체내지방의 과다축적으로 정의되는 비만은 미용상의 문 제만이 아니라 사회, 경제적으로도 큰 부담이 되는 질병으 로 비만에 동반되는 다른 대사성질환인 당뇨병, 동맥경화증 이 크게 증가하고 있는 실정이다. 이와 같은 질환들 (대사증 후군)의 공통적인 원인으로 잘 알려진 것이 인슐린저항성이 다. 최근 미토콘드리아의 기능이상이 제2형 당뇨병, 비만 및 인슐린저항성과 관련됨이 알려지면서 많은 주목을 받고 있 다.
골격근은 고인슐린 포도당 클램프 (euglycemic hyperinsulinemic clamp)와 같은 고인슐린혈증 상태에서 전 신 포도당 이용/저장의 70~80%를 차지하는 조직이다. 뿐만 아니라 인슐린저항성 상태에서 가장 뚜렷한 포도당 이용의 감소를 보여 많은 학자들이 골격근에서의 포도당 이용의 감 소가 인슐린저항성의 일차적 원인이 될 것으로 생각하고 있 다. 골격근에서의 포도당 이용의 감소는 포도당의 세포 내 로의 운반 과정 (glucose transport), 포도당의 인산화 과정, 당원으로의 합성 단계 (glycogen synthesis) 등 여러 단계의 장애에 기인할 수 있는데 이 중 당원 합성과정이 인슐린저 항성 상태에서 양적으로 가장 뚜렷하게 감소된다1). 한편 골 격근에서의 포도당 운반 과정, 당원 합성 과정 등이 insulin receptor substrate-1 (IRS-1) 인산화와 phosphatidyl inositol-3 kinase (PI3K)와 같은 세포 내 인슐린 신호 전달 체계의 조절을 받고 이들 신호전달 체계의 활성이 비만 및 제2형 당뇨병 환자의 골격근에서 감소되어 있음이 여러 연 구에서 밝혀져 있다2).
Glucose-fatty acid cycle
1963년 Randle 등은 근육 내 지방산 산화의 증가가 포도 당 산화를 감소시킴을 보고하고 혈중 유리 지방산 농도의
증가에 따른 근육 내 지방산 산화의 증가가 인슐린저항성의 원인이 됨을 제시하였다 (glucose fatty acid cycle)3). 이 가 설에 의하면 지방산 산화의 증가는 미토콘드리아의 acetyl -CoA/CoA 비를 증가시켜서 피루브산 탈수소효소 (pyruvate dehydrogenase)의 활성도를 억제함으로써 포도당 산화를 억제하고, 세포 내에 증가된 citrate는 phosphofructokinase 를 억제하여 glucose-6-phosphate의 축적을 일으킴으로써 결과적으로 포도당의 세포 내 유입을 억제한다는 것이다.
이 가설은 그 이후 고지방 식이나4) 지방질 정맥 투여를 통 한5) 여러 실험을 통해 증명되었다.
지방산 산화의 증가가 골격근 내에서 포도당 산화의 감 소를 일으킨다는 사실은 거의 모든 연구자들에 의해 확인되 었지만 실제 비만이나 당뇨병환자에서 보이는 인슐린저항 성의 발생에 포도당 산화보다 더 중요한 기여를 하는 당원 합성이 감소되는지에 대해서는 논란의 여지가 있었다. 대부 분의 초기 연구들은 혈액 내 지방산 농도를 높일 경우 골격 근 내 포도당 산화뿐만 아니라 당원 합성도 감소함을 보고 한 반면 Kim 등은 고지방 식이에 의한 유리 지방산의 증가 가 골격근에서의 해당작용을 억제시키나 glucose-6 -phosphate 농도 및 당원 합성은 오히려 증가시킨다고 하였 다4). 이와 같은 controversy를 해결하기 위해 본 교실에서는 쥐에 지방질을 5시간 동안 정맥 주사하면서 시간에 따른 포 도당 대사의 변화를 관찰하였다. 연구에 따르면, 지방질 투 여 직후부터 포도당 산화를 대변하는 해당작용이 뚜렷하게 억제되었으나 초기에는 세포 내 glucose-6-phosphate 농도 가 올라가면서 당원 합성이 보상적으로 증가하고 이에 따라 전신 및 골격근 내 포도당 이용은 변하지 않았다. 그러나 시 간이 경과함에 따라 당원 합성 역시 감소하기 시작하였고 전신 포도당 이용 역시 뚜렷하게 감소함을 보여 지방산 산화 가 장기적으로 증가할 경우 당원합성이 감소됨을 보였다6).
