Anti-inflammatory Activity of Antimicrobial Peptide Zophobacin 1 Derived from the Zophobas atratus
Yong Pyo Shin†, Joon Ha Lee†, In-Woo Kim, Minchul Seo, Mi-Ae Kim, Hwa Jeong Lee, Minhee Baek, Seong Hyun Kim and Jae Sam Hwang*
Department of Agricultural Biology, National Institute of Agricultural Sciences, Rural Development Administration, Wanju 55365, Korea Received July 21, 2020 /Revised September 7, 2020 /Accepted September 21, 2020
The giant mealworm beetle, Zophobas atratus (Coleoptera: Tenebrionidae) has been used as a protein source for small pets and mammals. Recently, it was temporarily registered in the list of the Food Code. We previously performed an in silico analysis of the Zophobas atratus transcriptome to identify putative antimicrobial peptides and identified several antimicrobial peptide candidates. Among them, we assessed the antimicrobial and anti-inflammatory activities of zophobacin 1 that was selected bio- informatically based on its physicochemical properties against microorganisms and mouse macrophage Raw264.7 cells. Zophobacin 1 showed antimicrobial activities against microorganisms without induc- ing hemolysis and decreased the nitric oxide production of the lipopolysaccharide-induced Raw264.7 cells. Moreover, ELISA and Western blot analysis revealed that zophobacin 1 reduced expression lev- els of pro-inflammatory enzymes such as inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2). We also investigated expression of pro-inflammatory cytokines (interleukin-6 and interleukin- 1β) production through quantitative real time-PCR and ELISA. Zophobacin 1 markedly reduced the expression level of cytokines through the regulation of mitogen-activated protein kinases (MAPKs) and nuclear factor kappa B (NF-κB) signaling. We confirmed that zophobacin 1 bound to bacterial cell membranes via a specific interaction with lipopolysaccharides. These data suggest that zophobacin 1 could be promising molecules for development as antimicrobial and anti-inflammatory therapeutic agents.
Key words : Anti-inflammatory activity, antimicrobial activity, antimicrobial peptide, RNA sequencing, Zophobas atratus
†Authors contributed equally.
*Corresponding author
*Tel : +82-63-238-2974, Fax : +82-63-238-3833
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
서 론
염증반응(inflammation)은 외부인자들의 침입에 대항하기 위한 방어기전 중 하나로 선천성 면역체제(innate immunity system)의 중요한 부분을 차지한다. 선천성 면역체제는 호스 트 세포(host cell)의 인식단백질인 pattern recognition recep- tors (PRRs)가 병원성 유기체들을 인식하여 활성화 되는 기능 적이고 물리적인 방어 기전이다[6]. 단핵구(monocytes)와 대 식세포(macrophages) 같은 호스트 세포들은 선천성 면역체제 에서 침입한 미생물들을 제거하기 위한 첫 면역 방어선으로 작용하며, 감염된 장소로 다수의 세포들이 이동하여 면역반응 에 관여하게 된다. 또한, 일부 전염증성 사이토카인(pro-in- flammatory cytokines)들도 대식세포로부터 매개되는 선천성
면역체제에서 중요한 분자로서 작용하게 된다[3].
Lipopolysaccharide (LPS)는 그람 음성균에 의한 패혈증과 패혈증 쇼크(inflammation sepsis and shock)의 병태생리학 (pathophysiology)에서 중요한 역할을 한다[1]. LPS는 그람음 성균의 세포벽 성분 중 하나로 세균이 분화하거나 사멸하는 과정에서 외부로 방출될 수 있으며, 외부로 방출된 LPS가 혈 류체제(blood system)로 들어가게 되면 혈액 내 단핵구와 대 식세포를 자극하여 tumor necrosis factor-α (TNF-α), inter- leukin-6 (IL-6), interleukin-8 (IL-8)과 같은 사이토카인들의 발 현을 유도한다. 이러한 사이토카인들이 조절되지 않고 과하게 발현되는 경우 사이토카인 폭풍(cytokine storm)과 같이 다양 한 조직에 심각한 손상을 주게 된다[7, 12]. 패혈증은 중환자실 (Intensive Care Unit, ICU)의 사망률을 높이는 대표적인 원인 이 되고 있으나, 아직까지 패혈증을 치료하기 위한 효과적이 고 안전한 약물은 없는 실정이다. 일반적으로 패혈증을 제어 하기 위해 항생제와 비스테로이드성 항염증제(non-steroidal anti-inflammatory drug)가 사용되고 있지만 많은 부작용이 동반되고 있다. 항생제의 사용은 사이토카인 생산을 위한 면 역체제를 활성화하여 세균으로부터 LPS의 방출을 가속화 시 키고 이로 인해 내독소(endotoxin)에 의한 쇼크반응들을 유도 하게 된다[20]. 이러한 이유들로 인해 미생물에 대한 활성을
가지며 LPS와 같은 내독소를 중화(neutralization)시키는 기능 을 할 수 있는 새로운 항염증성 신약의 개발이 절실한 상황이다.
항균 펩타이드(antimicrobial peptides, AMPs)는 미생물에 서부터 포유동물에 이르기까지 다양한 종에서 발견되었다.
