kfas
11
0
0
전체 글
(2) 김정배 . 박정임 . 이원찬 . 이근섭. 66. km2으로 잘 발달해 있다(Yoon, 2006; Kim et al., 2010). 섬진 강 하구에는 한국 연안에 서식하는 해초류 9종 중에서 애기거 머리말과 거머리말의 2종이 자생하고 있다(Kim et al., 2009a; Kim, 2012). 우리나라에 출현하는 거머리말은 대부분 조하대 에서 출현하지만, 섬진강 하구에서 생육하고 있는 거머리말은 조간대 하부지역의 수로부분에 patchy 형태로 자생하고 있으 며, 얕은 수심의 조하대에는 넓은 초지를 형성하여 출현하였다 (Kim et al., 2010). 해초는 대부분 조하대에서 생육하고 재생 산이 이루어지지만, 일부 종들은 조간대에서 생육하기도 한다 (Leuschner and Rees, 1993; Koch, 2001; Lee et al., 2006). 항 상 물에 잠겨있는 조하대와는 대조적으로 조간대 하부 거머리 말은 매일 2회 물이 빠져나가는 썰물 시간 동안 공기에 노출되 어 있으며, 건조의 위험, 극한 온도와 자외선 광에 의한 손상 이 일어날 수 있다(Leuschner and Rees, 1993; Hemminga et al., 1994; Leuschner et al., 1998; Shafer et al., 2007). 독일의 Wadden Sea의 조간대에는 Z. noltii와 Z. marina가 혼합되어 생육하고 있고, 조하대에는 Z. marina가 생육하고 있지만, 저 조수 때에는 건조에 의하여 조간대에 서식하는 Z. noltii와 Z. marina의 탄소동화작용에 대한 제한과 생성된 탄소의 양이 감 소하였다고 하였다(Leuschner and Rees, 1993; Leuschner et al., 1998; Shafer et al., 2007). 본 연구는 섬진강 하구유역의 조 간대와 조하대에 생육하는 거머리말의 계절에 따른 탄소와 질 소 성분 변화를 비교하고자 하였다.. 재료 및 방법 거머리말 자생지의 환경특성 거머리말 자생지의 환경 특성은 2003년 5월부터 2004년 12 월까지 월별로 조사하였다. 조사 정점은 조간대 하부(St. 1)와 조하대 상부 수심 2 m가 되는지역(St. 2)을 선정하였고,두 정 점간의 직선 거리차이는 약 1 km이었다(Fig. 1). 수온은 각 조사장소에 수온계(StowAway TidbitⓇ temperature data logger, Onset computer, USA)를 설치하여 매 15분 간격 으로 측정하였으며, 측정된 수온은 일일 평균수온으로 나타 내었다. 조사기간 동안 수중광량을 조사하기 위해 거머리말 잎이 위치하는 수심에 조도계(HOBO-Light Intensity, Onset Computer, USA)를 설치하여 매 15분 간격으로 측정하였다. HOBO 조도계로 측정된 수치(lumens.ft-2)는 동 시간에 광 측 정기(LI-1400, LI-COR, Inc)로 측정된 수치와의 회귀분석을 통하여 유입광량(PFD, mol photons m-2d-1)으로 변환하여 월 평균으로 나타내었다. 두 조사 장소에 조사 기간 중 일정기 간 해초의 엽체가 위치하는 수심에 수심계(HOBO-Water level logger, Onset computer, USA)를 각 정점에 설치하여 매 10분 간격으로 압력을 측정하였으며, 이 값을 수심으로 변환 하였 다. HOBO 수심계로 측정된 수치는 국립해양조사원에서 1시 간 간격으로 측정하여 제공하는 조위 수치와의 회귀분석을 통. 34° 55'N. 34° 54'N. 127° 47'E. 127° 50'E. Fig. 1. Study sites in Seomjin Estuary on the south coast of the Korean peninsula from May 2003 to December 2004 (St 1 : Lower intertidal, St 2 : Upper subtidal)..
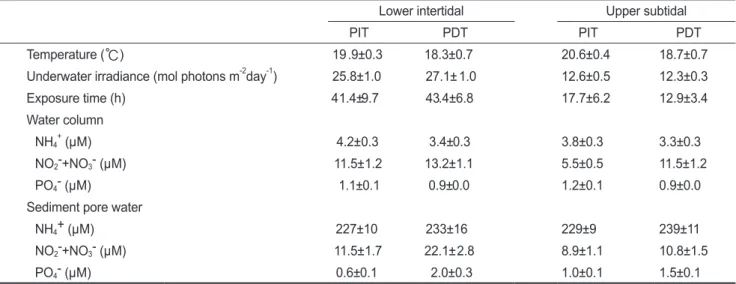
(3) 67. 조간대와 조하대 거머리말의 탄소 및 질소 성분 변화. 해 조사기간 동안 1시간 간격의 자료를 월간과 밤낮을 구분하 여 조간대 하부와 조하대 상부의 노출시간을 계산 하였다. 해 수와 퇴적물 공극수의 영양염류 농도를 측정하기 위하여 5-6개 씩 샘플을 채취하였고, 퇴적물은 지름 5 cm, 길이 13 cm의 주 상 시료 채취기를 이용하여 채취하였다. 퇴적물 공극수의 영양 염류 분석을 위한 시료는 퇴적물을 3,000 rpm에서 20분간 원 심 분리하여 추출한 다음 0.45 μm 막 여과지로 여과하여 분석 할 때까지 -20℃에서 냉동 보관하였다. 해수 및 퇴적물 공극수 의 암모늄염(NH4+-N), 아질산염(NO2--N)+질산염(NO3--N)과 인산염(PO4--P) 농도는 흡광광도법으로 흡광도를 측정하여 계 산하였다(MOMAF, 2002).. 