137
Copyright © 2016 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
대부분의척추동물및무척추동물들은세균
,
바이러스,
곰팡 이,
또는기생충등과같은외부미생물의침입으로부터자신 을방어하기위한효과적인면역체계를각각포함하고 있다(Hancock and Diamond, 2000).
선천면역과적응면역이각각 잘발달되어있는척추동물과는달리무척추동물은선천면역 만을포함하고있으므로외부환경에대한방어작용은오로지 선천면역에만의존하고있다.
일반적으로선천면역은외부침 입물질에대한초기방어및적응면역의유도를담당하므로그 기능과역할에대한관심이최근들어서증대되고있는실정이 다.
선천면역을담당하는대표적인물질로는lectin, propheno- loxidase activating factor, phagocytosis, antimicrobial peptide
등이알려져있으며,
이들은외부로부터생체내로의이물질침입에대한
1
차방어역할을담당한다고알려져있다(Relf et al., 1999; Tincu and Taylor, 2004).
최근들어서여러가지의선천 면역요소들중에서비특이적이고빠른반응성을가지는자연 친화적인항균펩타이드에대한관심이증가되고있는실정이 다(Zasloff, 2002).
항균펩타이드는유전자에의해암호화되어있는양친매성의
α-helix
나β-sheet
구조를취하는positive net charge
를가지는12-100
개의아미노산으로구성된선천면역에관여하는물질로서
,
미생물뿐만아니라무척추동물에서척추동물에이르는넓 은범위에서분포하고있는단백질성항균물질이다(van’'t Hof et al., 2001; Stark et al., 2002; Izadpanah and Gallo, 2005).
또 한항균펩타이드는막을주요target site
로삼고있으므로내성 균유발의가능성이거의없는특징을가지고있어서차세대항 생제개발을위한주요후보물질로서각광을받고있다(Han-
바지락( Ruditapes philippinarum) 아가미로부터 항균 펩타이드의 탐색 및 정제
서정길*
국립 군산대학교 해양과학대학 식품생명공학과
Screening and Purification of an Antimicrobial Peptide from the Gill of the Manila Clam Ruditapes philippinarum
Jung-Kil Seo*
Department of Food Science and Biotechnology, Kunsan National University, Kunsan 54150, Korea
This study screened the biological activity of an acidified gill extract of the Manila clam Ruditapes philippinarum in- cluding antimicrobial, hemolytic, membrane permeabilization, and DNA-binding activity, and purified the antimicro- bial material. The acidified gill extract showed potent antimicrobial activity against Bacillus subtilis and Escherichia coli without significant hemolytic activity, but showed no membrane permeabilization or DNA-binding ability. An antimicrobial material was purified from the acidified gill extract using C 18 reversed-phase and cation-exchange high- performance liquid chromatography (HPLC). Treatment of the purified material with trypsin completely abolished all of the antibacterial activity against Bacillus subtilis , suggesting that the purified material is a proteinaceous antibiotic.
The molecular weight of the purified material was 2571.9 Da, but no primary structural information was obtained due to N-terminal blocking. A future study should confirm the primary structure. Our results suggest that the Manila clam gill contains proteinaceous antibiotics that have a role in first-line defense. This information could be used to better understand the Manila clam innate immune system.
Key words: Manila clam, Ruditapes philippinarum , Antimicrobial peptide, Innate defense, Purification
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2016.0137 Korean J Fish Aquat Sci 49(2) 137-145, April 2016
Received 29 February 2016; Revised 1 April 2016; Accepted 7 April 2016
*Corresponding author: Tel: +82. 63. 469. 1827 Fax: +82. 63. 469. 7448
E-mail address: [email protected]
138
서정길cock REW and Diamond G, 2000; Zasloff M, 2002).
항균펩타이드는구조에따라서크게
4
가지종류로나눌수 있다(Andreu and Rivas, 1998): i) Cys
을포함하지않는linear α-helical peptide, ii) Cys
을포함하는looped peptide, iii)
특 이구조를취하지않는random extended peptide, iv)
특이아 미노산을많이포함한peptide
등이다.
