위암에서 새로운 종양원인 유전자 Nemo-like Kinase의 발현 증가
김민규*,**·정광화*,**·남석우*,**,#
*가톨릭대학교 의과대 가톨릭대학교 의과대학 병리학교실, **가톨릭대학교 기능전사체 연구소 (Received September 10, 2012; Revised November 14, 2012; Accepted November 19, 2012)
The Overexpression of Oncogenic Nemo-like Kinase in Gastric Cancer
Min Gyu Kim*,**, Kwang Hwa Jung*,** and Suk Woo Nam*,**,#
*Department of Pathology, College of Medicine, The Catholic University of Korea, Seoul 137-701, Korea
**Functional RNomics Research Center, The Catholic University of Korea, Seoul 137-701, Korea
Abstract − Nemo-like kinase (NLK), an evolutionarily conserved serine/threonine protein kinase, plays an important role in wide variety of developmental events. NLK phosphorylates T-cell factor/lymphoid enhancer factor (TCF/LEF) tran- scriptional complex and suppresses wnt signaling pathway through inhibition of β-catenin/TCF complex interaction. How- ever, the function of NLK in gastric carcinogenesis has not been investigated. In the present study, we have examined whether the NLK gene is involved in the development and/or progression of gastric cancers. NLK expression was analyzed by immunohistochemical staining in 153 advanced gastric cancer specimens. Immunhistochemical analysis showed increased expression of NLK in 91 (59.5%) out of 153 gastric cancer specimens. Statistically, there was no significant rela- tionship between altered expression of NLK protein and clinicopathological parameters, including tumor differentiation, location, lymph node metastasis. We identified that mRNA and protein expression of NLK was significantly up-regulated in human gastric cancer tissues compare to corresponding normal gastric tissues. In addition, we found that human gastric can- cer cell lines exhibited relatively high expression of NLK, as compared with normal gastric cells. The results of this study suggest that aberrant regulation of NLK may contribute to the development or progression of gastric cancers and serve as a potential biomarker for advanced gastric cancer patients.
Keywords □ NLK expression, gastric cancer, immunhistochemistry
위암은 세계적으로 암과 관련된 사망 원인 중 두 번째를 차지 하고 있으며, 한국에서도 가장 높은 발생률을 차지하고 있고 악 성 종양으로 인한 사망 원인 중에서도 두 번째를 기록할 정도로 흔한 종양이다.1,2)위암은 암 종의 특징상 발생초기 자각증상이 없어 조기 진단이 어렵고, 대부분 위암진단 시 이미 상당한 진행 성 암 종으로 진단되기 때문에 치료방법이 매우 제한되어있고, 예후도 극히 나쁘다. 따라서 위암 초기 발생기전을 이해하고 이 를 이용한 빠른 진단과 효과적인 치료 수단을 위한 새로운 항암 요법 개발이 요구된다.
위암에서 가장 중요한 예후인자는 조직학적 병기이지만, 동일 병기 내에서도 재발성과 전이 정도가 다르기 때문에 동일병기의 환자들에게 동일한 치료를 하여도 서로 다른 임상 결과를 보이
는 경우가 많다. 따라서 위암의 예후 예측이나 화학치료 여부를 결정하는데 중요한 분자생물학적 인자들에 관한 많은 연구가 진 행 중이지만 위암 발생 과정에 관여하는 예후 인자나 분자유전 학적 기전들은 아직 명확히 밝혀지지 않고 있다.
NLK(Nemo-like kinase)는 Extracellular signal-regulated kinase/MAPK(Erk/MAPK)와 cyclin-dependent kinases(CDKs) 의 구성원이며, MAPKK(MAPK kinase kinase) super family인 TAK1(transforming growth-factor-b-activated kinase 1)이 Wnt signaling pathway에서 NLK의 잠재적 활성자로서 알려져 있다.3,4) 꼬마선충 및 초파리의 유전적 연구에서는, Nemo가 척추동물의 NLK와 상동성을 가지는 것으로 알려졌고, 특히, 꼬마선충의 세 포분열 및 초파리 날개의 발생동안 Wnt signaling의 조절자로 기 능함이 알려진 바 있다.5,6)따라서 NLK는 세포성장, 정형화 및 다양한 세포사멸에 매우 중요한 조절자로 작용한다.
