세 포 의 사 멸 을 다 른 기전 을 통 해 유 도 함 을 알 수 가 있 으 며 , 따 라 서 각 각 의 도 파 민 계 의 신 경 득 소 로 부 터 얻 어 진 결 과 외 해 석 에 유 의 할 펄 요 성 을 부 각 시 켰 다. 또 한.M N 9 D
와 같 은d o pam inergic n eu ron al cell lin e
을 이 용 할 경 우 에 본 실 험 과 같 온 정 량 적 인 해 석 이 필 요 한 다 각 적 인 연 구 가 가 능 케 되 며, 더 욱 이 최 근 에 밝 혀 지 기 시 작 하 는Bcl- 2
등 과 같 은anti-apoptotic gene
둘 이 신 경 세 포 의 사 멸 과 정 중 에 나 타 내 는 작 용 력 을 검 중 하 기 위 한gene transfection
등 외 연 구 가 가 능 케 되 었 다 고 본 다(T hom pson, 1995).
감사의 말씀
본 연 구 는 교 육 부 학 술 연 구 조 성 과 제 ( 유 전 공 학 분 야)
연 구 비 와 학 술 진 흥 재 단 (자 유 공 모 과 제 ) 의 지 원 으 로 수 행 되 였 으 며 이 에 감 사 드 럽 니 다.
문 헌
1) Bottenstein. J. E. and Sato, G. H.
:Grow th of a rat neuroblastoma cell line in serum-free sup
plemented m edium . Proc. N atl Acad. Sci. USA.
76.
514 (1979).
2) Chom czynski. P. an d Sacchi, N . - Single-step method of R N A isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. A nal Biochem.
162.156 (1987).
3) Choi. H. K.. W on, L. A.. Kontur, P. J .. H a m mond. D. N.. Fox. A. P.. W ainer. B. H .. Hoff
m ann. P. C. and Heller, A.
:Im m ortalization of embryonic m esencephalic dopam inergic n e u rons by somatic cell fusion. Brain Res.
552.67 (1991).
4) G andelm an. K. Y., H arm on. S.. Todd. R. D..
and O'M alley. K. L. • Analysis of the structure and expression of the h u m a n dopamine D2A re
ceptor gene. ]■ Neurochem.
56,1024 (1991).
5) Gerlach. M .. Riederer, P., Przuntek, H. and Youdim, M . B. H. • M PTP mechanisms of neu
rotoxicity and their implications for Parkinson's disease. Eur. J. Pharmacol.
208,273 (1991).
6) Hansen. M. B.. Nielson. S. E. and Berg. K. - R eexam ination an d further development of a precise a n d rapid dye m ethod for m easuring cell growth/cell kill. /. Immunol Metk.
119,203
(1989).
7) Heikkila. R. and Cohen. G.
:Inhibition of biogen
ic amine uptake by hydrogen peroxide
:a mec
hanism for toxic effects of G-hydroxydopamine.
Science
172,1257 (1971).
8) Hockenbery. D. M .. Oltvai. Z. N.. Yin. X. M., M illim an, C. L. an d Korsmeyer. S. J .
:Bch2 functions in an antioxidant pathw ay to prevent apoptosis. Cell
75.241 (1993).
9) Kane. D. J.. Sarafian, T. A., Anton. R . H ahn, H., G ralla, E. B ., Valentine. S. J .. Ord, T. and Bredesen. D. E.
:Bcl-2 inhibition of neural death
:Decreased generation of reactive oxygen species.
Science 262, 1274 (1993).
10) M artin. D. P., Schmidt. R. E .. DiSefano, P. S..
Lowry, D. H .. Carter. J . G. and Johnson. Jr. E.
M . ■ Inhibitors of protein synthesis an d R N A synthesis prevent neuronal cell death caused by nerve growth factor deprivation. /. Cell Biol.
106.
829 (1988).
11) Nicklas. W . J., Vyas, I. and Heikkila, R. E.
:In
/.