었고, 시기는 수온이 해초의 생육을 조절하는 가장 중요한 요 인으로 작용하고 있으므로(Lee and Dunton, 1996; Lee et. al., 2005), 수온 상승기(3월에서 8월)와 수온 하강기(9월에서 이듬 해 2월)로 나누었다. 통계적으로 유의한 차이는 normality로 검정 한 후 two-way ANOVA로 homogeneity of variance와 유의성을 검정하였다 (P<0.05). 통계분석은 SPSS 10.1을 이용하였으며, 측정치는 평균(mean)과 표준오차(SE)로 나타내었다. 단위면적당 잎의 무게를 탄소와 질소 성분에 대하여 회귀분석을 실시하여 회귀 직선 식을 구하였다.. 결. 잎의 무게, 해초 조직 내의 탄소, 질소 성분 측정 조사 기간 동안 거머리말 잎의 단위 면적당 무게(mg dry wt cm-2)는 매월 성체 10개체 중 2-3번째 잎 중에서 잎 폭을 측정 하고, 10 cm길이로 절단하여 측정하였다. 거머리말 잎 및 지하 경 조직 내의 탄소 및 질소 농도분석을 위하여 매월 6-10개체의 성체 중에서 잎은 2-3번째 잎을 사용하였고, 지하경은 첫째 마 디에서 여섯째 마디까지의 지하경을 60℃ 에서 완전히 건조하 였다(Lee et al., 2003, 2004, 2005; Kim et al., 2009b). 건조된 시료를 분쇄한 후 2-3 mg을 분취하여 CHN 원소분석기(Flash EA1112, CE Instruments, UK)로 잎과 지하경 조직 내의 탄소 및 질소 성분을 측정 하였다.. 과. 거머리말 자생지의 환경 특성 거머리말 자생지의 환경 특성 인자인 수온, 광량, 노출시간, 해수와 퇴적물 공극수의 영양염류 농도에 대하여 시기별 평균 값을 Table 1에 나타내었다. 수온 범위는 조간대 하부 중 수온 상승기의 경우 19.9±0.3℃, 수온 하강기의 경우 18.3±0.7℃이었고, 조하대 상부 중 수온 상승기의 경우 20.6±0.4℃, 수온 하강기의 경우 18.7±0.7℃이 었다. 수온은 조간대 하부와 조하대 상부 간에는 유의한 차이 를 보이지 않았으나(P =0.297), 조사 시기 중 3월-8월이 9월-이 듬해 2월 보다 유의하게 높은 것으로 나타나(P<0.05) 온대 지 역의 특징을 잘 나타내었다. 광량 범위는 조간대 하부 중 수 온 상승기의 경우 25.8±1.0 mol photons m-2day-1, 수온 하강 기의 경우 27.1±1.0 mol photons m-2day-1이었고, 조하대 상 부 중 수온 상승기의 경우 12.6±0.5 mol photons m-2 day-1, 수 온 하강기의 경우 12.3±0.3 mol photons m-2 day-1이었다. 광 량은 조하대 상부가 조간대 하부의 생육지 보다 유의하게 낮. 통계분석 거머리말 자생지의 환경 특성, 잎과 지하경의 탄소와 질소 성 분, 단위 면적당 잎의 무게, 잎의 탄소와 질소 농도/단위 면적 당 잎의 무게와의 비 값은 조사 장소와 시기를 나누어서 통계 분석 하였다. 조사 장소는 조간대 하부와 조하대 상부로 나누. Table 1. Mean values and ranges of physical and chemical parameters in Seomjin Estuary on the south coast of the Korean peninsula from May 2003 to December 2004 (PIT: Period of increased temperature, PDT: Period of decreased temperature) Lower intertidal PIT Temperature (℃). PDT. Upper subtidal PIT. PDT. 19 .9±0.3. 18.3±0.7. 20.6±0.4. 18.7±0.7. Underwater irradiance (mol photons m day ). 25.8±1.0. 27.1± 1.0. 12.6±0.5. 12.3±0.3. Exposure time (h). 41.4±9.7. 43.4±6.8. 17.7±6.2. 12.9±3.4. -2. -1. Water column +. NH4 (μM) NO2-+NO3- (μM). PO4- (μM). Sediment pore water NH4+ (μM) NO2-+NO3- (μM) PO4- (μM). 4.2±0.3. 3.4±0.3. 3.8±0.3. 3.3±0.3. 11.5±1.2. 13.2±1.1. 5.5±0.5. 11.5±1.2. 1.1±0.1. 0.9±0.0. 1.2±0.1. 0.9±0.0. 227±10. 233±16. 229±9. 239±11. 11.5±1.7. 22.1± 2.8. 8.9±1.1. 10.8±1.5. 0.6±0.1. 2.0±0.3. 1.0±0.1. 1.5±0.1.
