www.earticle.net
소의 정원세포에서 Matrix Metalloproteinase-9(MMP-9) 발현 양상 구명
곽태일1· 이원영2· 정하연3· 이승윤4· 송 혁1*
건국대학교 줄기세포재생생물학과1, 건국대학교 식품생명과학부 동물성소재전공2, 국립축산과학원 낙농과3, 한별팜텍4
Matrix Metalloproteinase-9 (MMP-9) Expression Pattern in Bovine Testes
Tae Il Kwak1, Won Young Lee2, Ha Yeon Jung3, Seung Yoon Lee4 and Hyuk Song1*
1Department of Stem Cells and Regenerative Bioology, Konkuk University, Seoul 05029, Korea,
2Department of Food Bioscience, Konkuk University, Chungju 27478, Korea,
3Dairy Science Division, National Institute of Animal Science, RDA, Cheonan 31000, Korea,
4HanByol Farm Tech (Swine Consulting Group), Gyeonggi 12259, Korea
ABSTRACT1)
Identification of specific marker proteins in cells is useful for isolating cells and determining their cellular characteristics and functions. Based on our previous study showing that matrix metalloproteinase 9 (MMP-9) can be used as a marker for porcine spermatogonia, the expression pattern of MMP-9 was determined in both pre- (5-month old) and post-pubertal (11–month old) bovine testes. Histological analysis revealed that spermatogonia were located near the basement membrane in both testes, while spermatozoa were not detected in the 5-month old pre-pubertal bovine testes and epididymides. Mature spermatozoa were observed in the 11-month bovine testes and epididymides, and MMP-9 expression in 11-month old bovine testes was lower than 5-month old testes, according to reverse transcription-PCR and real-time-PCR data. To determine the specific expression sites of MMP-9 in the bovine testes, immunohistochemistry was performed. Expression of MMP-9 was observed in cells near the basement membrane of seminiferous tubules in both 5- and 11-month old testes. Furthermore, MMP-9 positive cells expressed protein gene product 9.5 (PGP9.5) and deleted in azoospermia (DAZL) that are already known as bovine spermatogonial stem cells markers. In the present study, MMP-9 expression was identified in both pre- and post-pubertal bovine spermatogonia expressing PGP9.5 and DAZL, and located near the basement membrane of seminiferous tubules. Thus, MMP-9 can be used as a marker for bovine spermatogonia, and may provide useful platforms for understanding the interaction between germ cells and extracellular matrix during spermatogenesis in the seminiferous tubules.
(Key words: Bovine, Spermatogonia, MMP-9, PGP9.5, DAZL)
* Corresponding author: Hyuk Song, Department of Stem Cells and Regenerative Biology, Konkuk University, Seoul 05029, Korea. Tel: +82-2-450-0562, E-mail; songh@konkuk.ac.kr
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc-nd/3.0/deed.ko), which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. The moral rights of the named author(s) have been asserted.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:54 PM
www.earticle.net
Ⅰ. 서론
조직 내 특정 세포에서 특이적으로 발현하는 단백질 표 지인자를 발굴하는 것은 세포의 특성과 기능을 구명하고 그 세포를 분리하는데 유용하게 이용된다. 정소 내 곡세정 관의 세포 중 정원세포를 확인하고 분리하는 것은 정자형 성과정의 이해와 체외배양 체계를 활용한 생식세포 연구 의 가장 필요한 단계이며, 분리된 웅성 생식세포를 공여 받은 수컷 정소에서 이식된 생식세포 유래 정자를 생산하 기도 하였다(Stockwell et al., 2008). 현재까지 소 정원세포 의 표지인자로 알려진 단백질은 THY1, DBA, protein gene product(PGP)9.5, DAZL 등이다(Reding et al., 2010;
Izadyar et al., 2002; Wrobel et al., 1995; Shah et al., 2015). 이러한 표지 단백질의 발굴에도 불구하고 소 정원세 포의 특성이나 정자형성과정을 연구하는데 있어서 더욱 다양한 표지 단백질의 발견이 필요한 상황이다.