항균펩타이드는이온 결합과소수성상호작용을이용한barrel-stave model, toroidal model
또는carpet model
을통해서membrane pore
를형성하 거나세포내부로들어가세포내기관이나대사과정의저해를 통해서항균효과를나타낸다고알려져있다(Shai, 2002; Li, 2009).
현재까지해양무척추동물유래항균peptide
는갯지렁 이(Ovchinnikova et al., 2004),
군소(Iijima et al., 2003),
투구 게(Miyata et al., 1989; Saito et al., 1995),
꽃게(Khoo et al., 1999),
흰다리새우(Destoumieux et al., 1997),
홍합(Charlet et al., 1996; Mitta et al., 1999; Mitta et al., 2000),
굴(Seo et al., 2005; 2012; 2013; Nam et al., 2015; Gueguen et al., 2006),
전 복(De Zoysa et al., 2009)
등이보고되어있다.
바지락은우리나라의패류산업에서큰비중을차지하는패류 양식자원중의하나로서중요성이크게증대되고있으나
,
최근 들어서다양한병원성미생물감염원에의한대량폐사가일어 나고있어서경제적,
환경적으로큰문제가되고있다(Park et al., 2006).
따라서이러한문제점을해결하기위한기초자료로 서바지락의host defense mechanism
을이해하고,
질병감염원 의종류또는분포와바지락의선천면역요소들간의상호연관 성연구가절실히요구되고있는실정이다.
특히,
다양한해양무 척추동물에있어서선천면역요소중의하나인항균펩타이드의 질병감염에대한효과적인방어기능이보고되면서질병감염에 대한연구가활발히진행되고있는반면,
바지락과연관된항균 펩타이드에대한연구는극히제한적으로진행되고있는실정 이다.
현재까지보고된바지락유래항균펩타이드는defensin (Zhang et al., 2015), big defensin (Zhao et al., 2010)
또는mus- sel defensin MGD-1
유사peptide (Kang et al., 2006)
등과같 은defensin
류에한정되어있으므로선천면역을담당하는다양 한항균펩타이드에대한연구가필요한실정이다.
따라서본연구는바지락
(Manila clam, Ruditapes philippina-
rum)
의면역관련조직인아가미추출물을이용해서항균활성의탐색및항균물질의정제과정을수행하고항균물질의구조 규명과정을수행함으로써바지락의선천면역에대한기초자료 를제공하고자수행되었다
.
재료 및 방법
실험동물
본실험에사용한바지락
(Manila clam, Ruditapes philippina- rum)
은충청남도곰소만에서구입하였으며바지락의아가미와 외투막조직을추출에사용하였다.
모든추출물은사용하기전까지
-70
oC
에보관하였다. 시약 및 재료
항균활성측정을위한
tryptic soy broth (TSB)
와agarose type I (Low EEO Agar)
은Merck
사(Merck, Darmstadt, Germany)
와Sigma
사(St. Louis, MO, USA)
에서각각 구입하여 사용 하였다.
정제과정에서 사용된HPLC
용water
와acetonitrile (CH
3CN)
은Tedia
사(Ohio, USA)
로부터구입하였고,
그이외 의모든시약들은특급을사용하였다.
추출방법
바지락의패각을흐르는수돗물에씻어서패각오염물을제거 한후
,
아가미와외투막조직을취하여5 mL volume
이될때까 지얼음속에서모았다.
모아진아가미와외투막조직에끓고있 는4
배량의1% acetic acid(v/v)
를첨가하여100℃
에서5
분동 안끓인후,
얼음에보관하여충분히냉각시켰다.
충분히냉각 된조직은homogenizer (T10 basic ULTRA-TURRAX, IKA, USA)
를사용하여완전히파쇄시켰다.
조직파쇄액은4℃
에서20
분동안8,000 g
로원심분리(VS-21SMT, Vision Scientific, Buchon, Korea)
를행한후에상층액을취해서항균활성탐색 및항균물질의정제과정에사용하였다.