AMPs는 선천성면역체제에서 매우 중요한 생체방어물질로써 잘 알려져 있으며, 체액과 체표면에 존재하며 염증성 자극에 의해 유도 생성되기도 한다[18]. 이러한 항균 펩타이드들은 기 존의 항감염제제(anti-infective agent)들과 비교하여 강력하고 광범위한 항균활성을 보임과 동시에 미생물들로부터 방출된 병원성 인자(lipopolysaccharide 또는 lipoprotein)들을 중화시 키는 것으로 잘 알려져 있어 중증 패혈증 및 패혈증 쇼크를 예방할 수 있을 것으로 판단된다. 그러나 자연계에서 발견된 항균 펩타이드를 의학적으로 활용하는데 있어 그들이 가지는 적혈구, 신장, 신경세포 등에 대한 독성 때문에 적용하기 쉽지 않은 것이 현실이다[4]. 따라서, 항균 펩타이드를 패혈증 및 다양한 염증성 질환에 안전하게 적용하기 위해 자연으로부터 유래된 항균 펩타이드의 아미노산 서열을 기초로 한 다양한 연구들을 통해 안전하고 효과적인 펩타이드를 고안하는 것이 중요한 과제로 사료된다.
수퍼웜(superworm) 또는 킹웜(kingworm)으로 불리는 아 메리카왕거저리(Zophobas atratus)는 애벌레 몸의 크기가 국내 에서 가장 큰 거저리로 높은 단백질과 지방을 함유하고 있어 애완동물의 먹이와 사료 첨가제로 널리 활용되고 있다[14, 19].
최근에는 아메리카왕거저리의 영양학적인 분석[8]을 토대로 식품의약품안전처로부터 한시적 식품원료로 등록(2020. 01.
16)이 완료된 상태이다. 유엔 식량 농업기구(FAO)에 따르면 네덜란드에서는 거저리과(갈색거저리, 아메리카왕거저리 등) 에 속하는 유충이 인간이 섭취하기에 적합하다고 판단하여 식품으로 판매한다고 보고된 바 있다. 하지만, 아메리카왕거 저리의 생리활성 물질들에 대한 약리학적 연구들은 아직까지 는 미비한 실정이어서 체계적인 연구가 필요하다. 이러한 측 면에서 본 연구에서는 아메리카왕거저리에 대한 기능성 연구 의 일환으로 아메리카왕거저리의 전사체 분석을 통해 선별된 항균 펩타이드 조포바신 1이 LPS로 유도된 대식세포의 염증 반응에서 염증매개인자들의 발현양상과 신호전달과정에서의 인산화 및 LPS와의 상호작용 유무를 조사하였다.
재료 및 방법
실험곤충
아메리카왕거저리(Zophobas atratus)는 국립농업과학원의 곤충사육동에서 계대사육을 통해 유지되어오고 있다. 아메리 카왕거저리 유충의 먹이원은 밀겨를 사용하였으며 25±3℃, 50-70%의 상대습도 조건으로 14L : 10D의 광주기로 사육되었 다.
펩타이드
조포바신 1은 애니젠(주)에서 고상법을 이용하여 합성하였 고, 95% 이상 순도의 펩타이드를 사용하였다. 합성한 펩타이 드는 산성화시킨 증류수(0.01% 아세트산용액)에 녹여서 사용 하였으며 사용 전 까지 -20℃에 보관하였다.
항균활성
조포바신 1의 항균활성은 radial diffusion assay [9] 방법을 통해 테스트하였다. 펩타이드 시료는 0.01% 아세트산용액으 로 준비하였으며 펩타이드 시료의 농도는 일정량으로 serial dilution 하였다. Radial diffusion assay에 사용한 Escherichia coli KCTC 1039, Staphylococcus aureus KCTC 1621, Candida albicans KCTC 7121는 3%(w/v) tryptic soy broth (TSB)에서 37℃, 180 rpm의 조건으로 18시간 배양한 후, E. coli와 S. aur- eus는 대수증식기(log phase)에 들어가도록 동일한 조건에서 2시간 30분간 2차 배양하였다. Citrate phosphate buffer (9 mM sodium phosphate, 1 mM sodium citrate, pH7.4) 와 1%
(w/v) type Ⅰ (low EEO) agarose, 0.03% TSB로 구성된 멸균시 킨 underlay gel에 배양된 미생물(4×106 colony forming units) 을 각각 넣고 혼합해준 뒤 사각플레이트에 붓고 underlay gel 이 굳으면 지름 3 mm의 펀치를 이용하여 구멍을 내어 준비하 였다. Serial dilution을 통해 준비한 농도별 펩타이드들은 5 μl씩 구멍에 넣었다. 시료들이 확산되도록 37℃에서 3시간 동 안 배양한 후, 멸균된 overlay agar (6% TSB, 1% agarose) 10 ml을 붓고 37℃에서 하룻밤 동안 추가 배양하였다. 다음날 미 생물이 성장하지 못한 명확한 영역(clear zone)의 부위들을 확 인하고 지름을 0.1 mm까지 측정하였고 그 값을 units로 표현 하였다(0.1 mm=1 units).