NLK는 wnt/β-catenin signaling pathway에서 TCF/LEF(T- cell factor/lymphoid enhancer factor)를 인산화시켜 β-catenin/
#본 논문에 관한 문의는 저자에게로
(전화) 02-2258-7314 (팩스) 02-537-6586 (E-mail) [email protected]
종설
TCF complex의 inter-action을 방해 함으로서 전사 활성을 억제 하며,7,8) 또한 NLK는 여러 신호전달 경로에서 다양한 전사 인자 들의 활성을 조절하는 것으로 잘 알려져 있다.9,10)최근 연구에 따르면, 대장암(colon cancer) 및 담낭암(gallbladder cancer) 세 포에서의 NLK 과발현은 세포주기전괴, 세포사멸 및 세포이동성 을 조절 함으로서 종양세포의 증식을 억제하여 종양 억제자로서 의 역할을 한다고 보고하였다.11,12)이와는 반대로 본 연구자 그 룹의 최근 연구에서는 간암(liver cancer)에서 NLK의 발현은 증 가되어 있음을 확인하였고, 간암세포에서 NLK를 선택적으로 억 제 하였을 때 세포주기 관련 단백질들을 조절 함으로서 종양세 포의 증식을 억제한다는 것을 확인하였다.13)
이와 같이 NLK는 여러 암 종의 발생 및 진행과정에서 중요한 역할을 한다고 알려져 있지만, 위암에서 NLK의 분자생물학적 기전에 대한 연구와 신호전달 체계에 대해서는 지금까지 알려진 바가 없다. 따라서 본 연구에서는 위암에서 NLK의 비 특이적인 조절이 종양발생과 진행에 관여하는지를 알아보기 위해, 위암 환자 조직을 대상으로 NLK 발현 변화에 대한 연구를 수행하 였다.
실험방법
조직 시료 및 면역조직화학염색
면역조직화학염색을 위하여 153 case의 파라핀 포매 된 위암 조 직을 이용하여 Tissue Microarray Block을 제작하였다. 2명의 해 부병리 의사가 독립적으로 조직 절편을 확인하여 병소를 구분하 였고, tissue microarray block 제작 기구(Beecher Instruments, Micro-Array Technologies, USA)를 이용하여, 2 mm 지름으로 암 종에서는 두 개의 core를 채취하고 정상 위점막에서는 하나 의 core를 채취하여 tissue microarray(TMA)를 제작하였다. 그 리고, 단면을 고르게 하기 위해서 조직 미세배열 블록을 50oC 보 온기에서 20분간 방치하였다. 환자의 성별, 연령 및 진단을 제외 한 개인 정보는 모두 익명으로 처리 하였으며, 연구 계획은 가톨 릭 의과대학 기관윤리위원회의 심의 및 승인을 받았다 (CUMC09U116). 그 다음 TMA 파라핀 블록을 3 µm 두께의 절 편으로 박절하여 슬라이드에 부착한 후 자일렌에 5분간 3회에 걸쳐 탈파라핀 시키고, 100%, 90%, 80%, 70% 알코올에 각각 5 분간 순서대로 처리하고 증류수로 씻어냈다. 또 항원성 회복을 위해 TMA 슬라이드를 구연산염 완충용액(10 nM, pH 6.0)으로 15분을 끓인 다음 20분간 냉각한 후, 이차 증류수로 씻어냈다.
조직 내의 내인성 과산화효소를 제거하기 위해 3% 과산화수소 용액에 담가 실온에서 15분간 반응시켰다. 일차 항체로 NLK (SantaCruz, USA)응 1 : 100으로 희석한 후, 4oC에서 16~18시 간 동안 반응시킨 다음 biotin이 부착된 이차 항체(biotinylated anti-rabbit immunoglobulin antibody, Sigma, USA)를 상온에서
1시간 반응시켰다. 그리고 세척 완충액으로 5분씩 3회 씻어낸 후 biotinyl tyramide signal amplification system(PerkinElmer, USA)을 이용하여 biotin-tyramide 특이 결합을 유도하였다. 그런 다음 DAB(3,3'-diaminobenzidine tetrahydrochloride solution, Dako, USA)으로 3~5분간 반응시켜 발색시키고, Mayer's hematoxylin으로 대조 염색하였다.