(4) 68. 김정배 . 박정임 . 이원찬 . 이근섭. 은 것으로 나타났고(P<0.001), 조사 시기는 유의한 차이가 없 었다(P =0.263). 노출시간은 조간대 하부 중 수온 상승기의 경 우 41.4±9.7(낮: 26.5±4.1, 밤: 14.9±5.9) h month-1, 수온 하강기 의 경우 43.4±6.8(낮: 15.9±1.9, 밤: 27.5±5.7) h month-1 이었고, 조하대 상부 중 수온 상승기의 경우 17.7±6.2(낮: 10.7±3.0, 밤: 7.0±3.6) h month-1, 수온 하강기의 경우 12.9±3.4(낮: 2.2±0.6, 밤: 10.7±3.1) h month-1 이었다. 노출 시간은 조간대 하부가 조하 대 상부의 생육지 보다 유의하게 높은 것으로 나타나(P<0.001) 조간대 하부는 노출에 의하여 해초 잎의 건조가 일어나며, 조사 시기는 유의한 차이가 없었다(P =0.841). 해수의 암모늄염 농도 범위는 조간대 하부 중 수온 상승기의 경우 4.2±0.3 μM, 수온 하강기의 경우 3.4±0.3 μM 이었고, 조 하대 상부 중 수온 상승기의 경우 3.8±0.3 μM, 수온 하강기의 경우 3.3±0.3 μM 이었다. 해수의 암모늄염 농도는 조사 정점 간에는 유의한 차이를 보이지 않았으나(P =0.431), 조사 시기 중 3월-8월이 9월-이듬해 2월 보다 유의하게 높은 것으로 나 타났다(P<0.05). 해수의 아질산염+질산염 농도 범위는 조간 대 하부 중 수온 상승기의 경우 11.5±1.2 μM, 수온 하강기의 경우 13.2±1.1 μM 이었고, 조하대 상부 중 수온 상승기의 경 우 5.5±0.5 μM, 수온 하강기의 경우 11.5±1.2 μM 이었다. 해 수의 아질산염+질산염 농도 범위는 조하대 상부가 조간대 하 부의 생육지 보다 유의하게 낮은 것으로 나타났고(P<0.001), 조사 시기 중 3월-8월이 9월-이듬해 2월 보다 유의하게 낮은 것으로 나타났다(P<0.001). 해수의 인산염 농도 범위는 조간 대 하부 중 수온 상승기의 경우 1.1±0.1 μM, 수온 하강기의 경 우 0.9±0.0 μM 이었고, 조하대 상부 중 수온 상승기의 경우 1.2±0.1 μM, 수온 하강기의 경우 0.9±0.0 μM 이었다. 해수의 인산염 농도 범위는 조사 정점간에는 유의한 차이를 보이지 않 았으나(P =0.613), 조사 시기 중 3월-8월이 9월-이듬해 2월 보 다 유의하게 낮은 것으로 나타났다(P<0.05). 퇴적물 공극수의 암모늄염 농도 범위는 조간대 하부 중 수온 상승기의 경우 227±10 μM, 수온 하강기의 경우 233±16 μM 이었고, 조하대 상부 중 수온 상승기의 경우 229±9 μM, 수온 하강기의 경우 239±11 μM 이었다. 퇴적물 공극수의 암모늄 염 농도 범위는 조사 정점간에 유의한 차이를 보이지 않았고(P =0.747), 조사 시기에 대해서도 유의한 차이를 보이지 않았다 (P =0.484). 퇴적물 공극수의 아질산염+질산염 농도 범위는 조 간대 하부 중 수온 상승기의 경우 11.5±1.7 μM, 수온 하강기의 경우 22.1±2.8 μM 이었고, 조하대 상부 중 수온 상승기의 경우 8.9±1.1μM, 수온 하강기의 경우 10.8±1.5 μM 이었다. 퇴적물 공극수의 아질산염+질산염 공극수의 농도 범위는 조하대 상부 가 조간대 하부의 생육지 보다 유의하게 낮은 것으로 나타났고 (P<0.001), 조사 시기 중 3월-8월이 9월-이듬해 2월 보다 유의 하게 낮은 것으로 나타났다(P<0.05). 퇴적물 공극수의 인산염 농도 범위는 조간대 하부 중 수온 상승기의 경우 0.6±0.1 μM, 수온 하강기의 경우 2.0±0.3 μM 이었고, 조하대 상부 중 수온. 상승기의 경우 1.0±0.1 μM, 수온 하강기의 경우 1.5±0.1 μM 이었다. 퇴적물의 인산염 농도 범위는 조사 정점간에는 유의한 차이를 보이지 않았으나(P =0.863), 조사 시기 중 3월-8월이 9 월-이듬해 2월 보다 유의하게 낮은 것으로 나타났다(P<0.001).. 거머리말의 탄소와 질소 성분비 거머리말 잎의 탄소 성분은 조간대 하부 중 수온 상승기의 경 우 34.6±0.2%, 수온 하강기의 경우 36.3±0.2%이었고, 조하 대 상부 중 수온 상승기의 경우 35.8±0.2%, 수온 하강기의 경 우 35.9±0.2%이었다(Fig. 2a). 거머리말 잎의 탄소 성분은 조 하대 상부가 조간대 하부 보다 유의하게 높은 것으로 나타났고 (P<0.05), 조사 시기 중 3월-8월이 9월-이듬해 2월 보다 유의 하게 낮은 것으로 나타났다(P<0.001). 거머리말 지하경의 탄소 성분은 조간대 하부 중 수온 상승기 의 경우 33.5±0.2%, 수온 하강기의 경우 34.9±0.1% 이었고, 조하대 상부 중 수온 상승기의 경우 35.3±0.4%, 수온 하강기의 경우 34.5±0.5%이었다(Fig. 2b). 거머리말 지하경의 탄소 성분 은 조하대 상부가 조간대 하부 보다 유의하게 높은 것으로 나타 났고(P<0.05), 조사 시기는 유의한 차이가 없었다(P=0.277). 거머리말 잎의 질소 성분은 조간대 하부 중 수온 상승기의 경우 2.9±0.1%, 수온 하강기의 경우 3.3±0.1%이었고, 조하대 상부 중 수온 상승기의 경우 3.0±0.1%, 수온 하강기의 경우 3.1±0.1%이었다(Fig. 3a). 거머리말 잎의 질소 성분은 조사 정 점간에는 유의한 차이를 보이지 않았고(P=0.120), 조사 시기 중 3월-8월이 9월-이듬해 2월 보다 유의하게 낮은 것으로 나타 났다(P<0.001). 거머리말 지하경의 질소 성분은 조간대 하부 중 수온 상승기 의 경우 2.4±0.1%, 수온 하강기의 경우 2.1±0.0%이었고, 조하 대 상부 중 수온 상승기의 경우 2.7±0.1%, 수온 하강기의 경우 2.5±0.1% 이었다(Fig. 3b). 거머리말 지하경의 질소 성분은 조 하대 상부가 조간대 하부 보다 유의하게 높은 것으로 나타났고 (P<0.05), 조사 시기 중 3월-8월이 9월-이듬해 2월 보다 유의 하게 낮은 것으로 나타났다(P<0.001). 거머리말 잎의 탄소와 질소 성분 비는 조간대 하부 중 수온 상 승기의 경우 12.0±0.2, 수온 하강기의 경우 11.1±0.2이었고, 조하대 상부 중 수온 상승기의 경우 12.5±0.3, 수온 하강기의 경우 12.0±0.3이었다(Fig. 4a). 거머리말 잎의 탄소와 질소 성 분 비는 조하대 상부가 조간대 하부 보다 유의하게 높은 것으로 나타났고(P<0.05), 조사 시기 중 3월-8월이 9월-이듬해 2월 보 다 유의하게 높은 것으로 나타났다(P<0.05). 거머리말 지하경의 탄소와 질소 성분 비는 조간대 하부 중 수 온 상승기의 경우 14.2±0.4, 수온 하강기의 경우 16.7±0.3이었 고, 조하대 상부 중 수온 상승기의 경우 13.5±0.3, 수온 하강 기의 경우 14.0±0.3% 이었다(Fig. 4b). 거머리말 지하경의 탄 소와 질소 성분 비는 조하대 상부가 조간대 하부 보다 유의하.