항균활성 측정방법 및 사용균주
항균활성 측정에는 그람 양성균인
Bacillus subtilis KCTC 1021
과그람음성균인Escherichia coli D31 (
미국North Caro- lina
주립대수의과대학Noga
교수가제공)
을사용하였다.
활 성측정방법으로는서로다른농도를포함한두층의배지를 사용하는ultrasensitive radial diffusion assay (URDA)
법을이 용하였다(Lehrer at al., 1991).
항균활성측정에사용된균주는 우선tryptic soy broth (TSB)
에접종한후18
시간동안37℃
에서
pre-culture
를 수행한후colorimeter (Product No. 52- 1210, BioMerieux, Inc., USA)
를사용하여균농도를84%T (≒ 1×10
8CFU/mL)
가되도록조정하였다.
그후, 9.5 mL
의0.03% TSB, 1% TypeⅠ agarose
및10 mM phosphate buffer (PB) (pH 6.5)
를포함하는underlay gel
에각각의농도로희석 된균액0.5 mL
를넣고잘섞은후에plate
에편평하게부어굳 혔다.
굳은plate
에punch
를사용하여직경2.5 mm
의well
을뚫은후에
5 μL
의각추출물또는정제단계의분획물을도입시켰다
.
추출물또는분획물이배지에스며들면3
시간동안37℃
에 서1
차배양한후,
그위에10 mL
의6% TSB, 1% TypeⅠ aga-
rose
및10 mM phosphate buffer (pH 6.5)
를포함하는overlay
gel
을붓고굳힌후에동일한온도에서18
시간동안2
차배양하 였다.
다음날well
주위에생긴clear zone
의크기를측정하여 항균활성을확인하였다.
항균활성측정동안positive control
로 는미국산잡종농어(Morone saxatilis×Morone chrysops)
의mast cell
에서정제된항균펩타이드인piscidin 1
을사용하였고negative control
로는1% acetic acid
또는0.01% acetic acid
를사용하였다
(Silphaduang and Noga, 2001).
Protease 처리에 의한 단백질성 확인
추출물또는정제된항균물질의단백질성확인을위해서
tryp- sin
처리전·
후의항균활성변화를확인하였다.
바지락추출물 또는정제된물질4 μL
에trypsin (1,000 μg/mL) 1 μL
를첨가 하고37℃
에서60
분간반응시킨뒤, URDA
법으로B. subtilis
KCTC1021
에대한항균활성변화를측정하였다.
Hemolysis
추출물에존재하는독성물질의유무를확인하기위해서인간 의적혈구에대한용혈활성을측정하였다
(Seo et al., 2014).
사 람의혈액(Blood type: B)
에동량의phosphate buffered saline (PBS, 50 mM sodium phosphate, 150 mM NaCl, pH 7.4)
을넣 고혼합한후800 rpm
에서15
분간원심분리하여상등액은버 리고침전물을얻었다.
이러한세척과정을3
회반복한후얻어 진적혈구를1%
혼합용액이되도록PBS
를첨가한후사용하 였다.
얻어진적혈구90 μL
를96-well microplate
에도입시키 고바지락추출물또는정제물질10 μL
를첨가한다음37℃
에 서60
분간반응시켰다.
이후반응액을원심분리(1,500 rpm, 10
분, 4℃)
하여얻어진상등액80 μL
를새로운microtiter plate
로 옮긴후헤모글로빈유출정도를확인하기위해서490 nm
에서 흡광도를측정하였다.
적혈구의100%
용혈을위한비교물질로 는triton X-100 (0.1%)
을사용하였다.
Acid-Urea PAGE와 bug-blot
바지락추출물내의양이온성단백질
/
펩타이드의조성분포와 활성band
를확인하기위해서acid-urea PAGE
와bug-blot
을E. coli D31
에대해서수행하였다(Seo et al., 2005). Bug-blot
에 사용된E. coli D31
은TSB
에서37℃
로18
시간동안배양한후colorimeter (Product No. 52-1210, BioMerieux, Inc., USA)
를사 용하여균농도를84%T (≒ 1×10
8CFU/mL)
가되게조정하 였다.