용혈활성
다양한 농도(6.25, 12.5, 25, 50, 100 μg/ml)의 펩타이드 시료 20 μl로 인산완충식염수(phosphate buffered saline, PBS)에 있 는 2.5% (v/v) ICR 마우스 적혈구 용액을 180 μl 넣어 주었다.
벌 독으로부터 분리된 α-나선 구조의 항균 펩타이드인 멜리틴 (melittin)은 적혈구를 파괴하는 양성 대조군으로 사용하였다.
펩타이드와 적혈구의 혼합물은 37℃에서 30분간 반응시킨 후 600 μl의 PBS를 각각의 튜브에 추가로 넣어주었다. 원심분리 기를 이용하여 10,000×g에서 3분간 원심분리 후 상등액을 취 하여 분광 광도계로 540 nm에서 흡광도를 측정하였다.
세포배양
마우스의 대식세포인 Raw264.7 세포주는 Dulbecco’s Modi- fied Eagle Medium (DMEM)에 10% fetal bovine serum (FBS) 과 1% penicillin G-streptomycin을 첨가하여 37℃, 5% CO2
조건하에서 계대배양하고 유지하였다. 마우스 적혈구는 농촌 진흥청 국립농업과학원 동물윤리위원회의 승인을 받아 실험
Table 1. Sequences of primers used for qRT-PCR
cDNAs Primer sequences Accession number
iNOS Forward, 5’-CAGCACAGGAAATGTTTCAGC-3’
Reverse, 5’-TAGCCAGCGTACCGGATGA-3’ NM_010927
COX-2 Forward, 5’-CAGACAACATAAACTGCGCCTT-3’
Reverse, 5’-GATACACCTCTCCACCAATGACC-3’ NM_011198
IL-6 Forward, 5’-GAGGATACCACTCCCAACAGACC-3’
Reverse, 5’-AAGTGCATCATCGTTGTTCATACA-3’ NM_031168
IL-1β Forward, 5’-CCTTCCAGGATGAGGACATGA-3’
Reverse, 5’-TGAGTCACAGAGGATGGGCTC-3’ NM_008361
TNF-α Forward, 5’-ATGAGAAGTTCCCAAATGGC-3’
Reverse, 5’-CTCCACTTGGTGGTGGTTTGCTA-3’ NM_013693
GAPDH Forward, 5’-AAGGTCATCCCAGAGCTGAA-3’
Reverse, 5’-CTGCTTCACCACCTTCTTGA-3’ NM_008084
이 진했되었다(승인번호: NAS-202014).
세포생존율 측정
조포바신 1의 세포독성을 확인하기 위하여 Raw264.7 세포 를 96-well cell culture plate에 2×104 cells/well로 분주하고 18시간 동안 배양하였다. 그 후 조포바신 1을 25, 50, 100, 200 μg/ml의 농도로 처리하여 24시간 동안 추가 배양한 후 MTS (3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethox- yphenyl)-2-(4-sulfophenyl)-2H-tetrazolium) reagent를 사용 하여 세포 생존율을 측정하였다. 흡광도는 multi detector (Beckman, DTX8800, CA, USA)를 이용하여 490 nm 파장에서 측정하였다.
Nitric oxide (NO) assay
Raw264.7 세포로부터 lipopolysaccharide (LPS)에 의해 생 성된 nitric oxide (NO)의 양은 Griess reagent를 이용하여 측 정하였다. 계대배양된 Raw264.7 세포는 8×104 cells/well로 96-well cell culture plate에 분주하여 5% CO2 incubator에서 18시간 동안 배양하였다. NO 생성 유도를 위해 세포에 LPS를 처리하기 전 조포바신 1을 25, 50, 100, 200 μg/ml의 농도로 1시간 동안 전 처리 한 뒤 100 ng/ml의 LPS를 처리하여 24시 간 배양하였다. 배양액의 상등액 100 μl를 취하여 Griess 시약 과 반응 시킨 후 multi detector (Beckman, DTX8800, CA, USA)로 540 nm에서 흡광도를 측정하여 NO 생성량을 측정하 였다. 생성된 NO의 퍼센트는 LPS를 처리한 샘플을 최대 NO 생성량 기준으로 하여 계산하였다.
Quantitative real-time PCR (qRT-PCR)
Raw264.7 세포는 1×106 cells/well로 6-well plate에 분주하 여 5% CO2 incubator에서 18시간 동안 배양한 후 조포바신 1을 25, 50, 100, 200 μg/ml의 농도로 1시간 전처리 한 후에 100 ng/ml의 LPS를 처리하여 17시간 배양하였다. 배양한
Raw264.7 세포는 phosphate buffered saline (PBS)로 2회 세척 하고 TRIzol reagent (Invitrogen, Carlsbad, CA) 1 ml을 첨가 하여 total RNA를 추출하였다. 동량의 RNA (1 μg)로부터 High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster city, CA)를 이용하여 cDNA를 합성하였 고, 염증관련 유전자의 발현은 Table 1에서 제시한 각각의 pri- mer와 함께 AMPIGENE® qPCR Green Mix Lo-ROX (Enzo Life Sciences, USA)를 이용하여 ABI 7500 Real Time PCR System (PE Applied Biosystems, Foster City, CA, USA)으로 확인하였다.