Matrix metalloproteinase(MMP)-9는 MMP 단백질 집단 의 하나로서 세포외기질 물질과 결합조직의 기저막 물질 을 분해하는 단백질로 알려져 있다(Woessner et al., 1991).
특히, MMP-9는 정소의 곡세정관 내 세포외기질 구성물질 인 제 4형 콜라젠(type IV collagen)과 라미닌(laminin)을 분해하는 효소이다(Heikinheimo et al., 1995; Gu et al., 2005). 정소와 관련된 기능적인 측면에서 MMP-9는 정소 내 sertoli 세포의 치밀결합 구조 형성과 정소발달에 필요 한 세포외기질 재형성에 중요한 역할을 하는 것으로 밝혀 졌다(Robinson et al., 2001). 또한, MMP-9 유전자 녹아웃 생쥐의 경우 번식 장애를 보였고(Dubois et al., 2000), MMP-9는 근육양세포(myoid cell)의 이동에 밀접하게 연 관된다고 보고되었다(Moreno-Ruiz et al., 2016).
정소 내에서 MMP-9의 발현양상은 일부 포유류에서 보고 되고 있다. 쥐(rat)에서 MMP-9는 정모세포(spermatocytes) 와 정세포(spermatids)에서 발현이 확인 되었으며(Siu et al., 2003; Lydka et al., 2012), 개의 정소에서는 정모세포와 원형정자세포(round spermatids)에서 MMP-9가 발현한다 고 보고되었으나, 정원세포에서 발현은 나타나지 않았다 (Warinrak et al., 2015). 돼지 정소에서 MMP-9의 발현은 체내 정소조직의 정원세포와 체외 배양된 정원줄기세포에 서 확인 되었다(Lee et al., 2016). 하지만 현재까지 소의 정 소에서 MMP-9의 발현과 관련된 보고는 전무한 실정이다.
이전의 선행연구를 토대로 본 연구에서는 MMP-9가 소 의 정소 내 생식세포에서 발현 할 수 있고, 생식세포의 이 동에 밀접한 영향을 줄 수 있을 것이라는 가설을 세웠다.
이 가설을 증명하기 위해 성 성숙 이전과 이후의 소 정소 를 이용해 각각의 정소에서 MMP-9의 발현양상과 MMP-9 이 특이적으로 발현하는 세포를 구명하고자 한다.
Ⅱ. 재료 및 방법
1. 정소 조직 준비
본 실험에 사용된 정소는 충청북도 청주시 인근 농장에 서 사육된 5, 11 개월령 한우로부터 거세된 정소를 구하여 사용하였다. 거세된 소 정소는 1cm×1cm 크기로 절편한 뒤 급속 냉동 후 -70℃에 보관하고 조직 분석을 위해 Bouin’s solution(Sigma-Aldrich, St. Louis, MO, USA) 에 고정하였다.
2. 유전자 발현 분석
Qiagen RNA extraction kit(Qiagen, Venlo, Netherlands) 을 이용하여 5, 11 개월령 소 정소로부터 RNA를 회수한 후 1ug 총 RNA를 RT-PCR premix kit(Intron Biotechnology, Korea)을 이용하여 cDNA 로 합성 하였다. PCR 반응은 9 5℃ 20초, 58℃ 20초, 72℃ 20초 조건으로 35회 반응 시켰 으며, AccuPower PCR PreMix kit(BIONEER, Daejeon, South Korea)을 사용하였다.
RNA의 정량은 iQ5 Multicolor Real-Time PCR Detection System(Bio-Rad Laboratories, Hercules, CA, USA)을 이용하여 분석하였으며, 반은 조건은 반응은 95℃
20초, 58℃ 20초, 72℃ 20초 조건으로 40회 반응 시켜 CT 값을 GAPDH 로 정량화 하였다. 발달단계별 정소에서의 mRNA 발현은 5개월 소 정소에서의 발현을 기순으로 상 대 정량 하였다. RT-PCR과 Real-time PCR에 사용한 Primer는 동일하며 Table1에 정리하였다.