그후, 9.5 mL
의LB, 1% TypeⅠ agarose, 10 mM phos- phate buffer (PB) (pH 6.7)
및NaCl
를포함하는gel
에희석된균 액0.5 mL
를넣고잘섞은후에plate
에편평하게부어굳혔다.
AU-PAGE
를위한gel
은urea 3.6 g, D.W 3.4 mL, 30% acryl- amide/1.6% bis-acrylamide solution 5 mL, glacial HAc 0.65 mL
를비커에넣고stirring (10min, RT)
과degassing (20 min, RT, desiccator)
을한후TEMED 0.06 mL, 10% APS 0.275 mL
를첨가한후stirring (1 min, RT)
하고, gel
액을붓고유출 유무를확인한뒤comb
를꽂은후충분히실온에서굳혔다.
이 후장치에5% HAc
를붓고pre-run (150 v, 60 min, RT)
을하 였다. Well
에도입할각시료는20 µL
의2×sample buffer
와1:1(v/v)
로혼합해서사용하였고, standard
는histone H1 (~21 kDa), lysozyme (~11 kDa), aprotinin (~6.5 kDa)
및piscidin 1 (~2.5 kDa)
을포함하는20 µL
의0.01% HAc
와2×Sample buffer
를1:1 (v/v)
로혼합해서사용하였다. Pre-run
이끝난후running buffer (5% HAC)
를교체하고well
에시료를도입시 킨후전기영동을실시하였다(150 v, 45 min).
전기영동후, 1
개의gel
은CBB R-250
으로염색시켰고,
나머지1
개의gel
을rinse buffer (10 mM PB buffer, pH 7.4)
로2
회세척을한후에E. coli D31
을포함하는LB plate
에overlay
시킨후37℃
에서3
시간배양하였다.
배양후에overlay
시켰던gel
을제거한후에LB plate
는동일한온도에서18
시간동안2
차배양하였다.
다음 날LB plate
에나타난clear zone
을염색및탈색된gel
의band
결과와비교하였다.
추출물들의 cytoplasmic membrane permeabili- zatin assay
바지락추출물의세균내막투과성측정을위해서
nonmem-
brane-permeative chromogenic substrate
인o-nitrophenyl-β- D-galactopyranoside (ONPG)
에대한추출물도입후의E. coli ML35p
의세포질에서유출된β-galactosidase
의활성을확인하 는실험을수행하였다(Skerlavaj et al., 1990).
배양된mid-log phase
의E. coli ML35p
를10 mM sodium phosphate buffer (pH 7.4)
로세척을한후, 1.5 mM
의ONPG
를포함하는동일butter
에용해시켰다.
그후,
측정할추출물들을첨가한뒤37℃
에서
60
분간배양하면서10
분간격으로유출된β-galactosidase
에의한ONPG
의o-nitrophenol
로의가수분해정도를405 nm
에서측정하였다.
막투과활성측정의표준물질로는강한막투 과성을포함하는것으로알려진항균펩타이드인piscidin 1
을 사용하였다.
DNA Binding Assay
바지락추출물에존재하는염기성의
DNA
결합성물질의존재유무를확인하기위해서
DNA binding
에의한DNA band
들 의agarose gel-electrophoresis
에서의이동의저해정도를확인 하는DNA-binding assay
를수행하였다(Hsu et al., 2003; Al- fred et al., 2013).
이를위해서1 kb DNA marker (λ-HindIII- digested DNA, 50 ng)
와추출물을혼합해서37℃
에서60
분동 안반응시킨후0.5 μg/mL EtBr
을포함하는1.4% agarose gel
에서전기영동을수행한후DNA-binding
정도를확인하였다. 항균활성물질의 분리 및 정제
아가미추출물에존재하는 항균물질의분리 및정제과정은
HPLC (YL9100 HPLC system, Young Lin Instrument Co.,
Korea)
를이용하여수행하였다.
추출물에존재하는항균물질의정제과정은역상및이온교환성을갖는
2
가지특성의HPLC
column
을사용하여수행되었고,
정제과정중의분획들에대한항균활성은
B. subtilis KCTC1021
에대해서URDA
법으로측 정하였다.