Enzyme-linked immunosorbent assay (ELISA) Raw264.7 세포는 1×106 cells/well로 6-well plate에 분주하 여 5% CO2 incubator에서 18시간 동안 배양한 후 조포바신 1을 25, 50, 100, 200 μg/ml의 농도로 1시간 전처리 한 후에 100 ng/ml의 LPS를 처리하여 24시간 배양하였다. 그 후 배양 액으로부터 유리된 TNF-α, IL-6와 IL-1β를 ELISA kit (Ther- moFisher, Waltham, MA)를 이용하여 측정하였다.
Western blot analysis
Raw264.7 세포는 1×106 cells/well로 6-well plate에 분주하 여 5% CO2 incubator에서 18시간 동안 배양한 후 조포바신 1을 25, 50, 100, 200 μg/ml의 농도로 1시간 전처리 한 후에 100 ng/ml의 LPS를 처리하여 24시간 동안 배양하였다. 그 후 배양액을 제거 한 뒤 M-PER™ Mammalian Protein Extrac- tion Reagent (Thermo)과 protease & phosphatase inhibitor cocktail (Thermo)를 이용하여 세포를 lysis 시킨 후 원심분리 (12,000 rpm, 15 min)하여 상등액을 수거하였다. BCA protein assay kit (Pierce, Rockford, IL, USA)로 정량한 동일량의 단백 질을 sodium dodecyl sulfate-polyacrylamide gel electropho- resis (SDS-PAGE)로 분리한 후, nitrocellulose membrane으로 transfer하였다. Membrane을 5% skim milk로 1시간 동안 반
A B
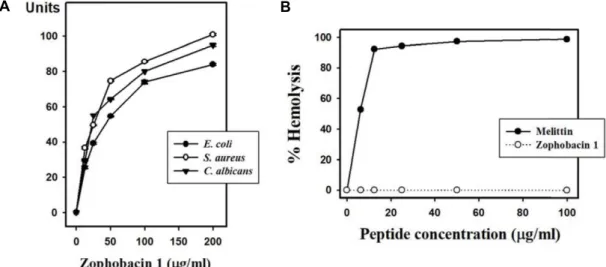
Fig. 1. Antimicrobial and hemolytic activities of zophobacin 1 against microorganisms and mouse erythrocytes determined through radial diffusion assay and hemolysis assay. (A) Zophobacin 1 concentration (x-axis) was plotted against the diameter of the microbial growth inhibition zone (y-axis) after incubation for 16 hr, and is expressed in units (1 mm = 10 units). Data represent the mean ± S.D. with three separate experiments. (B) Melittin was used as a control. Percentage hemolysis was calculated follows: % Hemolysis = (A540 of sample–A540 of peptide-free control)/(A540 of 100% control–A540 of peptide-free control) × 100.
응시켜 비특이적 단백질에 대한 반응성을 차단하고 β-actin, iNOS, COX-2, anti-phospho and total-p44/p42 MAPK, an- ti-phospho and total-p38 MAPK, anti-phospho and total JNK, and IkB (Cell signaling, MA, USA) 항체를 각각 4℃에서 하룻밤 동안 반응시킨 후 horseradish peroxidase (HRP)가 결 합되어 있는 2차 항체로 1시간 반응시켰다. 각 반응 사이에 0.05% TBST로 10분씩 3회 수세 하였다. 그 후 항체에 대한 대응 단백질 band를 Western Lightning Plus (PerkinElmer, Boston, MA, USA)를 사용하여 FluorChem (Alpha Innotech Corporation, San Leandro, CA, USA) 이미지 분석장비로 확 인하였다.
통계처리
모든 실험은 3회 반복하여 측정하였으며, 그 결과는 평균과 표준편차(mean ± SD)로 나타냈다. 실험군 간의 유의성은 Dun- can post-hoc ANOVA analysis를 통해 검정하였고, p<0.05일 때 군 간의 차이가 유의적인 것으로 판단하였다.
결과 및 고찰
조포바신 1의 항균활성 및 용혈활성 확인
아메리카왕거저리 유충의 전사체 분석을 통해 선별한 항균 펩타이드는 조포바신으로 명명하였으며, 선별된 순서에 따라 숫자를 붙여 분류하도록 하였다. 그 중 16개의 아미노산으로 구성된 조포바신 1을 이용하여 미생물에 대한 항균활성을 확 인하였다(Fig. 1A). 그 결과 그람음성균인 대장균(Escherichia coli)과 그람양성균인 황색포도상구균(Staphylococcus aureus)
및 진균인 칸디다 알비칸스(Candida albicans)에 대해 조포바신 1이 농도의존적으로 광범위한 활성을 나타내었다. 또한 ICR 마우스의 적혈구를 이용하여 용혈활성 테스트를 통해 세포독 성이 있는지 확인한 결과 대조군으로 사용된 멜리틴(melittin) 은 12.5 μg/ml에서 약 90%의 용혈활성을 나타낸 반면 조포바 신 1은 최고농도인 100 μg/ml에서도 용혈활성이 없는 것으로 확인되었다(Fig. 1B). C-말단이 아미드화(amidation) 되어 있 는 조포바신 1은 중성 pH에서 총전하(net charge) +6을 갖는 양이온성 항균 펩타이드(cationic antimicrobial peptides, CAPs)이다. 이러한 CAPs는 LPS와 상호작용을 통해 LPS를 중화 시키는 것으로 잘 알려져 있다. LPS를 중화 시키는 기전 은 패혈증 쇼크(septic shock)를 막기 위한 하나의 접근법으로 활용되어 왔다[16]. 그러므로 CAPs는 패혈증과 패혈증 쇼크를 치료하기 위한 가능성 있는 새로운 물질로 사료된다.