3. 조직 Hematoxylin & Eosin 염색 분석
Bouin’s solution에 의해 24시간 고정된 정소 조직을 70-100%(v/v) 에틸 알코올에 처리 하여 탈수 시킨 뒤, xylene에 10분간 2회 처리하여 투명화 시켰다. 투명화된 소 정소 조직은 파라핀 침투 후 고정하여 6마이크로미터 두께로 microtome(Thermo Scientific, Waltham, MA, USA)을 이용해 절편 하였다. 절편 한 소 정소 조직은 slide glass 위에서 건조되었다.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:54 PM
www.earticle.net
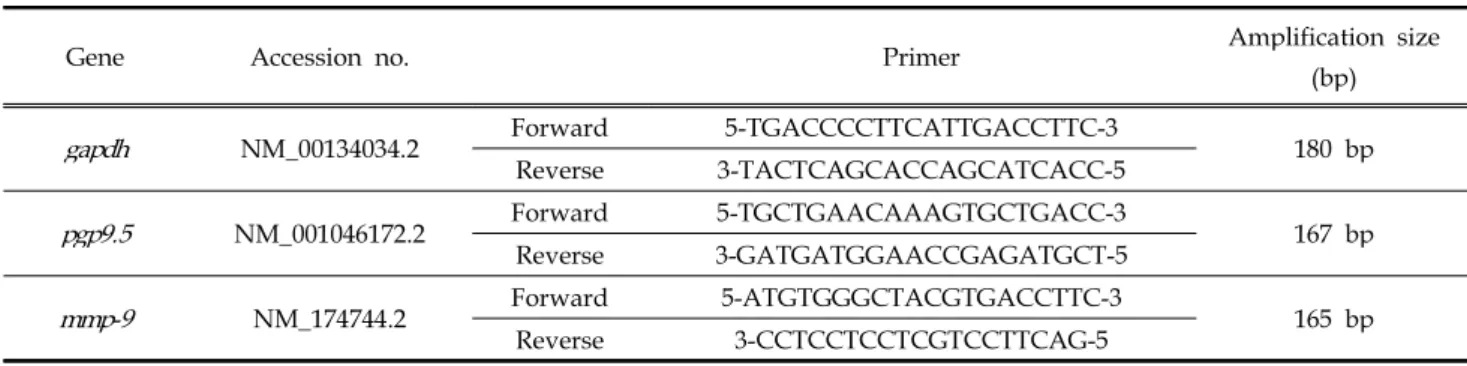
Table 1. Bovine primer sequence used in RT-PCR and real-time PCR.
Slide glass 에 건조된 소 조직은 xylene에 10분 처리 후 100% 에서 50%(v/v) 에틸 알코올에 침수 시킨 뒤 hematoxylin (Sigma-Aldrich) 에 5분간 처리하고 3회 수세 후 eosin (Sigma, St. Louis, MO, USA)에 1분간 처리하였다. 염색된 조직은 DPX mounting medium(Fluka, Buchs, Switzerland) 을 이용하여 고정한 후 광학 현미경(Nikon, Tokyo, Japan) 을 이용하여 분석하였다.
4. 조직 면역염색 분석
침수된 조직은 tris-ethylenediamine tetra acetic acid (EDTA) solution에 20분간 끓여서 항원을 복구시켰다. 슬 라이드가 실온이 되었을 때 2% Bovine serum albumin (Bovogen Biologicals, Melbourne, Australia), 0.025%
Triton-X100 을 포함하는 PBS 를 1시간 처리하여 세포막의 투과성을 높이고 비 특이적인 결합이 일어나지 않도록 하 였다. 이 후 같은 PBS 용액에 1:100 로 희석된 MMP-9 항 체(Santa Cruz Biotechnology, Santa Cruz, CA, USA), 1:500 으로 희석된 PGP9.5 항체(AbD Serotec, Raleigh, NC, USA), 1:100으로 희석된 DAZL 항체(goat polyclonal IgG, Santa Cruz Biotechnology)를 처리한 후 3회 수세 후 1:1000로 희석된 AlexaFluor®568donkeyanti-rabbitIgG (LifeTechnologies, Carlsbad, CA, USA), AlexaFluor® 488rabbit anti-goatIgG(LifeTechnologies)와 AlexaFluor® 488goat anti-mouseIgG(LifeTechnologies)를 2차 항체로 사 용하였다. 1차 항체는 4°C 에서 12시간 반응 시켰으며 2차 항체 는 상온에서 1시간 반응 시켰다. 4′,6-diamidino-2-phenylindol (DAPI; Sigma-Aldrich) 를 상온에서 10분간 처리하여 핵을 염색하였고 형광 현미경(Nikon) 을 이용해 단백질 발현을 관찰하였다.