첫번째정제과정으로CapCell-Pak C
18column (4.6
mm×250 mm, Shiseido, Japan)
을이용하여다음과같은조건 으로분리과정을수행하였다. A
용매는0.1% Trifluoroacetic
acid (TFA)
를포함하는H
2O (pH 2.2)
이며B
용매는0.1% TFA
140
서정길를포함하는
100% CH
3CN (pH 2.2)
를사용하였다.
분리조건 은5%
에서65%
까지60
분간B
용매를순차적으로증가시켰으 며,
유속은1.0 mL/min,
파장은220 nm
에서확인하였고모든 정제과정은실온에서수행되었다.
항균활성을지닌분획들을 모아서TSK-gel SP-5PW column (7.5 mm×75 mm, Tosoh,
Japan)
에도입시키고다음과같은조건으로분리과정을수행하였다
. A buffer
는10 mM phosphate buffer (PB, pH 6.0)
이 며B buffer
는1.0 M NaCl
을포함하는10 mM PB (pH 6.0)
를사용하였다.
분리조건은0%
에서100%
까지100
분동안B buffer
를순차적으로증가시켰으며,
유속은1.0 mL/min,
파장 은220 nm
에서분리과정을수행하였다.
활성이확인된분획은CapCell-Pak C
18column (4.6 mm×250 mm)
에적용시켜다 음과같은조건으로최종분리과정을수행하여단일peak
를정 제하였다. A
용매는0.1 % TFA
를포함하는H
2O (pH 2.2)
이며B
용매는0.1% TFA
를포함하는100% CH
3CN (pH 2.2)
를사 용하였다.
분리조건은5%
에서45%
까지40
분간B
용매를순차 적으로증가시켰으며,
유속은1.0 mL/min,
파장은220 nm
에 서확인하였다.
분자량 측정
최종정제된물질의분자량은
Ultraflex III matrix-assisted la- ser desorption ionisation time-of-flight (MALDI-TOF) mass spectrometer (Bruker Daltonik GmbH, Bremen, Germany)
를 사용하여linear mode
로 측정하였다.
정제물질을0.1%
TFA/50% CH
3CN (1:1, v/v)
에녹인후α-cyano-4- hydroxy- cinnamic acid (CHCA) matrix solution (10 mg/mL CHCA in 0.1% TFA/50% CH
3CN, 1:1, v/v)
과혼합하여MALDI plate
에도입한후에분자량을측정하였다. Standard
로는Calmix 2
를사용하였다.
펩타이드 일차구조 분석
정제물의
N-terminal amino acid sequence
는pulse liquid au- tomatic sequencer (model 473A; Applied Biosystems, Foster City, CA, USA)
를사용하여분석하였다.
결과 및 고찰
바지락 추출물의 생리활성
바지락 추출물의 항균활성
바지락
(Manila clam, Ruditapes philippinarum)
으로부터채취한아가미와외투막조직들은
1% HAC
를이용한약산추출과정을수행하였으며
,
각추출물내의단백질성항균물질의존 재유무를확인하기위하여각추출물(4 μL)
의trypsin
처리전∙
후의항균활성변화를URDA
법을이용하여B. subtilis
및E.
coli D31
에대해서측정하였다(Fig. 1).
실험결과,
각추출물은B. subtilis
와E. coli D31
에대해서강한항균활성을나타내었으며
, trypsin
처리후에는항균활성이현저히감소되었다.
이러 한결과는바지락추출물에단백질성항균물질이포함되어있 음을의미하는것이다.
또한추출물또는균주간의활성비교 에서는아가미추출물이외투막추출물보다는강한항균활성 을나타내었으며, E. coli D31
보다B. subtilis
에더욱효과적인 것으로나타났다.
이러한결과를토대로바지락유래단백질성 항균물질을정제하기위해서아가미추출물을연구과정에사 용하였다.
Acid-urea PAGE와 bug-blot
바지락추출물내의염기성단백질
/
펩타이드의구성분포와그 들의항균활성유무를확인하기위해서AU-PAGE
와bug-blot
를수행하였다(Fig. 2).