유전자 발현 Microarray 분석
위암에서 NLK의 mRNA 발현 양상을 알아보기 위하여 National Center for Biotechnology Information(NCBI) Gene Expression Omnibus(GEO) database에서, microarray 실험을 통하여 보고된 큰집단의 위암 환자조직에서의 유전자 발현 데이 터를 획득하여 NLK의 유전자 발현양상을 확인하였다(accession numbers GSE13195, GSE27342).
세포 배양
인간 정상 위점막세포인 GES-1과 MKN-1, MKN-28, MKN- 45, AGS, SNU-216, SNU-484, SNU-719 인간위암세포를 실험 에 사용하였다. 세포들의 배양을 RPMI 1640에 열처리한 FBS를 10% 처리하고, penicillin과 streptomycin을 각각 100 unit/ml과 100µg/ml의 농도로 처리하여 37, 5% CO2배양기에서 배양하였 다. 일 주일에 두 번씩 1 : 4의 비율로 계대 배양하였다.
QRT-PCR (Quantitative Real-Time Polymerase Chain Reaction)
세포 plate에서 배양된 정상 위세포 및 위암세포에 Trizol reagent(Invitrogen, USA) 1 ml을 분주하여, 세포 용해물을 즉시 튜브로 옮긴 다음, 클로로포름을 200 µl를 넣어서, 4oC에서 13,000 rpm으로 15분 동안 원심분리 하였다. 원심분리 후 얻은 상층액과 이소 프로필 알코올을 첨가하여 섞고 상온에서 10분간 반응시키고 13,000 rpm에서 10분 동안 원심분리 하였다. 원심분 리하여 생긴 침전물을 75%의 알코올을 첨가하여 vortex한 후 13,000 rpm에서 다시 5분간 원심분리하여 용액을 덜어낸 후 침 전물을 공기 중에서 말리고, 각각의 튜브마다 RNase free water 를 첨가하여 RNA를 얻었고 Nanodrop Spectrophotometer (Thermo scientific, USA)을 이용하여 total RNA의 농도를 측정 하였다. RNA의 quality check는 Experion Automated Electro- phoresis Station(Bio-Rad, USA)을 이용하여 수행하였고, 1 µg의 Total RNA를 Reverse transcription kit(Roche, USA)을 이용하 여 cDNA로 만든 후 NLK 및 GAPDH 프라이머를 이용하여 PCR을 수행하였다. 프라이머에 대한 정보는 다음과 같다: NLK forward 5'-CTT TGA GCC TGT CAC CAA TCC CAA-3', reverse 5'-CTT CCA CCA TCA CTC CCA CAC CA-3';
GAPDH forward 5'-ACC AGG TGG TCT CCT CTG AC-3',
reverse 5'-TGC TGT AGC CAA ATT CGT TG-3'. 증폭반응을 위하여, PCR을 40회 실시(IQ5 Real Time PCR Machine, Bio- Rad, USA) 하였으며, denaturation은 94oC에서 30초, annealing 은 60oC에서 15초, extension은 72oC에서 15초간 각각 시행하 였다.
단백질 발현 Western blot 분석
NLK 유전자의 단백질발현을 알아보기 위하여 위암 환자 조직 및 위암 세포를 lysis buffer(50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% nonidet P-40, 0.25% sodium deoxycholate, 1 mM Phenylmethane-sulfonylfluoride)를 이용하여, 1 ml 주사기로 세 포를 분쇄한 후 균질화 시켰다. 단백질 정량은 BCA protein assay reagent(Thermo scientific, USA)를 이용하여 측정하였다. 정량 한 단백질 15~20 µg을 10% SDS-PAGE로 분리한 후 gel을 membrane(Bio-Rad, USA)에 이동시키고 5% skim milk로 상 온에서 1시간 blocking한 후 1 : 500 비율로 희석시킨 primary NLK antibody(SantaCruz, USA)와 4oC에서 overnight하였다.