(5) 69. 조간대와 조하대 거머리말의 탄소 및 질소 성분 변화 42. Leaf C content (%). 40. (. ). M. J. (. ). M. J. Lower intertidal Upper subtidal. 38 36 34 32 30 28. Rhizome C content (%). 42 40. J. A. S. O. N. D. J. F. M. A. M. J. J. A. S. O. N. D. J. A S 2003. O. N. D. J. F. M. A. M. J J 2004. A. S. O. N. D. 38 36 34 32 30 28. Fig. 2. Carbon content of eelgrass Zostera marina leaf (a) and rhizome (b) tissues in Seomjin Estuary on the south coast of the Korean peninsula from May 2003 to December 2004. Values represent means±SE (n=6-10). 4.0. Leaf N content (%). 3.5. ). 3.0 2.5. Lower intertidal Upper subtidal. 2.0 1.5 4.0. Rhizome N content (%). (. M. J. (. ). M. J. J. A. S. O. N. D. J. F. M. A. M. J. J. A. S. O. N. D. J. A S 2003. O. N. D. J. F. M. A. M. J J 2004. A. S. O. N. D. 3.5 3.0 2.5 2.0 1.5. Fig. 3. Nitrogen content of eelgrass Zostera marina leaf (a) and rhizome (b) tissues in Seomjin Estuary on the south coast of the Korean peninsula from May 2003 to December 2004. Values represent means±SE (n=6-10)..
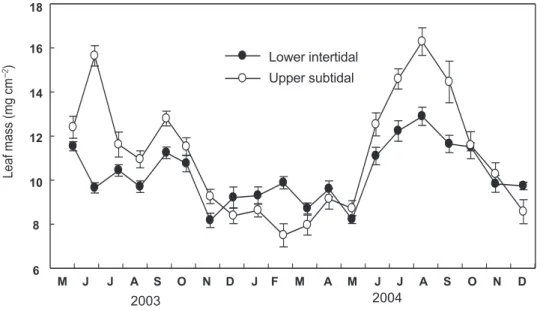
(6) 김정배 . 박정임 . 이원찬 . 이근섭. 70 20. Leaf C/N ratio. 18. (. ). M. J. (. ). M. J. Lower intertidal Upper subtidal. 16 14 12 10 8 J. A. S. O. N. D. J. F. M. A. M. J. J. A. S. O. N. D. J. A. S. O. N. D. J. F. M. A. M. J. J. A. S. O. N. D. 20. Rhizome C/N ratio. 18 16 14 12 10 8. 2003. 2004. Fig. 4. C/N ratio of eelgrass Zostera marina leaf (a) and rhizome (b) tissues in Seomjin Estuary on the south coast of the Korean peninsula from May 2003 to December 2004. Values represent means±SE (n=6-10). 게 낮은 것으로 나타났고(P<0.001), 조사 시기도 3월-8월이 9 월-이듬해 2월 보다 유의하게 낮은 것으로 나타났다(P<0.001). 거머리말 잎의 단위 면적당 무게는 조간대 하부 중 수온 상승 기의 경우 10.5±0.2 mg DW cm-2, 수온 하강기의 경우 10.1±0.1 mg DW cm-2이었고, 조하대 상부 중 수온 상승기의 경우 12.4±0.3 mg DW cm-2, 수온 하강기의 경우10.3±0.3 mg DW cm-2 이었다 (Fig. 5). 거머리말 잎의 단위 면적당 무게는 조하대 상부가 조간 대 하부 보다 유의하게 높은 것으로 나타났고(P<0.001), 조사 시 기 중 3월-8월이 9월-이듬해 2월 보다 유의하게 낮은 것으로 나 타났다(P<0.001). 거머리말 잎의 단위 면적당 무게와 탄소 성분은 조간대 하부 에서는 상관 관계가 없었으나(P=0.769, Fig. 6a), 조하대 상부 에서의 회귀 직선 식은 y=2.27x-70.36으로 상관 관계가 높았다 (r2=0.50, P<0.001, Fig. 6b). 거머리말 잎의 단위 면적당 무게와질 소 성분은 조간대 하부에서의 회귀 직선 식이 y= -2.62x +18.45 으로 상관관계가 높았다(r2=0.45, P<0.05, Fig. 7a). 조하대상부에서 의회귀직선식은y= -4.33x+24.2으로 상관관계가 높았다(r2=0.52, P<0.001, Fig. 7b).. 고. 찰. 거머리말 자생지의 환경특성 해초는 서식수심에 따라 분포하는 종이 다르거나, 같은 종에 대해서도 형태와 생리학적 현상이 다르게 나타난다(Harrison, 1982). 대부분의 해초는 건조에 견디지 못하거나, 조간대에 서 성장하지 않지만 거머리말속(Zostera sp.)의 Z. japonica, Z. marina, Z. noltii 및 Z. novazelandica 종들이 생장하고(Leuschner and Rees, 1993; Koch, 2001; Lee et al., 2006; Kim et al., 2009b) 있으며, 거머리말은 종종 물이 고여 있는 조수웅덩이나 수로 등에서 발견된다(Harrison, 1982; Thom, 1990). 조간대에 분포 하는 종의 하부제한은 포식과 경쟁에 의한 생물인자에 의하여 조절되는 반면 상부제한은 노출과 건조의 생리학적 내성에 의 하여 결정되고, 조간대의 수평분포패턴의 주요인자로서 작용 한다(Schonbeck and Norton, 1978, 1980; Shafer et al., 2007). 섬진강 하구의 갈사 갯벌이 있는 조간대에는 patchy형태로 분 포하는 애기거머리말과 거머리말이 있고, 조하대에는 거머리.