그결과,
바지락추출물들은저분자물 질보다는상대적으로고분자물질의농도가높게나타났으며,
외투막추출물보다는아가미추출물에서구성물질들의다양성 이더높게나타났다.
또한,
추출물의항균활성은slow migra- tion zone (SMZ) (~11 kDa)
과rapid migration zone (RMZ) (~2.5 kDa)
을포함하는두위치에서나타났으며, RMZ
의항 균활성이SMZ
보다는강하게나타났다.
이러한결과는바지락 추출물에는다양한분자량의항균물질이존재하고있으며,
아 마도저분자의항균물질이보다중추적인역할을담당하는것 으로판단된다.
바지락 추출물의 hemolysis
바지락추출물들에포함된독성물질의유무를확인하기위해 서추출물을인간적혈구
(
혈액형: B
형)
에대한용혈활성(hemo- lytic activity)
을측정하였다(Fig. 3).
비교물질로는piscidin 1
을 사용하였으며100% hemolysis
의표준물질로는0.1% Triton-
X 100
을사용하였다.
측정결과대조군으로사용된항균펩타이드인
piscidin 1
은낮은농도(12.5 μg/mL)
에서도강한용혈활 성을나타낸반면,
바지락추출물들은용혈활성을거의나타내 지않았다.
이러한결과는바지락추출물들은세포독성을나타 내는물질을거의포함하고있지않다는것을의미하는것이다.
Fig. 1. Antimicrobial activity of the crude extracts of the Manila clam Ruditapes philippinarum. Antimicrobial activity of gill or mantle extract (before) and trypsin treated extract (after) against B. subtilis KCTC1021 and E. coli D31. Scale bar indicates 5 mm.B. subtilis
KCTC1021
E. coli D31
Before After Before After
Gill Mantle
: 5 mm
Histone H1 (~21 kDa)
Lysozyme (~11 kDa) Aprotinin (~6.5 kDa) Piscidin 1 (~2.5 kDa)
1 2 3
(A)
1 2 3
Histone H1 (~21 kDa)
Lysozyme (~11 kDa) Aprotinin (~6.5 kDa) Piscidin 1 (~2.5 kDa) (B)
1 2 3 4 5
1: λ-HindIII-digested DNA 2: Buffer without peptide 3: Gill
4: Mantle 5: Piscidin 1
(---) % CH
3CN---
5 35 65
Absorbance at 220 nm
0 0.1 0.2 0.3 0.4 0.5
1.2
1
0.8
0.6
0.4
0.2
0 0
Piscidin 1 0.01% HAc Gill Mantle
10 20 30
Time (min)
Fluorescence Intensity
40 50 60
0 10 20 30 40 50 60
Time (min)
: 5 mm
(A)
Absorbance at 220 nm
0 0 0.15 0.30
Time (min)
20 40 60 80 100
0
0.5 1.0
(---) NaCl (M)
: 5 mm
(B)
(---) % CH
3CN
5 25 45
Absorbance at 220 nm
0 0.04 0.08 0.12 0.15
0 10 20 30 40
Time (min)
: 5 mm
(A)
Mass [m/z]
% Intensity
20 40 60 80
799.0 1441.8 2084.6 2727.4 3370.2 4013.0
100 1.0E+4
[M+2H]2+
1286.9
0
(B)
[M+H]+ 2571.9
100
90 80 70 60 50 40 30 20 10
0 0
Gill Mantle Piscidin 1
10 20 30
Peptide concentration (μg/mL)
Hemolysis (%)
40 50 60 70 80 90 100
바지락 아가미 항균 펩타이드
141
바지락 추출물에 포함된 항균물질의 작용부위 확인 실험
바지락추출물에포함된항균물질의target site
를예측하기위해서세균내막투과성과핵내구성물인
DNA
와의상호작용 유무를관찰하였다.
우선,
항균물질의target site
가핵내부의DNA
인지를확인하기 위해서DNA-peptide interaction
에의 한DNA migration
저해현상을관찰할수있는DNA-binding assay
를수행하였다(Fig. 4).