조포바신 1의 세포독성 및 Nitric Oxide 생성 억제 효과 확인
많은 연구들에서 다양한 항균 펩타이드가 LPS로부터 활성 화된 대식세포와 동물 모델에서 항염증 효과가 있는 것으로 보고 되어 왔다[1, 5]. 이러한 기능을 하는 항균 펩타이드는 대부분은 양이온 양극성의 펩타이드이며, 펩타이드가 가지는 양전하는 세균의 막 표면 분자들과 상호작용하는데 매우 중요 한 역할을 하는 것으로 밝혀졌다[10]. 조포바신 1이 Raw264.7 대식세포에 대해 세포독성을 나타내는지 확인하기 위해 MTS assay로 세포 생존율을 확인하였다. Raw264.7 세포에 25, 50, 100, 200 μg/ml의 조포바신 1을 처리하고 24시간 뒤에 MTS 시약을 처리하였다. 그 결과 조포바신 1은 200 μg/ml 까지는
A B
Fig. 2. Cell viability and inhibition of NO production in Raw264.7 cells after zophobacin 1 treatment. (A) Cell viability was measured through the MTS assay after 24 hr incubation with the indicated concentrations of the peptides. (B) Raw264.7 cells were pretreated with zophobacin 1 for 1 hr prior to incubation with LPS for 24 hr. The nitrite in the culture media was measured with the Griess reagent. Data represent the mean ± S.D. with three separate experiments. Statistical analyses were performed as described. #p<0.05 in comparison with control group. ***p<0.001, **p<0.01, *p<0.05, compared to the LPS-treated group.
CTR, control; LPS, lipopolysaccharide.
A B
Fig. 3. Inhibitory effects of zophobacin 1 on iNOS and COX-2 expression production in LPS-induced Raw264.7 cells. (A) Raw264.7 cells were pretreated with zophobacin 1 for 1 hr prior to incubation with LPS. Total RNA was isolated 18 hr after LPS treatment. The levels of iNOS and COX-2 mRNA were determined by qRT-PCR. The data were normalized to GAPDH mRNA levels. (B) Raw264.7 cells were pretreated with zophobacin 1 for 1 hr prior to incubation with LPS. Protein was isolated 24 hr after LPS treatment. The protein expression levels of iNOS and COX-2 were determined through Western blot analysis. The data were normalized to β-actin protein levels. Data represent the mean ± S.D. with three separate experi- ments and are representative of three independent experiments. #p<0.05 in comparison with control group. ***p<0.001, **p<0.01,
*p<0.05, compared to the LPS-treated group. CTR, control; LPS, lipopolysaccharide.
세포생존율이 90% 이상 유지하였으며(Fig. 2A), 이후 진행될 항염증 실험들은 200 μg/ml을 최대농도로 하여 실험을 진행 하였다. 조포바신 1이 LPS로 활성화된 Raw264.7 대식세포에 서 nitric oxide (NO)의 생성 정도에 미치는 영향을 확인하였 다. 그 결과 최저농도인 25 μg/ml에서는 NO의 생성을 억제 효과를 보이지 못하였지만 50 μg/ml부터는 농도의존적으로 NO의 생성이 억제되는 것을 확인할 수 있었다(Fig. 2B).
조포바신 1이 iNOS와 COX-2 단백질 발현에 미치는 영향 확인
조포바신 1의 NO 생성억제 효과뿐만 아니라 염증반응의 신호전달과정에서 중요한 매개인자로 알려진 iNOS와 COX-2 의 발현 양상에 미치는 영향을 확인하였다. LPS로 염증이 유 도된 Raw264.7 세포에 25, 50, 100, 200 μg/ml의 조포바신 1을 처리한 결과 iNOS와 COX-2 모두 조포바신 1의 농도에 따라 의존적으로 iNOS와 COX-2의 발현이 억제되는 것을 알 수 있 었다(Fig. 3).
A
B
Fig. 4. Inhibitory effects of zophobacin 1 on the production of proinflammatory cytokines in LPS-induced Raw264.7 cells. (A) Raw264.7 cells were pretreated with zophobacin 1 for 1 hr prior to incubation with LPS. Total RNA was isolated at 18 hr after LPS treatment. The levels of IL-6 and IL-1β mRNA were determined through quantitative real-time-PCR. The data were normalized to GAPDH mRNA level. (B) Raw264.7 cells were pretreated with zophobacin 1 for 1 hr prior to incubation with LPS for 24 hr. IL-6 and IL-1β levels in the culture media were measured through ELISA. Data represent the mean ± S.D. with three separate experiments. #p<0.05 in comparison with control group. ***p<0.001, **p<0.01, *p<0.05, compared to the LPS-treated group. CTR, control; LPS, lipopolysaccharide.