(7) 71. 조간대와 조하대 거머리말의 탄소 및 질소 성분 변화. 18. Leaf mass (mg cm-2). 16. Lower intertidal Upper subtidal. 14 12 10 8 6. M. J. J. A. S. O. N. D. J. F. M. A. M. 2003. J. J. 2004. A. S. O. N. D. Fig. 5. Area normalized leaf mass from the study sites in Seomjin Estuary on the south coast of the Korean peninsula from May 2003 to December 2004. Values represent means±SE (n=10).. Leaf mass Leaf C concent. 38 37. 10. 36 35. 5. 20. 34. M J. (. J. A S. O. N. D. J. F. M. A. M J. J. A. S. O. N D. ). 33 39 38. 15. 37. 10. 36 35. 5 0. Leaf C content (%). 39. ). 15. 0. Leaf mass (mg cm-2). (. 34. M. J. J. A S 2003. O. N. D. J. F M. A M. J J 2004. A. S. O. N. D. Leaf C content (%). Leaf mass (mg cm-2). 20. 33. Fig. 6. Area normalized leaf mass and leaf C content in lower intertidal (a) and upper subtidal (b) in Seomjin Estuary on the south coast of the Korean peninsula from May 2003 to December 2004. Values represent means±SE (n=6-10). 말이 조하대 2 m 수심까지 자생하였다(Kim et al., 2010). 섬진강 하구에 자생하고 있는 거머리말(Zostera marina)의 상부제한에 대해서 보면 수온이 점차 증가하여 해초의 형태학 적 특성과 생산성이 높아지는(Han et al., 2005) 시기인 3월에. 서 8월에 조간대 하부 노출 시간(41.4±9.7 h month-1)이 조하 대 상부 노출 시간(17.7±6.2 h month-1)보다 많았다. 조간대 하 부의 노출 시간 중에서 해초가 광합성을 할 수 있는 낮 시간이 전체 노출 시간의 64%를 차지하여 노출과 건조의 내성에 의.
(8) 김정배 . 박정임 . 이원찬 . 이근섭. 72. Leaf mass (mg cm-2). ( 15. 3.5. 10. 3.0. 5. 2.5. 0 20. Leaf mass (mg cm-2). ). M. J. (. ). J. A. S. O. N. D. J. F. M. A. M. J. J. A. S. O. N. D. 2.0 4.0. 15. 3.5. 10. 3.0. 5. 2.5. 0. M. J. J. A. S. O. N. D. J. F. 2003. M. A. M. J. J. A. S. O. N. D. Leaf N content (%). 4.0. Leaf mass Leaf N concent. Leaf N content (%). 20. 2.0. 2004. Fig. 7. Area normalized leaf mass and leaf N content in lower intertidal (a) and upper subtidal (b) in Seomjin Estuary on the south coast of the Korean peninsula from May 2003 to December 2004. Values represent means±SE (n=6-10). 하여 생육하고 있는 것으로 판단된다(Schonbeck and Norton, 1978, 1980; Harrison, 1982; Thom, 1990; Leuschner et al., 1998; Shafer et al., 2007). 이러한, 노출과 건조의 내성에 의하 여 조간대 하부 해초는 생존뿐만 아니라 재생산도 이루지는 것 으로 판단된다(Han et al., 2005; Shafer et al., 2007; Kahn and Durako, 2009). 해초는 높은 생산성에 필요한 많은 양의 영양염류를 필요로 하며, 지상부와 지하부의 질소 고정 율은 51대 49로서 잎을 통해 해수에서, 그리고 지하경과 뿌리에서 퇴적물 공극수 내 의 영양염류를 흡수한다(Short and McRoy, 1984; Ward et al., 1984; Fonseca and Fisher, 1986; Lee and Dunton, 1999b). 해 초가 이용할 수 있는 무기질소는 해수에 용존된 NO3-와 NH4+ 그리고 공극수에 포함된 NH4+ 이다(Lee et al., 2007). 해수중 의 영양염은 플랑크톤의 대 발생에 의한 흡수와 빛의 차단을 통 해 해초지 소멸의 원인이 되기도 한다(Burkholder et al., 1992, 1994; Orth and Moore, 1983; Silberstein et al., 1986; Giesen et al., 1990; Tomasko and Lapointe, 1991). 퇴적물 공극수의 영양염은 해초의 지하경을 통한 영양염 공 급원으로 해초의 생장에 밀접한 영향을 미친다(Iizumi and Hattori, 1982; Short and McRoy, 1984; Zimmerman et al., 1987). 거머리말이 생장을 하기 위해서는 높은 무기 영양염류. 가 요구되고 있으나, 우리나라 거머리말 생육지의 퇴적물 공극 수 영양염의 농도는 계절에 따른 생장을 제한 하지 않을 정도로 풍부한 것으로 나타났다(Lee, 2004). 퇴적물 공극수의 무기 영 양염류 농도는 공간적인 변위가 매우 높고, 일시적인 변동으로 계절적 경향은 보이지 않았다(Czerny and Dunton, 1995; Udy and Dennison, 1997; Lee and Dunton, 2000; Lee et al., 2007).. 거머리말 조직 내의 탄소, 질소 성분 해수중의 무기 영양염류 흡수를 보면 조하대 상부 해초의 경 우 잎으로 무기 영양염류를 흡수하며, 조간대 하부 해초가 물 에 잠겨 있을 때에는 조하대 상부와 같은 기작으로 무기영양 염류를 흡수 한다. 그러나, 썰물로 인하여 물이 빠지면 거머 리말 잎은 충분히 물에 잠기는 것이 아니라 몇 센티미터 깊이 의 조수웅덩이에 잎이 드리워진다. 이때 조수웅덩이 속에 있 던 암모늄염과 질산염의 감소와 더불어 퇴적물속에 있던 암모 늄염의 용출이 일어난다(Hemminga, 1994). 용출된 암모늄염 은 조수 웅덩이의 암모늄염이 증가되어 거머리말이 잎의 표면 을 통하여 퇴적물 공극수 중의 암모늄염을 직접적으로 이용한 다(Hemminga, 1994). 독일의 Wadden Sea에서 거머리말속의 분포형태를 보면 조 간대에는 Z. noltii와 Z. marina가 혼합되어 생육하고 있고,.