그결과,
양성대조군으로사용한piscidin 1
은강한DNA-binding
활성을나타낸반면바지락추 출물들은DNA migration
에전혀영향을주지않는것으로나 타났다.
이러한결과는바지락추출물에존재하는항균물질은 핵내의DNA
와의상호작용에의해항균작용을나타내는것은 아님을의미하는것이다.
두번째로,
추출물에포함된항균물질 의target site
가세포막자체인지를확인하기위해서membrane permeabilization assay
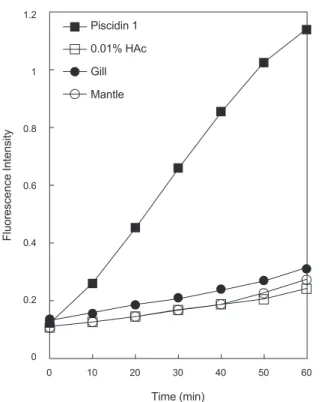
를 수행하였다(Fig. 5).
실험결과양성 대조군으로사용한piscidin 1
은강한내막투과성을나타낸반 면음성대조군으로사용한1% HAc
와측정에사용된바지락 추출물들은막투과활성을거의나타내지않았다.
이것은실험에사용된
1% HAc
는막투과활성에거의영향이없음을나타내는것이므로바지락추출물들에는막투과활성물질이거의 포함하고있지않다는것을의미하는것이다
.
이러한결과는추 출물내의항균물질의예상작용target site
은막자체는아니라 는것을의미하는것이다.
두가지의실험결과바지락추출물에 포함된항균물질의작용기작은세포막을직접공격하거나핵 내부의DNA
와의직접적인상호작용에의한것보다는세포표 면물질들과의상호작용또는세포질에존재하는세포내구성 성분에작용할가능성이있다는것을의미하는것이다.
Fig. 2. AU-PAGE gel overlay assay of gill and mantle extractagainst E. coli D31. (A) AU-PAGE run and stained with Coomass- ie Brilliant Blue R-250. Lane 1, molecular weight markers: 3 μg of human histone H1, 2 μg of human lysozyme, 1 μg of aprotinin, and 1 μg of piscidin 1, lane 2: 20 μL of gill extract, lane 3: 20 μL of mantle extract. (B) AU-PAGE gel overlay assay of gill and mantle extract against E. coli D31.
B. subtilis
KCTC1021
E. coli D31
Before After Before After
Gill Mantle
: 5 mm
Histone H1 (~21 kDa)
Lysozyme (~11 kDa) Aprotinin (~6.5 kDa) Piscidin 1 (~2.5 kDa)
1 2 3
(A)
1 2 3
Histone H1 (~21 kDa)
Lysozyme (~11 kDa) Aprotinin (~6.5 kDa) Piscidin 1 (~2.5 kDa) (B)
1 2 3 4 5
1: λ-HindIII-digested DNA 2: Buffer without peptide 3: Gill
4: Mantle 5: Piscidin 1
(---) % CH
3CN---
5 35 65
Absorbance at 220 nm
0 0.1 0.2 0.3 0.4 0.5
1.2
1
0.8
0.6
0.4
0.2
0 0
Piscidin 1 0.01% HAc Gill Mantle
10 20 30
Time (min)
Fluorescence Intensity
40 50 60
0 10 20 30 40 50 60
Time (min)
: 5 mm
(A)
Absorbance at 220 nm
0 0 0.15 0.30
Time (min)
20 40 60 80 100
0
0.5 1.0
(---) NaCl (M)
: 5 mm
(B)
(---) % CH
3CN
5 25 45
Absorbance at 220 nm
0 0.04 0.08 0.12 0.15
0 10 20 30 40
Time (min)
: 5 mm
(A)
Mass [m/z]
% Intensity
20 40 60 80
799.0 1441.8 2084.6 2727.4 3370.2 4013.0
100 1.0E+4
[M+2H]2+
1286.9
0
(B)
[M+H]+ 2571.9
100
90 80 70 60 50 40 30 20 10
0 0
Gill Mantle Piscidin 1
10 20 30
Peptide concentration (μg/mL)
Hemolysis (%)
40 50 60 70 80 90 100
Fig. 3. Hemolytic activities of gill or mantle extract and piscidin 1 against human erythrocytes (blood type: B).