다음으로 Tris-buffered saline tween-20(TBS-T)으로 5분 동안 3번 세척한 후 1 : 5000의 비율로 희석시킨 일차 NLK 항체 (Sigma, USA)와 상온에서 1시간 반응시켰다. 이 후 TBS-T로 5 분 동안 3번 세척하고 ECL 용액(Millipore, USA)을 이용하여 단백질 발현에 대한 검출은 LAS-4000(Fujifilm, Japan)을 사용 하였다.
통계학적 분석
모든 실험결과는 평균±표준편차(Mean±SD)로 나타내었다. 대 조군과 비교하여 통계 유의성Student's t-test로 처리하였으며 P < 0.05 이하인 경우 유의성 있는 차이가 있는 것으로 판정하 였다.
실험결과
면역조직화학염색에 의한 NLK 단백 발현
면역조직화학염색 결과 NLK는 정상 위점막세포와 종양세포 의 핵 및 세포질에 주요하게 염색된 것이 관찰되었고(Fig. 1), 위암 153예 중 91예(59.5%)에서 강한 양성발현을 보였다(Table I). 위암의 Lauren 분류법에 따라서 intestinal type 55예 중 35예(63.6%), diffuse type 67예 중 42예(62.7%), mixed type 31예 중 14예(45.2%)에서 NLK의 양성발현이 각각 관찰되 었고, 림프절 전이가 확인되는 150예 중 90예(60%)에서 NLK 의 과발현이 확인되었다. 그러나 위암에서의 NLK의 과발현 과 병소의 분류, 전이, 위치, 크기와 같은 임상 병리학적 인자 들과의 상관관계는 통계학적으로 유의성이 없다는 것을 확인 하였다.
위암조직에서의 NLK mRNA 및 단백 발현
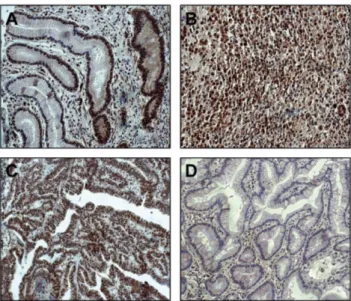
위암조직에서 NLK의 mRNA 발현 양상을 알아보기 위하여 NCBI GEO database에서, 보고된 두 종류의 large cohort의 위 암 환자조직에서의 유전자 발현 dataset에서 NLK 유전자 발현 양상을 확인하였다(Fig. 2A and 2B). 결과에서 볼 수 있는 것처 럼, NLK 유전자 발현은 두 개의 GEO database 모두에서 위암 조직에서 정상 위점막조직에 비해 통계적으로 유의성 있게 증가 되는 것을 확인하였다. 또한 Western blot을 이용한 단백질 발 Fig. 1− NLK immunostaining of gastric cancers. (A) Positive staining for NLK was seen in non-cancerous gastric epithelial cells (B and C) Strong immuno-reactivity was found in diffuse- and intestinal-type gastric cancers. (D) Example of intestinal-type gastric cancer with negative NLK immuno-staining.
Table I− Relationship between the expression of NLK protein and the clinical/pathological parameters
Parameter NLK expression
P value
+ -
Lauren†
Intestinal Diffuse Mixed
35 42 14
20 25 17
0.190
Lymph node metastasis†
0 1~5≥6
01 27 63
02 15 45
0.361
Site†
Upper Middle Lower
02 36 53
00 30 32
0.313
Size†
<6.5 cm
≥6.5cm 51
40
34 28
0.882
Total 91 62
현에서도, 임의로 선정한 10명(Patient No. 1-10)의 위암 환자의 조직 중 6명의 위암환자 조직(Patient No. 2, 3, 4, 6, 8, 9)에서 의 NLK 단백질의 발현이 정상 위점막조직과 비교하여 증가 되 는 것을 확인하였다(Fig. 2C).