(9) 73. 조간대와 조하대 거머리말의 탄소 및 질소 성분 변화. 조하대에는 Z. marina가 생육하고 있다(Leuschner and Rees, 1993). Leuscher et al. (1998)에 의하면 조간대에 생육하고 있 는 Z. noltii와 Z. marina 중에서 Z. noltii보다 Z. marina의 잎 에서 건조에 더 민감하다고 하였고, 조간대에 생육하는 해초 가 건조에 의하여 탄소 동화 작용이 제약을 받는다고 하였다. Shafer et al. (2007)에 의하면 조간대에서 거머리말의 수분함 량이 15%에서는 광합성을 전혀 할 수 없었고, 수분 함량이 32%에서는 100%의 수분함량의 광합성 효율의 0.79를 가지고 있었으며, 수분 함량이 53% 이상이 되어야 100%의 수분 함량 과 같은 광합성 효율을 가지는 것으로 나타났다. 그러므로 조 간대 하부에 자생하고 있는 거머리말은 조하대 상부에 자생하 고 있는 거머리말 보다 건조에 의하여 광합성 효율이 낮을 것 으로 판단된다. 대부분 해초의 생장 제한 요인은 질소와 인에 의해서 일어 나지만, 탄소가 해초 조직 건 중량의 30-40%를 차지하고, 질 소 및 인은 각각 1-4% 및 0.1-1.0%를 차지하여 탄소 값의 변동 이 질소 및 인보다도 매우 낮을 경우 상대적으로 낮은 탄소 값 의 변동으로 인하여 해초의 생장 제한 요인으로 작용할 수 있 다(Short et al, 1985; Duarte, 1990; Lee and Dunton, 1999a, 2000; Lee, 2004; Lee et al., 2005). 거머리말 잎의 탄소와 질 소 성분 비는 조하대에서 잎은 12.2±0.5, 지하경은 13.8±0.5이 었고, 조간대에서 잎은 11.6±0.5, 지하경은 15.4±0.8로서 조간 대에서 잎과 지하경의 탄소/질소비가 크게 나타났다. 탄소/질 소비가 조하대에서는 잎이 증가하면 지하경은 감소하고, 잎이 감소하면 지하경은 증가 하였으나(Lee and Dunton, 1999a), 조 간대에서는 이러한 경향이 나타나지 않았다. 또한, 거머리말 이 해수 중에 존재하는 중탄산 이온을 선호하기 때문에 탄소 성 분이 조하대 보다 조간대에서 광합성을 제한하는 요인으로 판 단된다(Sand-Jensen and Gordon, 1984; Beer and Rehnberg, 1997; Invers et al., 2001). 섬진강 하구의 조하대 상부에서는 거머리말 잎의 질소 성분 이 수온이 점차 증가하여 잎으로의 생장이 활발한 시기인 3월 부터 8월 까지는 성분이 옅어지고, 9월부터 이듬해 2월 까지 는 증가하는 패턴으로 나타났으나, 조간대 하부에서는 이러한 경향을 볼 수 없었다(Lee et al., 2005). 해초의 지하경은 뿌리 를 통하여 퇴적물 공극수의 영양염류를 흡수하기도 하지만 광 합성에 의해 생성된 탄소를 저장하는 역할을 한다. 지하경에 저장된 탄소는 호흡을 하거나 오래된 잎이 떨어지고 새로운 잎 의 재생과 같은 잎에서의 탄소 생산을 보충하기 위하여 조하 대 상부에서는 10월부터 이듬해 3월까지 지하경의 탄소가 잎 으로 전이되어 지하경의 탄소 성분이 낮아 지고 광합성이 활발 한 4월에서 9월까지 지하경의 탄소 성분이 증가하였다(Dawes and Lawrence 1979; Lee and Dunton, 1996; Lee and Dunton, 1999a). 여기에서 잎에서의 질소 성분 증가와는 다르게 지하경 의 탄소성분이 4월에서 9월까지 증가하는 1개월의 시기적인 차이를 보였는데, 이것은 잎이 광합성에 의하여 탄소 성분을 지하경에 저장하는 시간적 차이 때문에 발생 한 것으로 판단된. 다. 또한, 섬진강 하구의 조간대 하부에 생육하는 거머리말이 탄소와 질소 성분에 대하여 계절경향이 잘 표현되지 않은 것은 노출에 의한 건조 스트레스의 영향인 것으로 판단된다.. 사. 사. 본 연구는 국립수산과학원(RP-2011-ME-022)의 지원에 의 해 운영되었습니다. 조사 및 분석에 도움을 주신 남서해수산연 구소 소장님, 자원환경과 및 해역산업과 박사님들께 감사의 말 씀을 드립니다. 무명의 심사자의 지적에 의해 본 논문의 질이 향상되었으며 이에 깊이 감사 드립니다. . 참고문헌. Beer S and Rehnberg J. 1997. The acquisition of inorganic carbon by the seagrass Zostera marina. Aquat Bot 56, 277-283. Burkholder JM, Mason KM and Glasgow Jr HB. 1992. Watercolumn nitrate enrichment promotes decline of eelgrass Zostera marina: evidence from seasonal mesocosm experiments. Mar Ecol Prog Ser 81, 163-178. Burkholder JM, Glasgow Jr HB and Cooke JE. 1994. Comparative effects of water-column nitrate enrichment on eelgrass Zostera marina, shoalgrass Halodule wrightii, and widgeongrass Ruppia maritima. Mar Ecol Prog Ser 105, 121-138. Cultural Heritage Administration. 2006. The investigation research on the actual condition of Brent Geese wintering ground of the Galsa area in the estuary of the Seomjin River. Bird Conservation Society of Korea, Chungnamkihoik, Korea, 59. Czerny AB and Dunton KH. 1995. The effects of in situ light reduction on the growth of two subtropical seagrasses, Thalassia testudinum and Halodule wrightii. Estuaries 18, 418-427. Dawes CJ and Lawrence JM. 1979. Effects of blade removal on the proximate composition of the rhizome of the seagrass Thalassia testudinum Banks ex König. Aquat Bot 7, 255-266. Day JW, Hall CAS, Kemp WM and Alejandro Y. 1989. Estuarine Ecology. John Wiley and Sons Inc., New York, Chichester, Brisbane, Toronto, Sigapore, 558. den Hartog C. 1970. The seagrass of the world. North-Holland Publishing Company, Amsterdam, Netherlands, 275. Dring MJ. 1982. The biology of marine plants. Cambridge University Press, Cambridge, UK, 199. Duarte CM. 1990. Seagrass nutrient content. Mar Ecol Prog Ser 67, 201-207. Fonseca MS and Fisher JS. 1986. A comparison of canopy friction and sediment movement between four species of seagrass with reference to their ecology and restoration..