미토콘드리아 기능장애와 인슐린저항성
울산대학교 의과대학 서울아산병원 내분비내과
조은희․고은희․김민선․박중열․이기업 Mitochondrial Dysfunction and Insulin Resistance
Cho EH, Koh EH, Kim MS, Park JY, Lee KU
Division of Endocrinology and Metabolism, Department of Internal Medicine, Asan Medical Center, University of Ulsan College of Medicine
한편 최근의 연구들은 지방질 투여가 앞에서 기술한 acetyl CoA나 citrate를 통해 포도당 대사 이상을 초래하는 이외에 도 PI3K와 같은 인슐린 신호 전달체계의 변화를 유발하여 인슐린저항성을 유발함을 밝히고 있다7).
세포 내 중성지방 및 지방산 대사산물 축적
이상 혈액 내 유리지방산의 증가가 골격근 내 포도당 산 화와 당원 합성을 억제하여 인슐린저항성을 유발한다는 사 실은 믿어 의심할 여지가 없다. 그러나 실제 혈중 지방산 농 도와 인슐린 감수성에 대한 상관계수는 0.6 이하로 보고되 어 왔다. 즉 혈중 유리 지방산의 증가만으로는 인슐린저항 성의 발생을 충분히 설명하지 못한다는 것이다. 근래에 비 만한 환자나 동물모델에서 골격근 조직 내 중성지방 축적이 증가되어 있음을 알게 되었고 이로부터 혈액 내 지방산 뿐 만 아니라 골격근에 저장되어 있던 중성지방으로부터의 국 소적인 지방분해에 의해 공급되는 지방산 역시 인슐린저항 성 발생에 중요한 역할을 할 것이라는 개념이 대두되었다8). 세포 내 지방산 대사산물인 long-chain fatty acyl-CoA (LCAC)는 glycogen synthase와 같은 포도당 대사 효소의 활성도를 억제하고 protein kinase C나 IkappaB 같은 다양 한 세포 내 신호전달 물질의 변화를 초래함이 밝혀졌고9), 장기간의 운동 훈련이나 단시간의 운동을 통해 세포 내 LCAC 농도를 감소시킬 경우 인슐린감수성이 뚜렷하게 감 소함이 밝혀져10) 세포 내 LCAC의 축적이 인슐린저항성의 주 원인기전이라는 주장이 강력하게 제기되고 있다. 뿐만 아니라 비만이나 당뇨병 동물모델에서는 혈관 조직 내에도
중성지방 및 LCAC의 축적이 증가되어 있는데, 증가된 LCAC는 세포 내 산화 스트레스를 증가시킴으로써 동맥경 화증 발생에 관여할 것이라는 가설이 제시된 바 있다11).
인슐린저항성 동물 모델에서 골격근 지방 분해의 감소
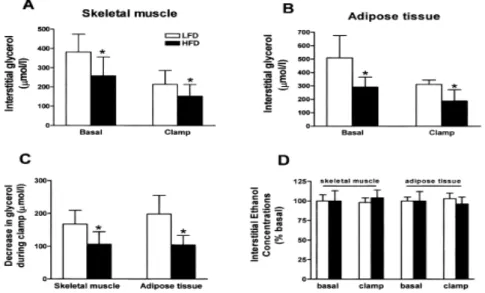
이상 대사증후군에서 골격근이나 혈관 조직에 지방산 대 사산물이 축적되고 이로 인해 인슐린저항성이나 동맥경화 증이 발생할 것이라는 학설이 최근 많은 학자들의 지지를 받고 있으나 이와 같은 현상이 나타나는 기전에 대한 연구 는 매우 부족한 상태이다. 골격근 세포 내에 축적된 중성지 방이 세포 내에서 분해되어 지방산 산화를 촉진시키고 이에 따라 인슐린저항성을 유발한다면 골격근으로부터의 지방 분해가 증가되어 있을 것이라고 가정할 수 있을 것이다. 그 러나 본 교실에서 시행한 연구결과12), 고지방식이를 투여한 백서에서 골격근 및 지방조직 간질의 글리세롤 농도가 기저 상태 및 고인슐린 포도당 클램프 모두에서 저지방식이군보 다 유의하게 낮았다 (Fig. 1). 이상의 결과는 기존의 개념과 는 상반되는 것으로 인슐린저항성 상태에서 관찰되는 골격 근 조직 내의 지방축적 증가는 세포 내에서의 지방산 산화 의 감소에 의해 나타남을 시사하고 있다.
미토콘드리아와 인슐린저항성
미토콘드리아는 세포 내에서 대부분의 ATP를 생산하는 소기관으로 이 외에도 산소 라디칼 형성, 세포 자멸사 및 세 포내 칼슘대사 등에서 중요한 역할을 한다. ATP의 주 원료
Fig. 1. Interstitial glycerol concentration in (A) skeletal muscle and (B) adipose tissue under basal and hyperinsulinemic euglycemic clamp conditions, and (C) net change in the interstitial glycerol concentrations during clamp (D) interstitial ethanol concentrations in skletal muscle and adipose tissue during microdialysis12).