B. subtilis
KCTC1021
E. coli D31
Before After Before After
Gill Mantle
: 5 mm
Histone H1 (~21 kDa)
Lysozyme (~11 kDa) Aprotinin (~6.5 kDa) Piscidin 1 (~2.5 kDa)
1 2 3
(A)
1 2 3
Histone H1 (~21 kDa)
Lysozyme (~11 kDa) Aprotinin (~6.5 kDa) Piscidin 1 (~2.5 kDa) (B)
1 2 3 4 5
1: λ-HindIII-digested DNA 2: Buffer without peptide 3: Gill
4: Mantle 5: Piscidin 1
(---) % CH
3CN---
5 35 65
Absorbance at 220 nm
0 0.1 0.2 0.3 0.4 0.5
1.2
1
0.8
0.6
0.4
0.2
0 0
Piscidin 1 0.01% HAc Gill Mantle
10 20 30
Time (min)
Fluorescence Intensity
40 50 60
0 10 20 30 40 50 60
Time (min)
: 5 mm
(A)
Absorbance at 220 nm
0 0
0.15 0.30
Time (min)
20 40 60 80 100
0
0.5 1.0
(---) NaCl (M)
: 5 mm
(B)
(---) % CH
3CN
5 25 45
Absorbance at 220 nm
0 0.04 0.08 0.12 0.15
0 10 20 30 40
Time (min)
: 5 mm
(A)
Mass [m/z]
% Intensity
20 40 60 80
799.0 1441.8 2084.6 2727.4 3370.2 4013.0
100 1.0E+4
[M+2H]2+
1286.9
0
(B)
[M+H]+ 2571.9
100
90 80 70 60 50 40 30 20 10
0 0
Gill Mantle Piscidin 1
10 20 30
Peptide concentration (μg/mL)
Hemolysis (%)
40 50 60 70 80 90 100
Fig. 4. Gel retardation analysis or the binding of gill or mantle extract to DNA. Binding of extract to DNA was assessed by mea- suring the retardation of commercial molecular weight marker λ-HindIII-digested DNA (50 ng) migration through an agarose gel.
Lane 1: λ-HindIII-digested DNA(50 ng), lane 2: negative control, 0.0.1% HAc, lane 3: gill extract, lane 4: mantle extract, lane 5:
positive control, piscidin 1 1 μg.
B. subtilis
KCTC1021
E. coli D31
: 5 mm
Histone H1 (~21 kDa)
Lysozyme (~11 kDa) Aprotinin (~6.5 kDa) Piscidin 1 (~2.5 kDa)
1 2 3
(A)
1 2 3
Histone H1 (~21 kDa)
Lysozyme (~11 kDa) Aprotinin (~6.5 kDa) Piscidin 1 (~2.5 kDa) (B)
1 2 3 4 5
1: λ-HindIII-digested DNA 2: Buffer without peptide 3: Gill
4: Mantle 5: Piscidin 1
(---) % CH
3CN---
5 35 65
Absorbance at 220 nm
0 0.1 0.2 0.3 0.4 0.5
1.2
1
0.8
0.6
0.4
0.2
0 0
Piscidin 1 0.01% HAc Gill Mantle
10 20 30
Time (min)
Fluorescence Intensity
40 50 60
0 10 20 30 40 50 60
Time (min)
: 5 mm
(A)
Absorbance at 220 nm
0 0
0.15 0.30
Time (min)
20 40 60 80 100
0
0.5 1.0
(---) NaCl (M)
: 5 mm
(B)
(---) % CH
3CN
5 25 45
Absorbance at 220 nm
0 0.04 0.08 0.12 0.15
0 10 20 30 40
Time (min)
: 5 mm
(A)
Mass [m/z]
% Intensity
20 40 60 80
799.0 1441.8 2084.6 2727.4 3370.2 4013.0
100 1.0E+4
[M+2H]2+
1286.9
0
(B)
[M+H]+ 2571.9