(10) 74. 김정배 . 박정임 . 이원찬 . 이근섭. Mar Ecol Prog Ser 29, 15-22. Giesen WBJT, van Katwijk MM and den Hartog C. 1990. Eelgrass condition and turbidity in the Dutch Wadden Sea. Aquat Bot 37, 71-85. Harrison PG. 1982. Comparative growth of Zostera japonica Aschers & Graebn and Z. marina L. under simulated intertidal and subtidal conditions. Aquat Bot 14, 373-379. Han S-J, Jung CK, Kim JB, Yoon S-J, Kim CW, Hwang H-K, Lee SG, Yu J and Choi YH. 2005. Research of seagrass restoration in the southern coast of Korea. In: Technical Report of National Fisheries Research and Development Institute, NFRDI, Guduk, Korea, 485-498. Hemminga MA, Koutstaal BP, van Soelen J and Merks AGA. 1994. The nitrogen supply to intertidal eelgrass (Zostera marina). Mar Biol 118, 223-227. Iizumi H and Hattori A. 1982. Growth and organic production of eelgrass(Zostera marina L.) in temperate waters of the Pacific coast of Japan. III. The kinetics of nitrogen uptake. Aquat Bot 12, 245-256. Invers O, Zimmerman RC, Alberte RS, Pérez M and Romero J. 2001. Inorganic carbon sources for seagrass phytosynthesis: an experimental evaluation of bicarbonate use in species inhabiting temperate waters. J Exp Mar Biol Ecol 265, 203-217. Kahn AE and Durako MJ. 2009. Photosynthetic tolerances to desiccation of the co-occurring seagrasses Halophila johnsonii and Halophila decipiens. Aquat Bot 90, 195198. Kim JB. 2012. Distribution and ecological characteristics of seagrasses in the Deukryang Bay and Seomjin Estuary and taxonomy of an unrecorded species from Korean flora. Ph. D. Thesis, University of Pusan, Busan, Korea. Kim JB, Park J-I, Jung C-S, Lee P-Y and Lee K-S. 2009a. Distributional range extension of the seagrass Halophila nipponica into coastal waters off the Korean peninsula. Aquat Bot 90, 269-272. Kim JB, Park J-I and Lee K-S. 2009b. Seagrass distribution in Deukryang Bay. Kor J Fish Aquat Sci 42, 509-517. Kim JB, Ryu JH and Kim JK. 2009c. Comparative analysis of fish community structure between eelgrass (Zostera marina L.) beds and an adjacent unvegetated area in southern Korea. J Fish Sci Technol 12, 60-69. Kim JB, Park J-I, Choi W-J, Lee JS and Lee K-S. 2010. Spatial distribution and ecological characteristics of Zostera marina and Zostera japonica in the Seomjin Estuary. Kor J Fish Aquat Sci 43, 351-361. Koch EW. 2001. Beyond light: physical, geological, and geochemical parameters as possible submersed aquatic vegetation habitat requirements. Estuaries 24, 1-17. Lee K-S. 2004. Development of indicator for coastal and estuarine eutrophication using morphological characteristics and tissue N content of eelgrass, Zostera marina.. Algae 19, 129-137. Lee K-S and Dunton KH. 1996. Production and carbon reserve dynamics of the seagrass Thalassia testudinum in Corpus christi Bay, Texas, USA. Mar Ecol Prog Ser 143, 201-210. Lee K-S and Dunton KH. 1997. Effects of in situ light reduction on the maintenance, growth and partitioning of carbon resources In Thalassia testudinum Banks ex König. J Exp Mar Biol Ecol 21, 53-73. Lee K-S and Dunton KH. 1999a. Influence of sediment nitrogen availability on carbon and nitrogen dynamics in the seagrass Thalassia testudinum. Mar Biol 134, 217-226. Lee K-S and Dunton KH. 1999b. Inorganic nitrogen acquisition in the seagrass Thalassia testudium: development of a whole plant nitrogen budget. Limnol Oceanogr 44, 1204-1215. Lee K-S and Dunton KH. 2000. Effects of nitrogen enrichment on biomass allocation, growth, and leaf morphology of the seagrass Thalassia testudium. Mar Ecol Prog Ser 196, 39-48. Lee K-S, Kang C-K and Kim Y-S. 2003. Seasonal dynamics of the seagrass Zostera marina on the south coast of the Korean penunsula. J Korean Soc Oceanogr 38, 68-79. Lee K-S, Park SR and Kim JB. 2005. Production dynamics of the eelgrass, Zostera marina in two systems on the south coast of the Korea peninsula. Mar Biol 147, 1091-1108. Lee K-S, Park SR and Kim YK. 2007. Effects of irradiance, temperature, and nutrients on growth dynamics of seagrass: A review. J Exp Mar Biol Ecol 350, 144-175. Lee K-S, Short FT and Burdick DM. 2004. Development of a nutrient pollution indicator using the seagrass, Zostera marina, along nutrient gradients in three New England estuaries. Aquat Bot 78, 197-216. Lee SY, Kim JB and Lee SM. 2006. Temporal dynamics of subtidal Zostera marina and intertidal Zostera japonica on the southern coast of Korea. Mar Ecol 27, 133-144. Leuschner C and Rees U. 1993. CO2 gas exchange of two intertidal seagrass species, Zostera marina L. and Zostera noltii Hornem., during emersion. Aquat Bot 45, 53-62. Leuschner C, Landwehr S and Mehlig U. 1998. Limitation of carbon assimilation of intertidal Zostera noltii and Z. marina by desiccation at low tide. Aquat Bot 62, 171-176 Marbà N, Holmer M, Gacia E and Barrón C. 2006. Seagrass beds and coastal biogeochemistry. In: Seagrasses: Biology, Ecology and Conservation. Larkum AWD, Orth RJ and Duarte CM, eds. Springer, Dordrecht, Netherlands, 135-157. MLTM. 2009. Establishment of action plans for management of estuary system(II), Youngsan Estuary, Korea, 619. MLTM. 2010a. Establishment of action plans for management of estuary system(III-1), Seomjin Estuary, Korea, 419..