는 포도당과 지방산이나 특히 기저상태의 골격근 세포에서 는 주로 지방산이 이용되기 때문에 미토콘드리아 기능 이상 이 있을 경우에는 세포 내 지방산 산화가 감소하게 되고 이 에 따라 LCAC의 축적이 일어나게 된다.
당뇨병에서 세포 내 에너지 결핍이 나타난다는 것은 잘 알려진 사실로 미토콘드리아 기능과 인슐린 작용이 연관되 어 있으리라는 것을 쉽게 예상할 수 있으나, 실제 당뇨병과 미토콘드리아 기능과의 관계가 알려진 것은 그리 오래되지 않았다. 1990년대 초 미토콘드리아 DNA에 돌연변이가 있 을 경우 모계로 유전되는 당뇨병이 나타난다는 것이 처음 보고된 이후13) 여러 종족에서 이 사실이 확인되었다. 미토 콘드리아 기능과 당뇨병의 관계에 대해서는 1998년 Lee 등 이 말초혈액 내 미토콘드리아의 DNA양이 감소가 제2형 당 뇨병 발생에 선행함을 처음 보고하였고14), 이후 Petersen 등 은 제2형 당뇨병환자의 자녀에서 보이는 근육에서의 인슐린 저항성이 근육 내 지방산대사의 이상에 의한 중성지방의 지 방 내 침착과 연관이 있으며, 주로 미토콘드리아의 산화적 인산화의 유전적 결함이 관여한다고 발표하였고15), 노인환 자에서의 인슐린저항성은 주로 미토콘드리아의 생성에 대 한 후천적인 결함으로 근육 내 미토콘드리아의 감소와 연관 된다고 보고하여16), 미토콘드리아 기능 저하와 인슐린저항 성의 연관성을 제시하였다.
PPARα 및 PPARγ 활성제에 의한 당뇨병 발생 예방
미토콘드리아에서의 지방산 산화를 조절하는 인자들로는 PPARα, PPARδ, PPARγ coactivator-1 (PGC-1) 및 AMPK (AMP-activated protein kinase) 등이 있다. 이 중 가장 잘 알려져 있는 PPARα는 핵수용체의 하나로서 활성시 지방산 수송 단백질 및 long-chain acyl-CoA synthase 발현을 유도 하여 미토콘드리아 내로의 지방산 흡수를 증가시키고,
acyl-CoA oxidase 및 carnitine palmitoyl transferase-1 (CPT-1)의 발현을 증가시켜 지방산 산화를 증가시킨다17). PPARδ는 최근 큰 관심을 끌고 있는 핵수용체로서 활성화 되면 지방산 산화가 증가되고 adaptive thermogenesis에 관 련된 uncoupling protein의 발현이 증가한다18). PGC-1은 미 토콘드리아의 합성과 호흡에 관여하는 다양한 전사조절인 자들 (NRF-1,2, mtTFA)의 전사활성을 촉진시키는 coactivator이며 미토콘드리아의 biogenesis, respiration 및 thermogenesis를 증가시키는 것으로 보고되었다19). 또한 갈 색지방조직에서의 열 생산, 근육에서의 포도당 이용을 증가 시키는데 간에서는 PEPCK 같은 포도당 신생 조절 효소를 활성화시키는 것으로 보고되고 있다20). 이외에도 핵호르몬 수용체인 liver X receptor는 지방산 합성 효소인 fatty acid synthase와 지방세포 분화에 관여하는 SREBP-1의 활성을 증가시켜 지방산 산화의 negative regulator로 작용할 가능 성이 있다21).
본 교실에서는 PPARα activator인 fenofibrate 및 PPARγ activator인 rosiglitazone이 비만 및 당뇨병 모델 쥐인 OLETF rats에서 당뇨병 발생을 예방함을 발견하였다22). 알 려진 바와 같이 rosiglitazone 투여군에서는 체중 증가 소견 을 보였는데 이와는 반대로 fenofibrate 투여군에서는 체중 과 내장 지방의 감소를 보였다. 지방조직에 대한 반대 효과 와는 달리 두 투여군 모두 골격근에서 지방산 산화를 증가 시켰고 중성지방 축적을 감소시켰다. 즉 PPARα뿐만 아니라 PPARγ agonist도 골격근 세포에서 지방산 산화를 증가시킴 으로서 인슐린 감수성의 호전을 가져옴이 시사되었다.
