Identification of differentially expressed genes using an annealing control primer system in periodontitis
6
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
수치

관련 문서
Comparing the gene expression profiles be- tween fungal infection and control silkworm, we detected 10 genes that were differentially expressed in fungal induction
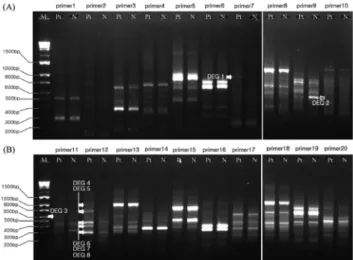
In this study, we applied annealing control primer (ACP)-based reverse transcriptase PCR (polymerase chain reaction) technique to identify differentially expressed
By using different gene fishing primer we identified several Cu- induced genes including UDP-glucuronic acid decarboxylase (DEG 4), transmembrane protein (DEG 10), small heat