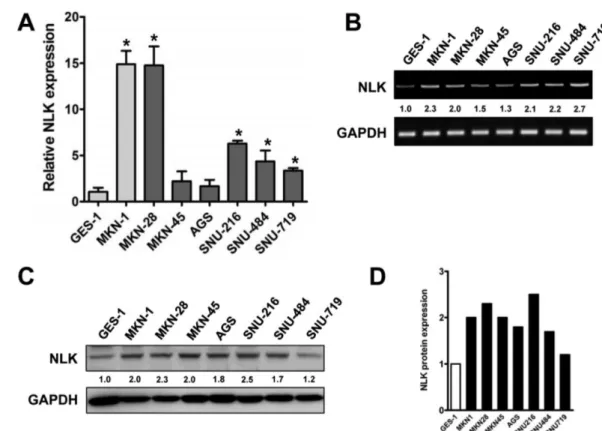
정상 위점막세포와 위암세포주에서의 NLK mRNA 및 단백 발현 정상 위점막세포인 GES-1과 7개의 서로 다른 위암세포주 (MKN-1, MKN-28, MKN-45, AGS, SNU-216, SNU-484, SNU- 719)에서 NLK의 mRNA 발현양상을 확인하기 위해서 quantitative real-time reverse transcriptase PCR(QRT-PCR)을 수행하였다 (Fig. 3A and 3B). 결과에서 볼 수 있듯이 위암세포주 MKN-1, MKN-28, MKN-45, AGS, SNU-216, SNU-484, SNU719에서의 NLK의 mRNA 발현이 정상 위점막세포인 GES-1보다 증가되어 있음을 확인하였고, 앞선 결과와 마찬가지로 NLK 단백의 발현 정도를 Western blot 분석하였을 때도, 실험에 사용된 모든 위암 세포주에서 NLK 단백질 발현이 정상 위점막세포와 비교하여 증
가되어 있다는 것을 관찰할 수 있었다(Fig. 3C and 3D).
고찰 및 결론
위암은 한국, 일본, 중국 등 아시아에서 높은 빈도를 차지하고 있는 암으로서 세계적으로 암과 관련된 주요한 사망 원인 중에 하나이다.1)그러나 위암의 발생과 진행에 관련된 분자적인 메커 니즘은 아직 많은 부분이 잘 알려져 있지 않다.
NLK는 serine/threonine protein kinase로서 암에 밀접하게 연 관되어 있는 wnt/beta-catenin signalling pathway를 조절하는 것 으로 잘 알려져 있고,7) 또한 NF-κB, Smads, AP1, p53 그리고 STAT3 같은 여러 전사 인자들의 인산화를 통하여 다양한 signaling pathways에서 중요한 역할을 한다고 알려져 있다.9,14-16) 여러 연구자 그룹에서 암의 발생과 진행과정에서 NLK의 조절 기작에 대한 연구를 수행하였고, 그 결과 대장암세포인 DLD-1 세포주에서의 NLK 과발현은 세포주기 관련 인자들을 조절함으 Fig. 2− Expression of NLK in gastric cancer tissues. (A and B) Recapitulated NLK gene expression levels of the large cohort of HCC patients.
Two sets of microarray data were obtained from the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO) database (accession numbers GSE13195 and GSE27342). The relative expression of NLK mRNA in normal gastric mucosa (Normal) and gastric cancer tissues (Tumor) is illustrated by scatter plot. The differential NLK expression for these two categories was determined by the unpaired t test (*P<0.05). (C) Analysis of NLK expression in human gastric cancer and adjacent non-cancerous tissues by Western blot. T, gastric cancer tissue; N, adjacent non-cancerous tissue. The α-tubulin was used as internal loading controls. (D) The graph bar shows the density of NLK protein expression relative to α-tubulin
로서, DLD-1세포의 증식을 억제한다는 것과,11)담낭암세포인 GBC-SD과 SGC-996 세포주에서의 NLK 과발현은 암 진행의 필 수 과정인 세포의 이동과 증식을 억제함을 보고하였다.12)또한 난소암(ovarian cancer) 조직에서는 NLK의 발현이 감소되어 있 고, NLK의 발현은 난소암 환자의 예후를 결정하는 중요한 인자 라는 것을 확인할 수 있었다.17)하지만 이와는 반대되는 결과로 서, 간암에서는 NLK 발현이 증가되어 있고, Hep3B 간암세포주 에서 NLK의 억제는 G1/S cell cycle arrest를 야기시켜 세포의 증식을 억제한다는 것을 알 수 있었다.13)따라서 이와 같은 연구 들은 서로 다른 암 종의 발생 및 진행과정에서 NLK의 복잡한 조절기작과 분자적인 메카니즘을 부분적으로 보여주는 중요한 결과다.