AMPK와 지방산 산화
AMP-activated protein kinase (AMPK)는 세포 내 에너 지 상태를 감지하는 효소로서 세포 내 에너지가 부족한 상 황, 즉 ATP에 비해 AMP가 증가하는 상황에서 활성화되어
↑ A M P K A l p h a - lip o ic a c id
R o s ig lit a z o n e L e p t in A d ip o n e c t in
↑ M it o c h o n d r ia l b io g e n e s is ↓ A C C ↑ P G C - 1 / P P A R α
↑ F a t t y a c id o x id a t io n
↓ In t r a c e llu la r L ip id m e t a b o lite s
↓ In s u lin re s is ta n c e
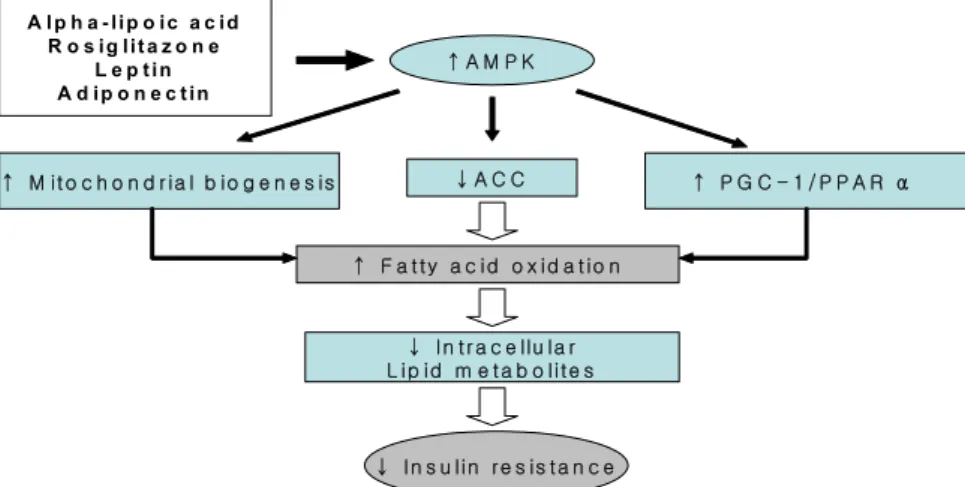
Fig. 2. AMPK in fatty acid oxidation and insulin resistance.
정상 에너지 균형을 회복시키기 위해 다양한 대사경로를 조 절한다23). AMPK는 근육에서 포도당 및 지방산 대사의 중 요한 조절인자로 작용하며, 특히 운동을 하는 근육에서 활 성화된다24). 골격근에서 AMPK가 활성화되면 포도당 이용 이 증가되는 이외에도 acetyl CoA carboxylase (ACC)의 인 산화를 통해 ACC의 작용을 억제함으로써 malonyl CoA 농 도가 감소된다. Malonyl CoA는 미토콘드리아 내로의 LCAC의 이송을 조절하는 CPT-1을 억제하는 역할을 하는 데 AMPK가 활성화되어 malonyl CoA가 감소되면 결과적 으로 CPT-1에 대한 억제 효과가 감소하게 되어 지방산 산 화가 증가된다25).
본 교실에서는 최근 연구에서 C2C12 골격근 세포에서 AMPK 활성물질인 5-aminoimidazole-4-carboxamide ribonucleoside (AICAR)가 PPAR-α 목표 유전자와 PPARγ coactivator-1 (PGC-1)의 발현을 증가시켜 지방산 산화를 증가시키고, PPAR-α와 PGC-1에 대한 siRNAs처리 시 AICAR에 의한 지방산산화의 증가를 억제함을 보여26), AMPK가 기존의 알려진 ACC의 작용을 억제하는 외에도 PPAR-α와 PGC-1를 통해서도 지방산 산화를 증가시킴을 증명하였다 (Fig. 2).
Alpha-lipoic acid (ALA)와 대사증후군
Alpha-lipoic acid (ALA)는 두 개의 황산 분자를 포함하 는 지방산으로 미토콘드리아의 호흡효소인 피루브산 탈수 소효소 (pyruvate dehydrogenase)의 필수 조효소로 노화한 쥐에서 미토콘드리아의 기능을 호전시키는 것으로 알려져 있다27). ALA는 매우 강력한 항산화제로 주로 당뇨병성 신 경병증의 치료제로 쓰이는 약제이다. 본 교실에서는 ALA의 장기 투여가 비만형 당뇨병 동물 모델인 OLETF (Otsuka Long-Evans Tokushima Fatty) 쥐에서 당뇨병의 발생을 예 방함을 발견하였다. ALA의 투여는 OLETF 쥐에서 나타나 는 췌장소도의 파괴를 억제하였으며, 골격근 및 췌장소도에 서의 지방축적을 감소시켰다28). ALA를 투여한 쥐에서 AMPK 활성도가 증가하였고 지방산의 산화의 증가와 중성 지방축적의 감소가 관찰되었다29).