5. 통계 분석
5개월과 11개월령 각 4마리의 소 정소가 사용 되었으며 3회 이상 반복하여 모든 실험을 진행 하였다. PGP9.5 와 MMP-9의 발현은 unpaired t-test 로 통계 분석 하였으며 유의성은 p<0.05로 분석 되었다.
Ⅲ. 결과
1. 소 정소와 정소상체의 조직학적 분석
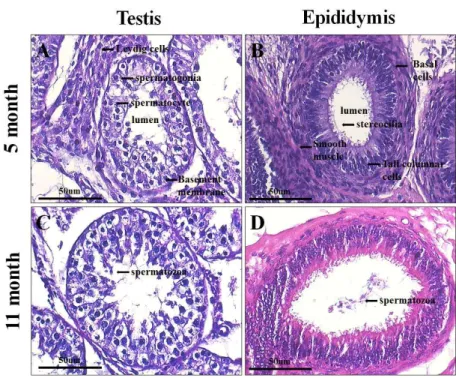
소 정소 내 생식세포들의 발달단계를 확인하기 위하여 5개월 령(성 성숙 전)과 11개월 령(성 성숙 후) 소 정소와 정소상체의 조직학적 분석을 수행하였다. 두 정소 모두 성 숙된 곡세정관 형태를 보여주고 있었다. 5개월 령 정소의 곡세정관 내에는 sertoli 세포가 분포하고 있으며, 정원세 포와, 발달된 정모세포가 관찰되었으나, 정자세포나 성숙 된 정자는 관찰되지 않았다(Fig. 1A). 11개월 령 정소의 곡 세정관 내에는 sertoli 세포, 정원세포, 정모세포, 원형정자 세포, 성숙된 정자 등 성 성숙 이후 볼 수 있는 모든 단계 의 생식세포가 관찰되었다(Fig. 1B). 또한 5개월 령 정소상 체에서 볼 수 없었던 성숙된 정자는 11개월 령 정소상체에 서 저장되어 있는 것이 관찰되었다(Fig. 1C, D).
2. 소 정소에서
mmp-9
유전자 발현 구명소 정소 내에 mmp-9 유전자의 발현 유무를 확인하기 위 하여,mmp-9와 소 정원세포의 표지인자로 알려진 pgp9.5의 발현 양상을 RT-PCR과 real-time PCR 기법을 이용해 분
Gene Accession no. Primer Amplification size
(bp) gapdh NM_00134034.2 Forward 5-TGACCCCTTCATTGACCTTC-3
180 bp Reverse 3-TACTCAGCACCAGCATCACC-5
pgp9.5 NM_001046172.2 Forward 5-TGCTGAACAAAGTGCTGACC-3
167 bp Reverse 3-GATGATGGAACCGAGATGCT-5
mmp-9 NM_174744.2 Forward 5-ATGTGGGCTACGTGACCTTC-3
165 bp
Reverse 3-CCTCCTCCTCGTCCTTCAG-5
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:54 PM
www.earticle.net
Fig. 1. Histological analysis of the bovine testis and epididymis by hematoxylin and eosin staining. 5 month old bovine (A) testis (B) epididymis. 11 month old bovine (C) testis (D) epididymis. Note the different sizes and the fact of existence or nonexistence of spermatozoa between 5 month and 11 month. Magnification: 400×.
Fig. 2. (A) Reverse transcription polymerase chain reaction (RT-PCR) analysis for the identification of MMP-9 and PGP9.5 in bovine testes. (B) Relative mRNA expression of pgp9.5 and mmp-9 in 5 month and 11 month bovine total cells. Error bars represent SD between three technical replicates. *p<0.05, unpaired t-test.