조포바신 1이 염증성 사이토카인 발현에 미치는 영향 확인 LPS로부터 유도된 염증반응은 사이토카인들이 발현되어 분비되는 일련의 과정들이 발생하는데, 이러한 인자들이 조절 되지 못하고 과도하게 분비되면 조직의 손상 및 죽음에 이르 는 원인이 되기도 한다[7, 12]. 그러므로 적절한 수준으로 염증 반응이 조절되는 것은 매우 중요한데 조포바신 1이 전염증성 사이토카인(proinflammatory cytokine)인 TNF-α, IL-6 그리고 IL-1β의 발현을 저해할 수 있는지 qRT-PCR과 ELISA 실험을 통해 유전자 및 단백질 수준에서 각각 확인하였다. 먼저 유전 자 수준에서 분석한 결과 IL-6와 IL-1β의 경우에는 25 μg/ml 의 농도부터 농도의존적으로 전염증성 사이토카인의 발현 저 해 효과를 나타내었다(Fig. 4A). 또한 ELISA를 통해 단백질 수준에서도 저해되는지 확인하였으며, 그 결과 IL-6와 IL-1β 모두 유전자 수준과 유사하게 25 μg/ml의 농도부터 농도의존 적으로 저해효과를 나타내었다(Fig. 4B). 그러나 TNF-α의 경
우에는 유전자와 단백질 수준에서 모두 현저한 저해효과를 확인하지는 못하였다.
조포바신 1과 LPS의 상호작용 확인
항균 펩타이드가 항균 및 항염증 활성을 나타내는데 있어 LPS와의 결합은 매우 중요한 것으로 알려져 있다. 항균 펩타 이드들이 가지는 소수성 아미노산들과 전하값은 세균의 인지 질 막(phospholipid membrane) 성분과 항균 펩타이드 사이의 상호작용 가능성을 결정하는데 있어 중요한 역할을 하게 된 다. 양전하의 항균 펩타이드들은 그람음성균의 막에 음전하를 띄는 LPS와 전기적 상호작용을 통해 강한 결합을 하게 되며 이로 인해 음전하가 작동하지 못하도록 막는 역할을 하며, 소 수성 아미노산들은 항균 펩타이드가 LPS로 구성된 마이셀 (micelle)로 쉽게 삽입될 수 있도록 해준다[20]. Y. Rosenfeld, Scott et al. 등의 연구진은 아미노산의 소수성 아미노산을 친
Fig. 5. Specific binding of zophobacin 1 to lipopolysaccharide. The radial diffusion assay was conducted by mixing varying amounts of LPS with 200 μg/ml of zophobacin 1. The mixtures of peptide with LPS were loaded into wells of the assay plate seeded with E. coli, which has been confirmed to be highly susceptible to zophobacin 1. The upper panel shows a photo of the gel from the radial diffusion assay. Numbers (x-axis) represent the LPS concentration (mg/ml) of the mixture load- ed in the wells. Five μl of 200 μg/ml zophobacin 1 was used for LPS-free control. The lower panel shows the antibacterial activity of zophobacin 1 in the mixture plotted against the con- centration of LPS. The diameters of the clearing zone are ex- pressed in units (1 mm = 10 units). Data represent the mean
± S.D. with three separate experiment. Statistical analyses were performed as described. ***p<0.001, **p<0.01, *p<0.05, com- pared to the LPS non-treated control. LPS, lipopolysaccharide.
A B
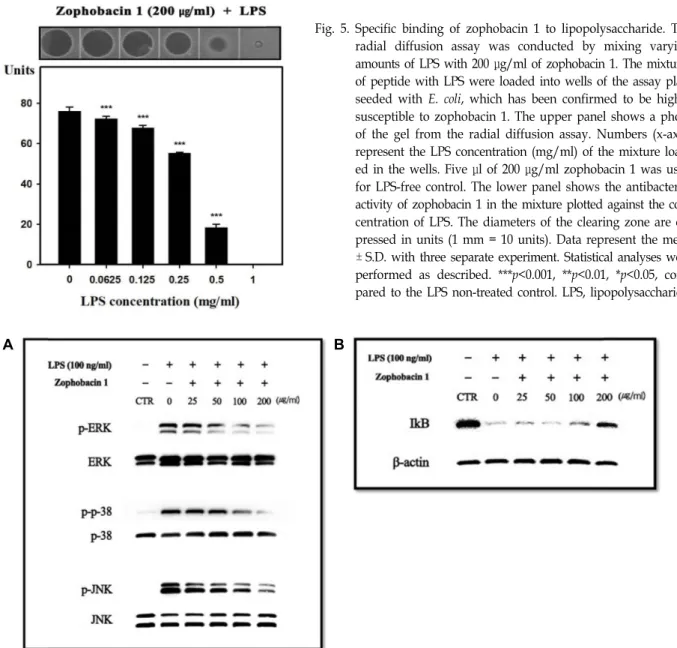
Fig. 6. Inhibitory effect of zophobacin 1 on MAPKs and NF-kB signaling pathways in Raw264.7 cells. (A) Raw264.7 cells (1×106 cells/well in a 6-well plate) were incubated with the indicated concentration of zophobacin 1. Protein was then isolated at 30 min after LPS treatment, and phosphorylation of ERK, p38, and JNK was detected using Western blot analysis. The data were normalized to total protein, respectively. (B) Degradation of IκB was detected using Western blot analysis. The data were normalized to β-actin. CTR, control; LPS, lipopolysaccharide.