한편 OLETF 쥐에서는 대조군에 비해 혈관내피의존성 혈관확장에 장애가 있고, 내피세포 사멸이 증가된 소견을 보였으며, 내피세포 내 중성지방 및 lipid peroxide가 증가 되어 있고, 산화질소 (Nitric oxide)생산은 감소되어 있었는 데, OLETF 쥐에 ALA를 투여 시 혈관내피 의존성 혈관확 장 장애의 호전을 보였고, 내피세포 사멸이 억제되었으며, 중성지방의 축적과 lipid peroxide는 감소시켰으며 산화질소 는 증가되었다30). 이러한 결과를 바탕으로 ALA가 AMPK 의 활성을 증가시키고, 지방산 산화를 증가시킴으로써
LCAC가 감소시킴을 알 수 있었고 이 기전에 의해 당뇨병 이 감소하고, 혈관기능이 개선되어 동맥경화증이 감소함을 추론할 수 있었다.
Uncoupling protein-2 (UCP-2)에 의한 혈관기능 호전
미토콘드리아는 세포 막에 있는 NADPH oxidase와 함께 세포 내에서 활성산소족 (ROS)을 생산하는 제일 중요한 부 위 중의 하나이다. TCA cycle에 의해 만들어진 NADH는 NAD+로 환원되면서 ATP를 생산하게 된다. 이 과정에서 미토콘드리아 내막의 electron transfer chain을 통해 전자를 이동시키는 한편 내막 및 외막 사이의 intermembranous space에 수소이온 (H+)을 축적시킴으로써 ATP 생산에 필요 한 에너지를 축적시킨다. 이와 같이 에너지 (mitochondrial membrane potential; MMP)를 축적하는 것은 ATP 생산을 위한 필수적인 과정이지만 어떠한 이유에서든 장기적으로 증가된 MMP가 ATP 생산과 같은 과정을 통하여 해결이 되 지 않을 경우에는 electron transfer chain의 활동이 저해되 게 되고 정상적으로 산소와 만나 물 분자가 되어야 하는 전 자가 ROS를 형성하게 된다.
Uncoupling protein (UCP)는 갈색지방조직에서 처음 발 견된 단백질로서 축적된 MMP를 ATP 생산에 이용하는 대 신 열로 발산시키는 기능을 가진다. 갈색지방에 있는 UCP 이외에도 이와 유사한 물질이 다른 조직에 존재함이 알려지 고 있는데, 그 중 uncoupling protein-2 (UCP-2)는 혈관세 포를 포함한 다양한 여러 조직에서 발현된다. UCP-2가 혈 관기능에 미치는 영향을 확인하기 위하여 혈관내피세포에 과발현시켰을 때 혈관확장에 관여하는 eNOS (endothelial nitric oxide synthase)의 mRNA 발현이 증가하였고, 혈관수 축에 관여하는 endothelin-1의 발현은 감소하였다. 또한 UCP-2는 혈관내피세포에서 lysophosphatidylcholine이나 linoleic acid에 이해 유발되는 ROS 증가, NF-kB 활성 및 세포사멸을 억제하였으며 이는 UCP-2가 혈관내피세포에서 ROS의 생성을 조절하는 중요한 역할을 함을 시사한다31). 또한 혈관평활근세포에서 고농도의 포도당과 안지오텐신 II 를 처리하였을때 세포 내 산화 스트레스 및 전사인자 AP-1, PAI-1 발현 및 NADPH oxidase가 활성화되고 동맥경화증 의 발생지표인 혈관평활근세포의 증식 및 이주 또한 증가하 였다. 하지만, UCP-2를 혈관평활근세포에 과발현시켰을 때 이러한 변화가 현저하게 억제되는 것을 관찰할 수 있었다32).
이와 같은 결과는 UCP-2가 혈관내피세포에서 미토콘드 리아의 ROS 생성을 조절함으로 동맥경화증의 발생을 줄일 수 있음을 보여주는 것으로, 미토콘드리아의 기능이상이 동 맥경화증의 발생에 연관됨을 시사하고 있다.
미토콘드리아와 아디포넥틴 생산
최근 지방조직이 능동적으로 호르몬 (adipokine, adipocytokine)을 분비함으로써 체내 에너지 항상성 유지에 중요한 역할을 한다는 개념이 보편화되고 있다. 아디포넥틴 (Adiponectin)은 지방세포에서 다른 adipokine에 비해 매우 과량으로 생산되는 물질인데 골격근세포나 혈관세포에서 AMPK를 활성화시켜 당뇨병이나 동맥경화증 발생을 예방 할 수 있음이 밝혀져 큰 관심을 끌고 있다34,35). 비만 환자에 서 혈액 내 다른 adipokine 농도는 증가하나 아디포넥틴 농 도는 역설적으로 감소되어 있는데, 아직까지 이와 같은 현 상이 나타나는 기전은 밝혀져 있지 않았다.