석하였다. RT-PCR 결과 각각 5개월 과 11개월 령 소 정소 에서 pgp9.5와 mmp-9이 발현하는 것을 확인할 수 있었다 (Fig. 2A). 실시간 Real-time PCR 결과 pgp9.5의 발현은 11 개월 령 보다 5개월 령 정소에서 유의적으로 높게 발현됨 을 확인할 수 있었고(p<0.05), mmp-9 발현량도 5개월 령 정소에서 유의적으로 높게 발현되었다(p<0.05, Fig. 2B).
3. 소 정소 내 MMP-9 특이 발현 세포 주 구명
성 성숙 전·후 소 정소에서 MMP-9가 발현되는 것을 확 인한 후 MMP-9가 발현하는 정소 내 세포 주를 확인하기 위하여 MMP-9 항체를 이용해 면역조직화학염색을 실시하 였다. 5개월 령 정소 조직에서 MMP-9는 곡세정관 내 기저 막에 부착된 세포에서 발현되는 것을 확인할 수 있었고 (Fig. 3A), 11개월 령 정소에서는 곡세정관 내 기저막 부위 의 세포와 기저막과 연접한 안쪽 세포에서 MMP-9의 발현 이 확인되었다(Fig. 3B). 더욱 정확한 세포의 특성을 구명 하기 위하여 소 정원세포의 표지인자로 알려진 PGP9.5 항 체와 공동 염색을 실시하였다. 5개월 령 정소의 곡세정관 에서 PGP9.5의 발현은 세정관 내 기저막 부착 세포에서 강하게 발현됨이 관찰되었고, PGP9.5가 발현되는 세포는
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:54 PM
www.earticle.net
Fig. 3. Immunofluorescence of bovine testis at 5 month (A and C) and 11 month (B and D). Green fluorescence represent MMP-9 positive cells (A and B). Red fluorescence indicates PGP9.5 (A and B) and DAZL positive cells (C and D). DAPI staining (blue) reveals the nucleus. White arrows indicate the cells with co-localization of MMP9 with PGP9.5 or DAZL. Magnification:
A and B are 200x; C and D are 400×.
MMP-9 가 발현되는 세포와 정확하게 일치하였다(Fig.
3A). 11개월 령 정소의 곡세정관 내에서 PGP9.5의 발현을 확인한 결과 주로 세정관 기저막에 부착된 세포 특이적으 로 발현했다(Fig. 3B). 또한 11개월 령 정소의 세정관 내 기 저막 부위의 일부 세포에서 MMP-9의 발현이 관찰되었으 나, PGP9.5의 발현은 확인되지 않았다(Fig. 3B).
MMP-9 가 발현되는 세포가 소 정원세포 및 생식세포라 는 것을 검증하기 위하여 PGP9.5외 또 다른 소 정원세포 의 표지인자로 알려진 DAZL 단백질의 항체와 공동염색을 실시하였다. 5개월 령 정소와 11개월 령 정소 모두에서 MMP-9와 DAZL 단백질의 발현이 곡세정관 내 생식세포 에서 발현되는 것이 확인 되었고, MMP-9와 DAZL이 발현 되는 세포가 일치하는 것을 확인할 수 있었다(Fig. 3C, D).