수성 아미노산으로 치환하거나 양이온 아미노산으로 치환한 경우 LPS에 대한 결합력이 더욱 증가하는 것을 증명하였다 [13, 15, 17]. 이러한 관점에서 조포바신 1이 LPS와 결합을 통해 내독소를 중화시킴으로써 염증반응을 제어할 수 있는지 여부 를 확인하였다. 조포바신 1과 LPS의 결합 유무는 radial dif- fusion assay를 통해 조사하였으며, 조포바신 1의 농도를 200 μg/ml로 고정하고 LPS의 농도를 증가시킴에 따라 항균활성 이 저해되는지 확인하였다. 그 결과 LPS가 존재하지 않을 경 우 강한 항균활성으로 clear zone이 잘 형성된 반면 LPS의 농 도가 증가함에 따라 농도의존적으로 조포바신 1의 활성이 감
소되어 사라지는 것을 알 수 있었다(Fig. 5). 이러한 결과는 조포바신 1이 LPS와 강하게 결합한다는 것으로 보여주는 증 거로 과도한 염증반응을 제어하기 위해 항균 펩타이드가 사용 될 수 있음을 보여주는 증거라 할 수 있다.
조포바신 1이 Raw264.7 세포에서 MAPKs 인산화에 미 치는 영향 확인
전염증성 사이토카인 매개인자의 생성에서 mitogen-acti- vated protein kinases (MAPKs)는 중요한 조절자로서 작용한 다[11]. 그러므로 조포바신 1이 Raw264.7 세포에서 LPS로부터
유도된 MAPKs의 인산화에 어떠한 영향을 주는지 Western blot analysis를 통해 확인하였다. Raw264.7 세포로 조포바신 1을 1시간 동안 전처리 한 후 LPS를 30분 동안 처리하여 염증 을 유발하였다. 그 결과 MAPKs의 ERK, p38, JNK의 인산화는 LPS에 의해 유도 발현되지만 조포바신 1의 농도가 증가함에 따라 인산화 정도가 농도의존적으로 감소하는 결과를 보였다 (Fig. 6A). NF-κB 신호전달과정의 활성화는 IκB kinase (IKK) complex에 의해 매개되며, IKK에 의해 분해되는 IκB의 정도 를 관찰함으로써 염증조절 여부를 판단할 수 있다. 앞선 방법 과 동일하게 염증을 유발한 후 조포바신 1을 처리한 결과 LPS 에 의해 IκB의 분해가 최대인 것에 비해 조포바신 1의 최고 농도인 200 μg/ml에서 IκB의 분해가 감소하는 것을 알 수 있 었다(Fig. 6B). 본 연구에서 수행된 결과들을 종합했을 때 이러 한 결과들은 항균 펩타이드인 조포바신 1이 패혈증과 같은 염증성 질환에 도움을 줄 수 있는 물질로 개발될 수 있을 것이 라 기대된다.
감사의 글
본 성과물은 농촌진흥청 차세대바이오그린21사업(세부과 제번호: PJ01325601)의 지원에 의해 이루어진 것임.
The Conflict of Interest Statement
The authors declare that they have no conflicts of interest with the contents of this article.
References
1. Brandengurg, K., Andra, J., Garidel, P. and Gutsmann, T.
2011. Peptide-based treatment of sepsis. Appl. Microbiol.
Biotechnol. 90, 799-808.
2. Breithaupt, H. 1999. The new antibiotics. Nat. Biotechnol. 17, 1165-1169.
3. Duque, G. A. and Descoteaux, A. 2014. Macrophage cyto- kine: Involvement in immunity and infectious diseases.
Front. Immunol. 5, 491.
4. Gordon, Y. J., Romanowski, E. G. and McDermott, A. M.
2005. A review of antimicrobial peptides and their ther- apeutic potential as anti-infective drugs. Curr. Eye. Res. 2005, 505-515.
5. Heinbockel, L., Marwitz, S., Varela, S. B., Espada, R. F., Reiling, N., Goldmann, T., Gutsmann, T., Mier, W., Schurholz, T., Dromann, D., Brandenburg, K. and de Tehada, G. M. 2015.
Therapeutical administration of peptide Pep19-2.5 and ibu- profen reduces inflammation and prevents lethal sepsis.
PLoS One 10, e0133291.
6. Janeway, C. A. Jr. and Medzhitov, R. 1998. Introduction: the
role of innate immunity in the adaptive immune response.
Semin. Immunol. 10, 349-350.
7. Karima, R., Matsumoto, S., Higashi, H. and Matsushima, K. 1999. The molecular pathogenesis of endotoxic shock and organ failure. Mol. Med. Today 5, 123-132.
8. Kim, S. Y., Kim, H. G., Ko, H. J., Kim, I. W., Seo, M. C., Lee, J. H., Lee, H. J., Baek, M. H., Hwang, J. S. and Yoon, H.