본 교실에서는 최근 지방세포에서의 아디포넥틴 생산에 미토콘드리아의 정상적인 기능이 필수적임을 발견하였고 비만 쥐의 지방세포에서 미토콘드리아의 양 및 기능이 저하 됨을 밝힘으로써 비만 환자에서 혈액 내 아디포넥틴 농도가 감소되는 paradox를 설명할 단서를 찾았다 (미발표 자료).
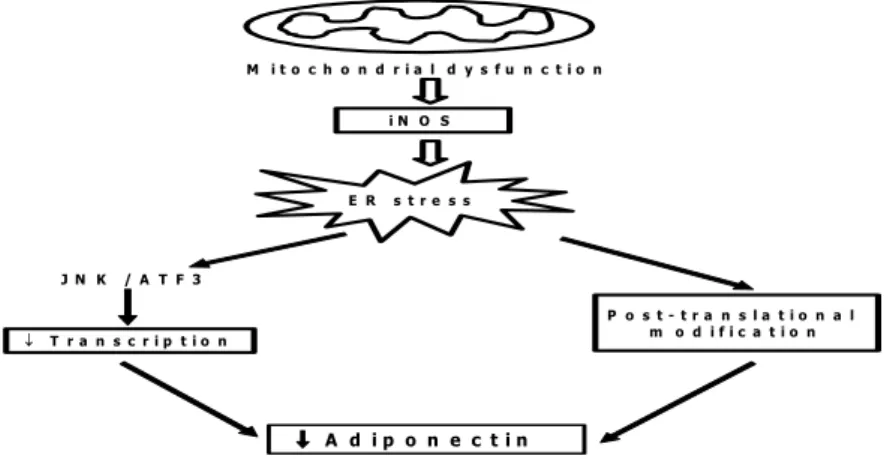
배양된 지방세포에 여러 가지 미토콘드리아 기능 저해 물질 을 처리하였을 때 iNOS (inducible nitric oxide synthase)활 성화, endoplasmic reticulum stress, JNK 활성화 및 ATF3 발현 증가를 통해 아디포넥틴의 전사 (transcription)가 감소 하였고, 고분자량 아디포넥틴 (high molecular weight adiponectin)생산도 감소하였다. 반면 rosiglitazone이나 nuclear respiratory factor-1 (NRF-1)에 의해 미토콘드리아 생성을 증가시켰을 때, 아디포넥틴 생산이 증가함을 보임으 로써 이제까지 알려지지 않았던 지방세포 미토콘드리아 기 능을 처음으로 밝혔다 (Fig. 3).
결 론
혈액 내 유리지방산의 증가가 골격근 내 포도당 산화와 당원 합성을 억제하여 인슐린저항성을 유발하나, 혈중 유리 지방산의 증가만으로는 인슐린저항성의 발생을 충분히 설 명하지 못한다. 근래 비만한 환자나 동물모델에서 골격근 조직 내 중성지방 축적이 증가되어 있음이 밝혀졌고 이와 같은 지방축적의 증가는 세포 내에서의 지방산 산화의 감소 에 의한다고 여겨지고 있다. 세포 내 지방산 산화의 감소로 인해 대사산물인 long-chain fatty acyl-CoA (LCAC)가 축 적되고, 이러한 LCAC가 glycogen synthase와 같은 포도당 대사 효소의 활성도를 억제하고 protein kinase C나 IkappaB 같은 다양한 세포 내 신호전달 물질을 변화시켜 인슐린저항성을 일으킨다고 생각되고 있다.
미토콘드리아에서 지방산 산화의 대부분이 일어나는데, PPARα, PPARδ, PGC-1 및 AMPK 등은 지방산 산화를 증 가시켜 인슐린 감수성을 증가시킨다. 한편 최근의 연구에 의해 미토코드리아 기능 이상과 ER stress의 연관성이 밝혀 지고 있어, 향후 미토콘드리아의 기능이상과 ER stress 그리 고 인슐린저항성에 대한 좀 더 많은 관심이 요구된다.
참 고 문 헌
1. Wright KS, Beck-Nielsen H, Kolterman OG, Mandarino LJ: Decreased activation of skeletal muscle glycogen synthase by mixed-meal ingestion in NIDDM. Diabetes 37:436-46, 1998
2. Kim YB, Nikoulina SE, Ciaraldi TP, Henry RR, Kahn BB: Normal insulin-dependent activation of
i N O S
↓ T r a n s c r i p t i o n
P o s t - t r a n s l a t i o n a l m o d i f i c a t i o n M i t o c h o n d r i a l d y s f u n c t i o n
E R s t r e s s
J N K / A T F 3
A d i p o n e c t i n
Fig. 3. Schematic model of the mechanism by which adiponectin synthesis is reduced in obesity.