본 연구에서는 위암에서 NLK의 발현 양상을 조사하였고, 위 암조직에서의 면역조직화학염색을 통하여 위암 153예 중 91예 (59.5%)에서 NLK가 과발현 되어 있음을 확인하였다(Fig. 1). 또 한 QRT-PCR 및 Western blot 분석을 통해서도 위암환자 조직 및 위암세포주에서 정상 위점막조직 및 세포에 비해 NLK가 증 가 되어 있음을 알 수 있었다(Fig. 2). 그러나 위암에서의 NLK 의 과발현은 병소의 분류, 전이, 위치, 크기와 같은 임상 병리학
적 인자들과는 통계학적으로 유의성이 없는 없다는 것을 확인하 였다(Table I). 한편, 간암에서도 NLK 발현에 대한 위의 내용과 비슷한 연구 결과가 도출이 되었고,13)따라서 다음과 같은 결과 들은 위암의 진행에서도 NLK가 세포 주기를 조절할 가능성을 시사하는 결과라고 판단된다.
이상의 연구 결과를 요약하면 NLK의 비정상적 과발현은 위 암의 발생 및 진행과정에서 밀접하게 연관되어 있음을 알 수 있 었고, 앞으로 위암에서 NLK의 작용기전을 규명하기 위해 보다 광범위한 추가 연구가 필요할 것으로 생각된다. 본 연구는 그 동 안 밝혀지지 밝혀 지지 않았던 위암에서의 NLK의 발현을 연구 함으로서 향후 새로운 위암 연구의 기초 자료로 활용될 수 있을 것이라 사료된다.
감사의 말씀
본 연구는 한국연구재단 중견연구 개발비 지원(Grant No.
2011-0010706, 남석우)에 의해 수행 되었으며 가톨릭대학교의과 대학의 IRB로 부터 승인(CUMC 09U116)받아 연구를 수행하였 으며 이에 감사드립니다.
Fig. 3− Expression of NLK in gastric cancer cell lines. (A) Endogenous expression level of NLK between human gastric mucosa and cancer cell lines by quantitative real-time reverse transcriptase PCR. NLK mRNA levels were normalized against GAPDH and data are presented as the means±standard deviations of three experiments (unpaired student t test, *P<0.05 vs. GES-1). (B) Representative agarose gel image of PCR product for the expression of NLK. The GAPDH was used to control for RNA loading. (C) Endogenous NLK expression between human gastric mucosa and cancer cell lines by Western blot analysis. The α-tubulin was used as internal loading controls. (D) The graph bar shows the density of NLK protein expression relative to GAPDH.
참고문헌
1) Jemal, A., Murray, T., Ward, E., Samuels, A., Tiwari, R. C., Ghafoor, A., Feuer, E. J. and Thun, M. J. : Cancer statistics, 2005. CA Cancer J. Clin. 55, 10 (2005).
2) Jung, K. W., Park, S., Kong, H. J., Won, Y. J., Lee, J. Y., Seo, H. G. and Lee, J. S. : Cancer statistics in Korea: incidence, mortality, survival, and prevalence in 2009. Cancer Res. Treat.
44, 11 (2012).
3) Ishitani, T., Ninomiya-Tsuji, J., Nagai, S., Nishita, M., Meneghini, M., Barker, N., Waterman, M., Bowerman, B., Clevers, H., Shibuya, H. and Matsumoto, K. : The TAK1-NLK- MAPK-related pathway antagonizes signalling between beta- catenin and transcription factor TCF. Nature 399, 798 (1999).