J. 2019. Comparative analysis of nutrients and hazardous substances in Zophobas atratus larvae. J. Life Sci. 29, 1378- 1385.
9. Lehrer, R. I., Rosenman, M., Harwig, S. S., Jackson, R. and Eisenhauer, P. 1991. Designer assays for antimicrobial pep- tides. J. Immunol. Methods 137, 167-173.
10. Malmsten, M. 2016. Interactions of antimicrobial peptides with bacterial membranes and membrane components. Curr.
Top. Med. Chem. 16, 16-24.
11. Marina, L., Kamal, R. M., Andrew, F., Gary, B., Jeremy, S., and Andrew, R. C. 2000. Regulation of cyclooxygenase 2 mRNA stability by the mitogen-activated protein kinase p38 signaling cascade. Mol. Cell. Biol. 20, 4265-4278.
12. Martin, G. S., Mannino, D. M., Eaton, S. and Moss, M. 2003.
The epidemiology of sepsis in the United States from 1979 through 2000. N. Engl. J. Med. 348, 1546-1554.
13. Monisha, G. S., Yan, H. and Hancock, R. E. W. 1999. Biologi- cal properties of structurally related α-helical cationic anti- microbial peptides. Infect. Immun. 67, 2005-2009.
14. Park, H. C., Jung, B. H., Han, T. M., Lee, Y. B., Kim, S.
H. and Kim, N. H. 2013. Taxonomy of introduced commer- cial insect, Zophobas atratus (Coleoptera; Tenebrionidae) and a comparision of DNA barcoding with similar tenebrionids, Promethis valgipes and Tenebrio molitor in Korea. J. Seric.
Entomol. Sci. 51,185-190.
15. Rosenfeld, Y., Lev, N. and Shai, Y. 2010. Effect of the hydro- phobicity to net positive charge ratio on antibacterial and anti-endotoxin activities of structurally similar antimicrobial peptides. Biochemistry 49, 853-861.
16. Rosenfeld, Y. and Shai, Y. 2006. Lipopolysaccharide (endotoxin)-host defense antibacterial peptides interactions:
role in bacterial resistance and prevention of sepsis. Biochim.
Biophys. Acta. 1758, 1513-1522.
17. Srivastava, S. and Ghosh, J. K. 2013. Introduction of a lysine residue promotes aggregation of temporin L in lipopoly- saccharides and augmentation of its antiendotoxin property.
Antimicrob. Agent Chemothera. 57, 2457-2466.
18. Steinstraesser, L., Kraneburg, U. M. and Hirsch, T. 2009.
Host defense peptides as effector molecules of the innate immune response: a sledgehammer for drug resistance? Int.
J. Mol. Sci. 10, 3951-3970.
19. Tschinkel, W. R. 1984. Zophobas atratus (Fab) and Z. rugipes Kirsch (Coleoptera: Tenebrionidae) are the same species.
Coleopts. Bull. 38, 325-333.
20. Yue, S. and Dejing, S. 2015. Inhibitory effects of anti- microbial peptides on lipopolysaccharide-induced inflam- mation. Mediators Inflamm. 2015, 167572.
초록:아메리카왕거저리 유래 항균 펩타이드 조포바신 1의 항염증활성
신용표†․이준하†․김인우․서민철․김미애․이화정․백민희․김성현․황재삼*
(농촌진흥청 국립농업과학원 농업생물부 곤충산업과)
본 연구에서는 아메리카왕거저리에 대한 기능성 연구의 일환으로 아메리카왕거저리 유충의 유전체 분석을 통 해 선별된 조포바신 1의 항균 및 항염증 활성을 확인하였다. 선행연구에서 RNA 시퀀싱을 통해 아메리카왕거저리 의 전사체를 분석하였으며, 결과를 바탕으로 인실리코(in silico) 분석을 수행하여 전사체 유래 항균 펩타이드를 스크리닝하고 선발하였다. 수행된 항균활성 및 용혈활성 테스트에서 조포바신 1은 세균 및 칸디다 진균에 대해 광범위한 항균활성을 나타낸 반면 마우스 적혈구에 대한 용혈활성은 전혀 없었다. 다음으로 마우스 대식세포주 Raw264.7 세포를 이용하여 조포바신 1의 항염증활성을 확인하였다. 그 결과 조포바신 1은 LPS로부터 유도된 Raw264.7 세포들의 산화질소 생성을 감소시키는 결과를 보여주었다. 뿐만 아니라 실시간 역전사 중합효소 연쇄반 응(qRT-PCR) 방법과 효소결합면역흡착측정법(ELISA)을 통해 조포바신 1이 Raw264.7 세포에서 전염증성 사이토 카인(IL-6, IL-1β)의 발현을 감소시킨다는 것을 확인할 수 있었다. 또한 염증반응의 신호전달인자들(MAPKs, NF-κ B)의 인산화를 억제하는 것을 확인하였다. 게다가 조포바신 1은 LPS와의 상호작용을 통해 결합한다는 것을 확인 하였다. 이러한 연구결과들은 아메리카왕거저리 유전체 분석을 통해 확인된 조포바신 1이 항균 및 항염증 치료를 위한 물질로서 개발하는데 가능성이 있을 것으로 사료된다.