Adipocyte hypertrophy in obesity is associated with mitochondrial dysfunction, and consequent activation of iNOS and ER stress affect transcription and post-translational modification to result in decreased production of adiponectin.
Akt/protein kinase B, with diminished activation of phosphoinositide 3-kinase, in muscle in type 2 diabetes. J Clin Invest 104:733-41, 1999
3. Randle PJ, Garland PB, Hales CN, Newsholm EA:
The glucose-fatty acid cycle: its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1:785-9, 1963
4. Kim JK, Wi JK, Youn JH: Metabolic impairment precedes insulin resistance in skeletal muscle during high-fat feeding in rats. Diabetes 45:651-8, 1996 5. Lee KU, Lee HK, Koh CS, Min HK: Artificial
induction of intravascular lipolysis by lipid-heparin infusion leads to insulin resistance in man.
Diabetologia 31:285-90, 1988
6. Park JY, Kim CH, Hong SK, Suh KI, Lee KU:
Effects of FFA on insulin-stimulated glucose fluxes and muscle glycogen synthase activity in rats. Am J Physiol 275:E338-44, 1998
7. Dresner A, Laurent D, Shulman GI: Effects of free fatty acids on glucose transport and IRS-1-associated phosphatidylinositol 3-kinase activity. J Clin Invest 103:253-9, 1999
8. Kelly DE, Goodpaster BH, Storlien L: Muscle triglyceride and insulin resistance. Annu Rev Nutr 22:325-46, 2002
9. Itani SI, Ruderman NB, Schmieder F, Boden G:
Lipid-induced insulin resistance in human muscle is associated with changes in diacylglycerol, protein kinase C, and IkappaB-alpha. Diabetes 51:2005-11, 2002.
10. Gan SK, Kriketos AD, Ellis BA, Thompson CH, Kraegen EW, Chisholm DJ: Changes in aerobic capacity and visceral fat but not myocyte lipid levels predict increased insulin action after exercise in overweight and obese men. Diabetes Care 26:1706-113, 2003
11. Bakker SJ, IJzerman RG, Teerlink T, Westerhoff HV, Gans RO, Heine RJ: Cytosolic triglycerides and oxidative stress in central obesity: the missing link between excessive atherosclerosis, endothelial dysfunction, and beta-cell failure? Atherosclerosis 148 :17-21, 2000
12. Kim CH, Kim MS, Youn JY, Park HS, Song HS, Song KH, Park JY, Lee KU: Lipolysis in skeletal muscle is decreased in high-fat-fed rats. Metabolism 52:1586-92, 2003
13. van den Ouweland JM, Lemkes HH, Ruitenbeek W, Sandkuijl LA, de Vijlder MF, Struyvenberg PA, van de Kamp JJ, Maassen JA: Mutation in mitochondrial tRNA(Leu)(UUR) gene in a large pedigree with maternally transmitted type II diabetes mellitus and deafness. Nat Genet 1:368-71, 1992
14. Lee HK, Song JH, Shin CS, Park KS, Lee KU, Koh CS: Decreased mitochondrial DNA content in peripheral blood precedes the development of non-insulin-dependent diabetes mellitus. Diabetes Res Clin Prac 42:161-7, 1998
15. Petersen KF, Dufour S, Befroy D, Garcia R, Shulman GI: Impaired Mitochondrial Activity in the Insulin-Resistant Offspring of Patients with Type 2 Diabetes. N Engl J Med 350:664-71, 2004
16. Petersen KF, Befroy D, Dufour S, Dziura J, Ariyan C, Rothman DL, DiPietro L, Cline GW, Shulman GI:
Mitochondrial dysfunction in the elderly: possible role in insulin resistance. Science 300:1140-2, 2003 17. Ferre P: The biology of peroxisome proliferator
-activated receptors: relationship with lipid metabolism and insulin sensitivity. Diabetes 53:S43-50, 2004
18. Wang YX, Lee CH, Tiep S, Yu RT, Ham J, Kang H, Evans RM: Peroxisome-proliferator-activated receptor delta activates fat metabolism to prevent obesity. Cell 113:159-70, 2003
19. Puigserver P, Spiegelman BM: Peroxisome proliferator-activated receptor-γ coactivator 1α (PGC-1α): Transcriptional coactivator and metabolic regulator. Endocr Rev 24:78-90, 2003
20. Rhee J, Inoue Y, Yoon JC, Puigserver P, Fan M, Gonzalez FJ, Spiegelman BM: Regulation of hepatic fasting response by PPARgamma coactivator-1alpha (PGC-1): requirement for hepatocyte nuclear factor 4alpha in gluconeogenesis. Proc Natl Acad Sci 100:4012-7, 2003
21. Steffensen KR, Gustafsson JA: Putative metabolic effects of the liver X receptor (LXR). Diabetes 53:S36-42, 2004
22. Koh EH, Kim MS, Park JY, Kim HS, Youn JY, Park HS, Youn JH, Lee KU: Peroxisome proliferator -activated receptor (PPAR)-alpha activation prevents diabetes in OLETF rats: comparison with PPAR-gamma activation. Diabetes 52:2331-7, 2003 23. Hardie DG, Carling D: The AMP-activated protein
kinase: Fuel gauge of the mammalian cell? Eur J Biochem 246:259-73, 1997
24. Kurth-Kracz EJ, Hirshman MF, Goodyear LJ, Winder WW: 5'AMP-activated protein kinase activation causes GLUT4 translocation in skeletal muscle.