(11) 조간대와 조하대 거머리말의 탄소 및 질소 성분 변화. MLTM. 2010b. Establishment of action plans for management of estuary system(III-2), Seomjin Estuary, Korea, 284. MOMAF. 2002. Standard Methods of Marine Environment. Korea, 330. Orth RJ and Moore KA. 1983. Chesapeake Bay: an unprecedented decline in submerged aquatic vegetation. Science 222, 51-53. Romero J, Lee K-S, Pérez M, Mateo MA and Alcoverro T. 2006. Nutrients dynamics in seagrass ecosystems. In: Seagrasses: Biology, Ecology and Conservation, Larkum AWD, Orth RJ and Duarte CM, eds. Springer Press, Dordrecht, Netherlands, 227-254. Sand-Jensen K and Gordon DM. 1984. Differential ability of marine and freshwater macrophytes to utilize HCO3- and CO2. Mar Biol 80, 247-253. Sand-Jensen K, Binzer T and Middelboe AL. 2007. Scaling of photosynthetic production of aquatic macrophytes: A review. Oikos 116, 280-294. Schonbeck M and Norton TA. 1978. Factors controlling the upper limits of fucoid algae on the shore. J Exp Mar Biol Ecol 31, 303-313. Schonbeck M and Norton TA. 1980. Factors controlling the lower limits of fucoid algae on the shore. J Exp Mar Biol Ecol 43, 131-150. Shafer DJ, Sherman TD and Wyllie-Echeverria S. 2007. Do desiccation tolerances control the vertical distribution of intertidal seagrasses? Aquat Bot 87, 161-166. Short FT, Davis MW, Gibson RA and Zimmermann CF. 1985. Evidence for Phosphorus Limitation in Carbonate Sediments of the Seagrass Syringodium filiforme. Estuar Coast Shelf Sci 20, 419-430. Short FT and McRoy CP. 1984. Nitrogen uptake by leaves and roots of the seagrass Zostera marina L. Bot Mar 27,. 75. 547-555. Silberstein K, Chiffings AW and McComb AJ. 1986. The loss of seagrass in Cockburn Sound, Western Australia. III. The effect of epiphytes on productivity of Posidonia australis Hook. F. Aquat Bot 24, 355-371. Thom RM. 1990. Spatial and temporal patterns in plant standing stock and primary production in a temperate seagrass system. Bot Mar 33, 497-510. Tomasko DA and Lapointe BE. 1991. Productivity and biomass of Thalassia testudinum as related to water column nutrient availability and epiphyte levels: field observations and experimental studies. Mar Ecol Prog Ser 75, 9-17. Udy JW and Dennison WC. 1997. Growth and physiological responses of three seagrass species to elevated sediment nutrients in Moreton Bay, Australia. J Exp Mar Biol Ecol 217, 253-277. Ward LG, Kemp WM and Boynton WR. 1984. The influence of waves and seagrass communities on suspended particulates in an estuarine embayment. Mar Geol 59, 85-103. Yoo S-H. 2007. Measurement of the environmental value of the Seomjin-river estuary. J Environmental Policy 6, 1-25. Yoon S-P. 2006. The eelgrass, Zostera marina and its associated macroinvertebrate communities in Gwangyang Bay. Korea. Ph. D. Thesis, University of Inha, Incheon, Korea. Zimmerman RC, Smith RD and Alberte RS. 1987. Is growth of eelgrass nitrogen limited? A numerical simulation of the effects of light and nitrogen on the growth dynamics of Zostera marina. Mar Ecol Prog Ser 41, 167-176. 2011년 8월 10일 접수 2011년 11월 12일 수정 2012년 2월 8일 수리.
(12)
수치



+2
관련 문서
따라서 바르게
세계물동량의 감소로 세계 해운시장이 불황을 겪고 있을 때 가장 먼저 해양플랜트 건조 시장에 뛰어 들어 지금까지 해양플랜트 건조 부문 세계 1위를 유지하고
즉 사각형 ABCD가
③ 분자가 반응하기 위해서는 반드시 충돌이 필요하며 그 충돌은 분자가 활성화에너지 이상 의 에너지를 갖고 화학적 반응에 필요한 방향이 알맞을 때 반응을 일으킬
따라서 계산
7 Crysis Warhead FPS Crytek/Electronic Arts 8 The Sims 2 Double Deluxe Simulation Electronic Arts 9 World of Warcraft MMORPG Blizzard 10 Spore Creature Creator
또는 해당 약제 투여 관련
[r]