Ⅳ. 고찰
본 연구에서 우리는 소 정소 곡세정관 내에서 MMP-9의 발현 양상을 확인하였다. MMP-9는 5개월 령과 11개월 령 정소 조직에서 공통적으로 기저막에 부착되어 있는 세포 에서 발현되는 것을 확인하였고, 11개월 령 정소에서 기저 막과 연접한 안쪽의 세포에서도 MMP-9가 발현하는 것을 알 수 있었다. 또한, 이미 소 정원세포의 표지인자라고 밝 혀진 PGP9.5와 DAZL를 갖고 MMP-9과 공동 염색한 결과 동시에 염색되는 세포를 확인할 수 있었는데, 이는 MMP-9 가 발현되는 세포는 곡세정관의 기저막에 존재하는 정원 세포라고 판단된다. PGP9.5와 DAZL은 소 뿐만 아니라 돼 지, 염소, 쥐에서도 정원세포의 표지인자로 밝혀져 있다
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:54 PM
www.earticle.net
(Luo et al., 2006; Heidari et al., 2012; Kon et al., 1999;
LEE et al., 2013; Niu Z, 2014; Kim et al., 2012). 쥐 정원세 포 표지인자인 Inhibitor of differentiation 4(Id4)는 생후발 달(postnatal development) 동안 지속적으로 감소하는 양 상을 보였는데(Oatley et al., 2011; Chan et al., 2014), 이는 성숙한 정소 내 세포에서의 정원세포 비율이 감소함을 시 사한다. 본 연구에서도 pgp9.5 과 mmp-9 발현양이 5개월 령 보다 11개월 령에서 감소하였다. 따라서, PGP9.5, DAZL과 동시에 발현하며 성 성숙 이후 발현량이 감소하 는 MMP-9는 소 정원세포의 또 다른 표지인자로 사용될 수 있다고 판단된다.
선행 연구 결과들을 통해 다른 포유류에서 MMP-9의 발 현 양상도 알 수 있었다. 쥐에서는 발달단계별 특이적인 (stage-specific) 정소의 면역화학염색을 통해 MMP-9가 기 저막 혹은 관강 격실 부분(adluminal compartment)의 정 모세포와 정세포에서 발현된다는 보고가 있었고, 정원세포 에서도 발현할 것이라는 추측되어졌다(Siu et al., 2003;
Lydka et al., 2012). 개에서도 면역화학염색을 통해 확인한 결과 MMP-9가 정모세포와 원형정자세포에서 유의적으로 발현하는 것을 알 수 있었고, 유의적이진 않으나 정원세포 에서 발현하는 것을 알 수 있었다(Warinrak et al., 2015).
돼지에서는 체외 배양된 정원세포 전체 RNA의 차세대 염 기서열분석 결과 MMP-9가 성 성숙 전과 후 모두에서 정 원세포에서 발현하는 것을 확인할 수 있었다(Lee et al., 2016). 본 논문에 게시하지 않았지만, MMP-9의 종별 유전 자의 유사도를 계통도를 통해 확인한 결과, MMP-9의 유전 자 유사도는 소가 다른 종에 비해 돼지와 가까웠다. 이러 한 결과를 종합해 보면, MMP-9가 소에서 정원세포의 표지 인자로 사용될 수 있다고 판단된다.
Payne 등(2013)은 생식 모세포(gonocyte)가 정원세포로 분화하면서 정소 내강(lumen)에서 기저막 부위로 이동한 다고 보고하였다. 생식세포 이동을 공초점 레이저 현미경 을 통해 관찰한 결과, sertoli 세포의 오래된 치밀 결합이 없어지고 새로운 치밀 결합이 생기는 것을 확인하였다 (Smith et al., 2012). 또한, 생식 세포가 이동할 때 혈액세 정관관문(blood-testis barrier)이 재구성 되면서 sertoli 세 포 치밀 결합과 기저 외부 원형질 특수화(ectoplasmic specialization)가 풀어지는 세포외기질의 변화가 생기는 것을 알 수 있었다(Siu et al., 2008). MMP-9는 곡세정관 내 세포외기질 구성성분인 제 4형 콜라젠과 라미닌을 분해한 다고 밝혀졌다(Heikinheimo et al., 1995; Gu et al., 2005).
MMP-9의 세포외 기질 분해 능력은 종양괴사인자 알파
(tumor necrosis alpha, TNF-α)와 함께 세포외기질 재결합 에 영향을 미치며(Robinson et al., 2001), sertoli 세포의 치 밀결합 구조와도 연관되어 있다고 보고되었다(Siu et al., 2003). 또한 MMP-9가 결핍된 생쥐는 번식 능력이 감소하 는 문제점이 있다(Dubois et al., 2000). 본 실험에서는 성 성숙 전 곡세정관에서 정원세포가 기저막에 위치하고 MMP-9가 발현한다는 것을 확인하였다. 따라서 생식 모세 포가 성 성숙 이전에 MMP-9가 발현함으로써 sertoli 세포 치밀 결합과 세포외기질에 변화를 만들며 기저막으로 이 동할 수도 있는 가능성을 제시하며, 정원세포에서 분비된 MMP-9가 곡세정관 내 세포외기질의 재구성에도 연관이 있음을 시사하고 있다.