Diabetes 48:1-5, 1999
25. Saha A, Ruderman NB: Malonyl-CoA and AMP-activated protein kinase: An expanding partnership. Mol Cell Biochem 253:65-70, 2003 26. Lee WJ, Kim M, Park HS, Kim HS, Jeon MJ, Oh
KS, Koh EH, Won JC, Kim MS, Oh GT, Yoon M, Lee KU, Park JY: AMPK activation increases fatty acid oxidation in skeletal muscle by activating PPARalpha and PGC-1. Biochem Biophys Res Commun 340:291-5, 2006
27. Hagen TM, Ingersoll RT, Lykkesfeldt J, Liu J, Wehr CM, Vinarsky V, Bartholomew JC, Ames AB:
(R)-Alpha-lipoic acid-supplemented old rats have improved mitochondrial function, decreased oxidative damage, and increased metabolic rate, FASEB J 13:411-8, 1999
28. Song KH, Lee WJ, Koh JM, Kim HS, Youn JY, Park HS, Koh EH, Kim MS, Youn JH, Lee KU, Park JY:
alpha-Lipoic acid prevents diabetes mellitus in diabetes-prone obese rats. Biochem Biophys Res Commun 326:197-20, 2005
29. Lee WJ, Song KH, Koh EH, Won JC, Kim HS, Park HS, Kim MS, Kim SW, Lee KU, Park JY:
alpha-Lipoic acid increases insulin sensitivity by activating AMPK in skeletal muscle. Biochem Biophys Res Commun 332:885-91, 2005
30. Lee WJ, Lee IK, Kim HS, Kim YM, Koh EH, Won JC, Han SM, Kim MS, Jo I, Oh GT, Park IS, Youn JH, Park SW, Lee KU, Park JY: Alpha-lipoic acid
prevents endothelial dysfunction in obese rats via activation of AMP-activated protein kinase.
Arterioscler Thromb Vasc Biol 25:2488-94, 2005 31. Lee KU, Lee IK, Han J, Song DK, Kim YM, Song
HS, Kim HS, Lee WJ, Koh EH, Song KH, Han SM, Kim MS, Park IS, Park JY: Effects of recombinant adenovirus-mediated uncoupling protein 2 overexpression on endothelial function and apoptosis.
Circ Res 96:1200-7, 2005
32. Park JY, Park KG, Kim HJ, Kang HG, Ahn JD, Kim HS, Kim YM, Son SM, Kim IJ, Kim YK, Kim CD, Lee KU, Lee IK: The effects of the overexpression of recombinant uncoupling protein 2 on proliferation, migration and plasminogen activator inhibitor 1 expression in human vascular smooth muscle cells.
Diabetologia 48:1022-8, 2005
33. Yamauchi T, Kamon J, Minokoshi Y, Ito Y, Waki H, Uchida S, Yamashita S, Noda M, Kita S, Ueki K, Eto K, Akanuma Y, Froguel P, Foufelle F, Ferre P, Carling D, Kimura S, Nagai R, Kahn BB, Kadowaki T: Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat Med 8:1288-95, 2002
34. Lindsay RS, Funahashi T, Hansen RL, Matsuzawa Y, Tanaka S, Tatarnni PA, Knowler WC, Krakoff J:
Adiponectin and development of type 2 diabetes in Pima Indian population. Lancet 360:57-8, 2002 35. Kumada M, Kihara S, Sumitsuji S, Kawamoto T,
Matsumoto S, Ouchi N, Arita Y, Okamoto Y, Shimomura I, Hiraoka H, Nakamura T, Funahashi T, Matsuzawa Y: Osaka CAD Study Group. Coronary artery disease. Association of hypoadiponectinemia with coronary artery disease in men. Arterioscler Thromb Vas Biol 23:85-9, 2003