4) Meneghini, M. D., Ishitani, T., Carter, J. C., Hisamoto, N., Ninomiya-Tsuji, J., Thorpe, C. J., Hamill, D. R., Matsumoto, K.
and Bowerman, B. : MAP kinase and Wnt pathways converge to downregulate an HMG-domain repressor in Caenorhabditis elegans. Nature 399, 793 (1999).
5) Choi, K. W. and Benzer, S. : Rotation of photoreceptor clusters in the developing Drosophila eye requires the nemo gene. Cell 78, 125 (1994).
6) Verheyen, E. M., Mirkovic, I., MacLean, S. J., Langmann, C., Andrews, B. C. and MacKinnon, C. : The tissue polarity gene nemo carries out multiple roles in patterning during Drosophila development. Mech. Dev. 101, 119 (2001).
7) Smit, L., Baas, A., Kuipers, J., Korswagen, H., van de Wetering, M. and Clevers, H. : Wnt activates the Tak1/Nemo- like kinase pathway. J. Biol. Chem. 279, 17232 (2004).
8) Ishitani, T., Ninomiya-Tsuji, J. and Matsumoto, K. : Regulation of lymphoid enhancer factor 1/T-cell factor by mitogen- activated protein kinase-related Nemo-like kinase-dependent phosphorylation in Wnt/beta-catenin signaling. Mol. Cell. Biol.
23, 1379 (2003).
9) Yasuda, J., Yokoo, H., Yamada, T., Kitabayashi, I., Sekiya, T. and Ichikawa, H. : Nemo-like kinase suppresses a wide range of transcription factors, including nuclear factor-kappaB. Cancer
Sci. 95, 52 (2004).
10) Kanei-Ishii, C., Ninomiya-Tsuji, J., Tanikawa, J., Nomura, T., Ishitani, T., Kishida, S., Kokura, K., Kurahashi, T., Ichikawa- Iwata, E., Kim, Y., Matsumoto, K. and Ishii, S. : Wnt-1 signal induces phosphorylation and degradation of c-Myb protein via TAK1, HIPK2, and NLK. Genes Dev. 18, 816 (2004).
11) Yasuda, J., Tsuchiya, A., Yamada, T., Sakamoto, M., Sekiya, T.
and Hirohashi, S. : Nemo-like kinase induces apoptosis in DLD-1 human colon cancer cells. Biochem. Biophys. Res.
Commun. 308, 227 (2003).
12) Tan, Z., Li, M., Wu, W., Zhang, L., Ding, Q., Wu, X., Mu, J. and Liu, Y. : NLK is a key regulator of proliferation and migration in gallbladder carcinoma cells. Mol. Cell. Biochem. 369, 27 (2012).
13) Jung, K. H., Kim, J. K., Noh, J. H., Eun, J. W., Bae, H. J., Xie, H. J., Ahn, Y. M., Park, W. S., Lee, J. Y. and Nam, S. W. : Targeted disruption of Nemo-like kinase inhibits tumor cell growth by simultaneous suppression of cyclin D1 and CDK2 in human hepatocellular carcinoma. J. Cell. Biochem. 110, 687 (2010).
14) Mirkovic, I., Charish, K., Gorski, S. M., McKnight, K. and Verheyen, E. M. : Drosophila nemo is an essential gene involved in the regulation of programmed cell death. Mech.
Dev. 119, 9 (2002).
15) Ohkawara, B., Shirakabe, K., Hyodo-Miura, J., Matsuo, R., Ueno, N., Matsumoto, K. and Shibuya, H. : Role of the TAK1- NLK-STAT3 pathway in TGF-beta-mediated mesoderm induction. Genes Dev. 18, 381 (2004).
16) Emami, K. H., Brown, L. G., Pitts, T. E., Sun, X., Vessella, R. L. and Corey, E. : Nemo-like kinase induces apoptosis and inhibits androgen receptor signaling in prostate cancer cells.
Prostate. 69, 1481 (2009).
17) Zhang, Y., Peng, C., Wu, G., Wang, Y., Liu, R., Yang, S., He, S., He, F., Yuan, Q., Huang, Y., Shen, A. and Cheng, C. : Expression of NLK and its potential effect in ovarian cancer chemotherapy. Int. J. Gynecol. Cancer 21, 1380 (2011).