본 연구에서는 MMP-9의 생식세포 내 발현 위치와 다른 표지인자와의 공동 염색을 통해 소에서 정원세포의 표지 인자로 사용될 수 있음을 확인하였다. 또한, 성 성숙 이전 MMP-9의 발현 위치를 통해 MMP-9가 생식세포 발달에 영향을 줄 수 있을 것으로 예측할 수 있다. 하지만 아직 MMP-9을 통한 정원세포 이동에 대한 기전이 밝혀지지 않 았으므로 이와 관련된 추가연구가 필요하다고 판단된다.
Ⅴ. 요약
표지인자 발견은 세포를 선택적으로 분리하고 세포의 특성과 기능을 이해하는 데 용이하다. 기존에 밝혀진 소 정원세포의 표지인자로는 THY1, DBA, PGP9.5, DAZL 등 이 있고, 소 정원세포의 특성이나 정자형성과정을 이해하 기 위해서는 다양한 표지인자 연구가 필요하다. MMP-9는 세포외기질 물질과 결합조직의 기저막 물질을 분해하고, 특히 정소 내에서는 sertoli 세포의 치밀결합 구조 형성과 세포외기질 재형성에 관여한다. 또한 MMP-9는 쥐에서 정 모세포와 정세포, 개에서 정모세포와 원형정자세포, 그리 고 돼지에서 정모세포와 체외 배양된 정원줄기세포에서 발현한다. 따라서 본 연구에서는 소 정소 내 MMP-9의 발 현양상과 발현세포를 구명하였다. 우선 5개월 령과 11개월 령 소 정소를 갖고 Hematoxylin & Eosin 염색한 결과 5개 월 령은 성 성숙 이전, 11개월 령은 성 성숙 이후라는 것을 알 수 있었다. 성 성숙 이전과 성 성숙 이후 정소를 MMP-9와 PGP9.5, DAZL로 공동 면역염색을 통해서는 MMP-9는 기저막에 부착된 세포에서 발현하고, 이미 밝혀 진 표지인자와 동시에 염색되었다. 또한 mmp-9의 발현량 변화 양상은 다른 표지인자의 발현량 변화 양상과 유사했
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:54 PM
www.earticle.net
다. 이러한 결과를 종합해 보면, MMP-9이 소에서 정원세 포 표지인자로 사용될 수 있다고 판단한다. 추후 MMP-9이 정원세포 이동 기전에 미치는 영향을 밝히기 위한 추가 연 구가 필요하다.
사사
본 논문은 농촌진흥청 공동연구사업(과제번호: PJ010924) 의 지원에 의해 이루어진 것임
Ⅵ. REFERENCES
1. Allais-Bonnet, A. 2014. Role of the prion protein family in the gonads. Front Cell Dev. Biol. 2:56.
2. Chan, F. 2014. Functional and molecular features of the Id4+ germline stem cell population in mouse testes. Genes. Dev. 28(12):1351-62.
3. Dubois, B. 2000. Gelatinase B deficiency impairs reproduction. J. Clin. Invest. 106(5):627-8.
4. Gu, Z. 2005. A highly specific inhibitor of matrix metalloproteinase-9 rescues laminin from proteolysis and neurons from apoptosis in transient focal cerebral ischemia. J. Neurosci. 25(27):6401-8.
5. Heidari B. 2012. Isolation, identification, and culture of goat spermatogonial stem cells using c-kit and PGP9.5 markers. J. Assist. Reprod. Genet.
29(10):1029-38.
6. Heikinheimo, K. 1995. Expression of basement membrane type IV collagen and type IV collagenases (MMP-2 and MMP-9) in human fetal teeth. J. Dent.
Res. 74(5):1226-34.
7. Izadyar, F. 2002. Isolation and purification of type A spermatogonia from the bovine testis. Reproduction.
124(1):85-94.
8. Kim, B. 2012. DAZL is essential for stress granule formation implicated in germ cell survival upon heat stress. Development. 139(3):568-78.
9. Kon, Y. 1999. Expression of protein gene product 9.5, a neuronal ubiquitin C-terminal hydrolase, and its developing change in sertoli cells of mouse testis.
Mol. Reprod. Dev. 54(4):333-41.
10. LEE, W. Y. 2013. Establishment and in vitro culture of porcine spermatogonial germ cells in low temperature culture conditions. Stem Cell Res.
11(3):1234-49.
11. LEE, W. Y. 2016. Identification of putative biomarkers for the early stage of porcine spermatogonial stem cells using next-generation sequencing. PLoS One. 11(1):e0147298.
12. Luo, J. 2006. Protein gene product 9.5 is a spermatogonia-specific marker in the pig testis:
application to enrichment and culture of porcine spermatogonia. Mol. Reprod. Dev. 73(12):1531-40.
13. Lydka, M. 2012. Tumor necrosis factor α-mediated restructuring of the Sertoli cell barrier in vitro involves matrix metalloprotease 9 (MMP9), membrane- bound intercellular adhesion molecule-1 (ICAM-1) and the actin cytoskeleton. Spermatogenesis. 2(4):294- 303.
14. Moreno-Ruiz, P. 2016. Testis peritubular myoid cells increase their motility and express matrix- metalloproteinase 9 (MMP-9) after interaction with embryonal carcinoma cells. Andrology. 4(1):111-20.
15. Niu, Z. 2014. Conservation and function of Dazl in promoting the meiosis of goat male germline stem cells. Mol. Biol. Rep. 41(5):2697-707.
16. Oatley, M. J. 2011, Inhibitor of DNA binding 5 is expressed selectively by single spermatogonia in the male germline and regulates the self-renewal of spermatogonial stem cells in mice. Biol. Reprod.
84:347-356.
17. Payne, C. J. 2013. Cycling to and from a stem cell niche: the temporal and spatial odyssey of mitotic male germ cells. Int. J. Dev. Biol. 57(2-4):169-77.
18. Reding, S. C. 2010. THY1 is a conserved marker of undifferentiated spermatogonia in the pre-pubertal bull testis. Reproduction. 139(5):893-903.
19. Robinson, L. L. 2001. Matrix metalloproteinases and tissue inhibitors of metalloproteinases in human fetal testis and ovary. Mol. Hum. Reprod. 7(7):641-8.
20. Shah, S. M. 2015. Bone morphogenetic protein 4 (BMP4) induces buffalo (Bubalus bubalis) embryonic
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:54 PM
www.earticle.net
stem cell differentiation into germ cells. Biochimie.
119:113-24.
21. Siu, M. K. 2003. The interplay of collagen IV, tumor necrosis factor-alpha, gelatinase B (matrix metalloprotease-9), and tissue inhibitor of metalloproteases-1 in the basal lamina regulates Sertoli cell-tight junction dynamics in the rat testis.
Endocrinology. 144(1):371-87.
22. Siu, M. K. 2008. Extracellular matrix and its role in spermatogenesis. Adv. Exp. Med. Biol. 636:74-91 23. Smith, B. E. 2012. Germ cell migration across sertoli
cell tight junctions. Science. 338(6108):798–802.
24. Stockwell, S. 2009. Microsatellite detection of donor-derived sperm DNA following germ cell transplantation in cattle. Reprod. Fertil. Dev.
21(3):462-8.
25. Warirak, C. 2015. Expression of matrix metalloproteinases (MMP-2, MMP-9) and their inhibitors (TIMP-1, TIMP-2) in canine testis, epididymis and semen.
Reprod. Domest. Anim. 50(1):48-57.
26. Woessner, J. F. 1991. Matrix metalloproteinases and their inhibitors in connective tissue remodeling.
FASEB J. 5(8):2145-54.
27. Wrobel, K. H. 1995. Configuration and distribution of bovine spermatogonia. Cell. Tissue. Res. 279(2):
277-89.
(Received 01 September 2016, Revised 24 September 2016, Accepted 26 September 2016)
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 7:54 PM